
烟草种质资源抗马铃薯Y 病毒病基因型鉴定
2021-11-22 09:17林世锋王仁刚任学良李尊强元野龙明锦张吉顺王自力
中国烟草学报 2021年5期
林世锋,王仁刚*,任学良,李尊强,元野,龙明锦,张吉顺,王自力
1 贵州省烟草科学研究院,烟草行业分子遗传重点实验室,贵阳市观山湖区龙滩坝路29号 550081;
2 中国烟草总公司黑龙江省烟草公司牡丹江烟草科学研究所,哈尔滨市道里区哈药路17号 150076
由马铃薯Y 病毒(Potato virus Y,PVY)引起的烟草PVY 病毒病是影响烟草生产的主要病害之一。实践证明,选育和种植抗病品种是控制该病害最经济有效的措施[1]。抗病种质资源的发掘、研究和利用是烟草抗PVY 育种的重要基础。已知烟草PVY 的抗源分为两大类,一类是以VAM(TI1406)及其衍生种质TN86 为代表的隐性基因位点(VAM和va)控制的抗性[2-3],另一类是以非洲烟草(Nicotiana africana)为代表的对PVY 免疫的抗性[4]。目前,受va位点控制的抗源被广泛应用于烟草抗病育种。为了提高va位点的育种利用效率,国内外研究者相继开发了与va位点相关的分子标记,如Randomly Amplified Polymorphic DNA(RAPD) 和Sequence Characterized Amplified Region(SCAR)标记[5-7]。这些标记与va位点间的遗传距离较远,导致利用标记数据判断抗性表型误差较大,进而限制了这类分子标记在育种中的实际应用。
在抗性获得的研究中,Noguchis 等[5]认为va基因型烟草植株的抗性是由于对PVY 感病的基因片段的缺失造成的。2014 年,Julio 等[8]通过更加深入的研究,进一步明确了va基因型烟草植株对PVY的抗性是由于真核翻译起始因子4E1(eukaryotic translation initiation factor 4E1,elF4E1)基因的缺失造成的,即eIF4E1基因为烟草隐性抗PVY 基因(感病基因)。2015 年,刘勇等[9]根据烟草eIF4E1基因及其家族基因的序列信息,开发出一个与eIF4E1野生型等位基因紧密连锁的显性标记,随后利用该标记进行分子标记辅助选择育成抗PVY 新品种云烟301[10],但由于该标记不能作为eIF4E1等位基因缺失的共显性标记,降低了其在实际育种中的应用价值和意义。2018 年,Dluge 等[11]利用RNA 测序的方法比较了VAM基因型、va基因型和野生型三种烟草中的基因表达差异,发现在具有PVY 抗性的VAM和va基因型烟草中,真核翻译起始因子eIF4E1基因所在的染色体区段发生大范围的缺失,由于建立eIF4E1等位基因缺失的共显性标记需要了解该基因缺失连接片段的序列特点,而eIF4E1基因及其侧翼序列的大范围缺失,造成研究者未能有效地克隆到该基因的缺失连接片段、获得序列信息,从而未能开发出等位基因缺失的共显性标记。
作物种质资源中蕴含着大量优异等位基因,如何鉴定并将这些变异应用于作物遗传改良是种质资源研究的中心任务之一[12]。先前研究结果表明, 在烟草种质中存在一定数量的PVY 抗性资源,但有关种质资源基因型,特别是有关烟草eIF4E1优异等位基因及其特异分子标记的研究报道甚少[13-15]。本研究目的是鉴定收集到的900 多份来自国内外的烟草种质资源的PVY 抗性表型,并通过抗性基因等位性测验和RTPCR 扩增测序相结合的方法,分析筛选出的抗源材料所携带的eIF4E1基因的基因型,以期为后续烟草PVY 抗性优异等位基因的发掘和育种利用提供基础材料和信息支撑。
1 材料与方法
1.1 试验材料
供试种质由贵州省烟草科学研究院提供,其中包括PVY 抗病对照烟草品种TN90 和感病对照烟草品种K326。以TN90 和K326 作母本与研究中筛选到的PVY 抗病资源作父本进行杂交获得F1 代杂交种材料,用于抗病资源中PVY 抗性基因的等位性检测。
1.2 PVY 人工接种
以普遍发生的脉坏死株系(PVYN)为接种毒源,由贵州省烟草科学研究院品种选育三室分离和保存。抗PVY 测试采用常规汁液摩擦接种法[16]。
1.3 烟草基因组DNA 的提取
采用AxyPrep 基因组DNA 小量制备试剂盒(Axygen)提取烟草基因组DNA,并通过紫外分光光度法(Nanodrop)和琼脂糖凝胶电泳法初步检测基因组DNA 的提取质量。
1.4 烟草RNA 提取和cDNA 合成
采用Invitrogen 公司的Trizol 试剂盒提取总RNA。cDNA 第一链的合成按照TaKaRa 公司的PrimeScriptTMⅡ 1st strand cDNA Synthesis Kit 说明书进行。
1.5 烟草eIF4E1 基因的特异分子标记检测
根据文献报道合成eIF4E1野生型等位基因的5’端显性分子标记引物[9]、开阳小黑烟和福泉柳叶烟eIF4E1突变型等位基因的共显性分子标记引物[17-18]及内参基因NR(硝酸还原酶基因)的特异性引物[19],同时设计合成eIF4E1野生型等位基因的3’端显性分子标记引物(表1),对筛选到的PVY 抗病资源中的eIF4E1基因进行PCR 扩增试验。
1.6 烟草eIF4E1 基因的全长cDNA 扩增
根据文献[20]合成eIF4E1基因全长序列扩增引物(表1),以各自烟草cDNA 为模板扩增eIF4E1基因全长cDNA 序列。PCR 扩增体系:TaKaRa LA Taq 酶0.25 μL,10×LA Taq Bufer Ⅱ2.5 μL,dNTP Mix 2 μL,正向引物(10 μmol/L)1 μL,反向引物(10 μmol/L)1 μL,模板1 μL,加入ddH2O 补平至25 μL。PCR 扩增程序:94℃预变性3 min,94℃变性30 s,94℃变性30 s,52℃退火30 s,72℃延伸1 min(30个循环),最后72℃延伸10 min。PCR 产物电泳检测,送上海生工公司进行基因测序。

表1 本研究所用引物序列Tab. 1 Primers used in this study

续表1
2 结果
2.1 抗性表型鉴定
从900 多份种质资源中,通过苗期盆栽人工接种PVY 抗性鉴定,获得抗PVY 资源开阳小黑烟、半坤村晒烟等19 份(表2)。抗PVY 资源大部分属于我国地方性晒烟及黄花烟品种。
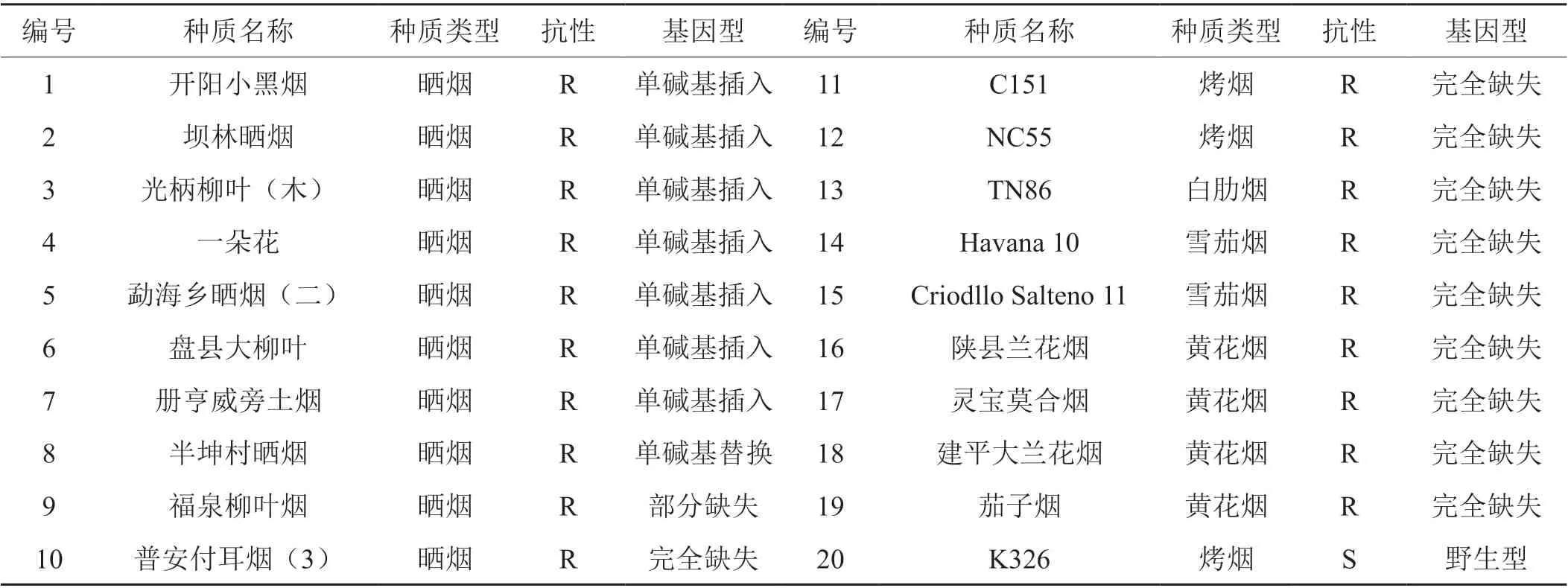
表2 抗PVY 的烟草种质资源信息Tab. 2 The information of the tobacco germplasm resource with resistance to PVY
2.2 与TN90 抗病位点的等位性检测
已知白肋烟品种TN90 对PVY 的抗性是由隐性基因位点(va)控制的,具体而言是由烟草PVY 感病基因eIF4E1缺失造成的[11,21]。为检测抗PVY 资源与TN90 对PVY 抗性间的等位性关系,利用抗PVY资源与TN90 及感病品种K326 配制抗抗及抗感杂交组合,各选取60 个F1代单株,通过人工摩擦接种法进行鉴定。结果显示,19 份抗PVY 资源与TN90 配制抗抗组合的F1代单株均表现为抗病,而19 份抗PVY 资源与K326 配制抗感组合的F1代单株均表现为感病。上述结果表明,筛选到的19 份抗PVY 资源与TN90 的PVY 抗病基因间具有等位性,即推断19份抗PVY 资源的PVY 抗性均是由感病基因eIF4E1缺失造成的。
2.3 eIF4E1 基因的特异分子标记检测
近年来研究发现烟草抗PVY 材料的eIF4E1基因可发生数种不同形式的突变,包括最为常见的基因全序列缺失突变[11],及单碱基插入和大片段缺失突变等[18,20];相继开发了不同的分子标记,包括烟草eIF4E1野生型等位基因5’端显性分子标记[9]、开阳小黑烟eIF4E1突变型等位基因共显性分子标记[17]和福泉柳叶烟eIF4E1突变型等位基因共显性分子标记[18],这些分子标记的相对位置如图1 所示。考虑到上述分子标记的检测范围未能涵盖烟草eIF4E1基因3’端,本研究另外在烟草eIF4E1基因3’端设计了烟草eIF4E1野生型等位基因3’端显性分子标记(图1)。接着本研究利用4 种分子标记引物对19 份抗PVY 资源和1 份感PVY 资源进行PCR 扩增试验,结果显示,4 种分子标记将20 份资源区分为4 个标记基因型(图2)。标记基因型A 包括开阳小黑烟、坝林晒烟、光柄柳叶(木)、一朵花、勐海乡晒烟(二)、盘县大柳叶和册亨威旁土烟共7 份抗病资源。标记基因型B 包括半坤村晒烟和K326,其中半坤村晒烟表型鉴定为抗PVY,K326 表型鉴定为感PVY。标记基因型C 只包括抗PVY 资源福泉柳叶烟,已知福泉柳叶烟eIF4E1基因3’端发生大片段缺失(部分缺失)[18]。标记基因型D 包括普安付耳烟(3)、C151、NC55、TN86、Havana 10、Criodllo Salteno 11、陕县兰花烟、灵宝莫合烟、茄子烟、建平大兰花烟共10 份抗病资源,利用4 种分子标记引物扩增均未产生任何条带,初步推测这10 份抗病资源eIF4E1基因发生了完全缺失。

图1 烟草eIF4E1 基因的4 种分子标记在基因组上的相对位置示意图Fig. 1 Relative positions of four molecular markers of tobacco eIF4E1 gene in the genome
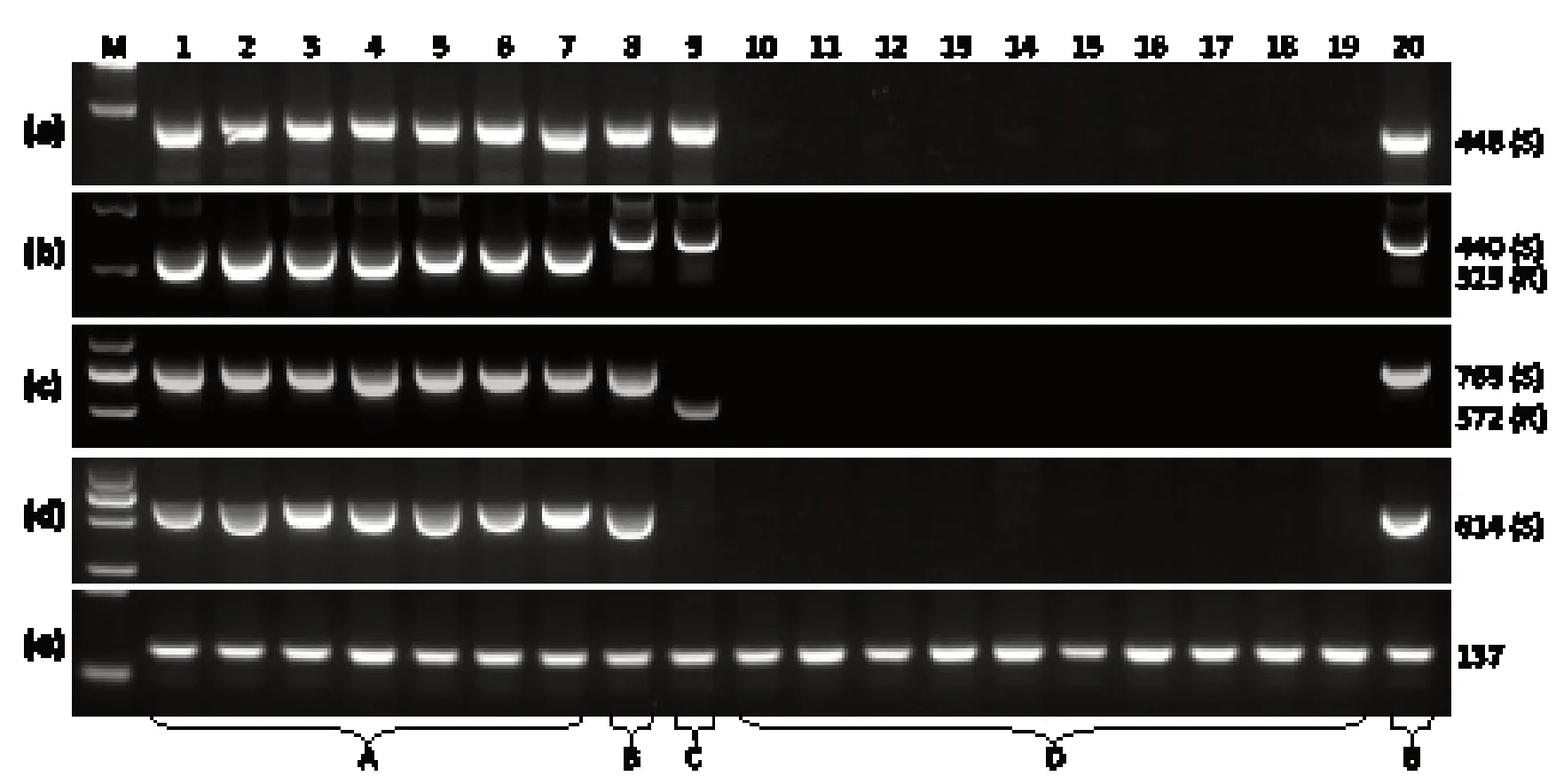
图2 抗PVY 烟草种质资源中eIF4E1 基因的分子标记检测Fig. 2 Molecular marker detection of eIF4E1 gene in tobacco germplasm resources resistant to PVY
2.4 eIF4E1 基因的RT-PCR 扩增及序列分析
应用RT-PCR 技术对19 份抗PVY 资源的eIF4E1基因进行RT-PCR 扩增和序列测定,结果显示,在福泉柳叶烟、普安付耳烟(3)、C151、NC55、TN86、Havana 10、Criodllo Salteno 11、陕县兰花烟、灵宝莫合烟、茄子烟和建平大兰花烟等11 份种质资源中未能扩增出eIF4E1基因的全长cDNA 序列(图3),已知在福泉柳叶烟中eIF4E1基因发生3’端大片段缺失(该种突变型等位基因被命名为eIF4E1.F),结合eIF4E1基因的特异分子标记检测结果,推测其他10 份种质资源中eIF4E1基因发生了完全缺失;在开阳小黑烟、坝林晒烟、光柄柳叶(木)、一朵花、勐海乡晒烟(二)、盘县大柳叶和册亨威旁土烟等7份种质资源中均能扩增出两个大小不同的条带(图3),进一步测序显示eIF4E1基因在这些资源中存在两种剪接模式(图4),其原因在于eIF4E1基因1号外显子3’端发生单一碱基(T)插入突变影响到部分前体RNA 内含子1 的正确剪接(GT-AG 法则),导致下游2 号外显子发生跳跃缺失突变,考虑到此种突变型等位基因最初在开阳小黑烟中被发现,故将其命名为eIF4E1.K[20];与上述eIF4E1基因突变类型不同,在半坤村晒烟中可以扩增出与感病品种K326 一样的单一条带(图3),测序结果显示,半坤村晒烟的eIF4E1基因编码区(CDS)第149 位碱基处存在一个等位基因突变,即由G 突变为C(图4),从而导致相应编码的蛋白质第50 位氨基酸残基由色氨酸(W)突变为丝氨酸(S),为将半坤村晒烟中发现的eIF4E1突变型等位基因与其他突变型等位基因区分开来,我们将半坤村晒烟中发现的eIF4E1突变型等位基因命名为eIF4E1.B。

图3 烟草eIF4E1 基因的全长cDNA 序列的PCR 扩增Fig. 3 PCR amplification of the full-length cDNA sequence of eIF4E1 gene in tobacco

图4 烟草eIF4E1 野生型与突变型等位基因的全长cDNA 序列比对Fig. 4 Alignment of the full-length cDNA sequences of wild-type and mutant alleles of eIF4E1 in tobacco
3 讨论
温室盆栽苗期人工接种鉴定结果,19 份烟草种质资源对PVY 表现为高抗,其中开阳小黑烟、坝林晒烟、半坤村晒烟、福泉柳叶烟、C151、NC55、TN86、Havana 10、Criodllo Salteno 11、陕县兰花烟、灵宝莫合烟、建平大兰花烟等12 份资源PVY 抗性在先前文献中已有报道[9,13,15,18,20],但除开阳小黑烟、福泉柳叶烟、C151、NC55 和TN86 等5 份种质资源外,对其他种质资源中PVY 感病基因eIF4E1的基因型未见报道。已知目前烟草育种利用的PVY 抗源可能全部衍生自烟草资源VAM,利用VAM 抗源及其衍生抗源育成的抗PVY 烟草品种包括白肋烟品种TN86、TN90 和烤烟品种NC744、NC55、NC102 等,但是在这些抗源及育成的抗病品种中往往围绕着eIF4E1基因及其侧翼序列的染色体区域发生了大范围的缺失,其中涵盖上百个基因的缺失[11]。上述染色体区域的大范围缺失及无法获得缺失连接片段序列信息不仅造成eIF4E1基因位点野生型和突变型共显性分子标记难以开发,不利于隐性性状的选择,而且也会由于一些功能基因的缺失带来不利的农艺性状。早有研究报道,VAM 基因型抗源材料的PVY 抗性存在连锁累赘,如叶片短小、产量较低、缺乏叶面分泌物等[22-23]。这些连锁累赘现象可能与eIF4E1基因上下游大量功能基因的缺失相关,因此即使针对这类抗源材料设计出共显性标记用于回交育种辅助选择或增加回交次数,也无法打破PVY 抗性定向改良带来的连锁累赘。
为克服或避开上述问题的困扰,本研究从鉴定烟草种质资源抗PVY 表型和基因型入手,通过分析发现所有筛选到的抗病资源无一例外在eIF4E1基因位点发生了不同类型的突变,其中筛选到的晒烟和黄花烟抗病资源均是我国地方品种(又称农家品种),是经过长期的自然繁衍、淘汰及人工驯化选择的结果,不涉及人工诱变或基因修饰。尽管筛选到的所有抗病农家品种均是由于eIF4E1基因位点发生自然突变造成的结果,还不能说明eIF4E1基因是烟草抗PVY 的唯一机制,但可能表明eIF4E1基因位点是目前进行烟草抗PVY 品种选育的最佳靶向位点,不会对其他农艺性状带来负面效应或带来的负面效应相对较小,同时利用筛选到的eIF4E1单个基因突变的抗病资源(如开阳小黑烟或半坤村晒烟)也可能消除或减小围绕eIF4E1基因位点发生大量基因缺失突变的抗病资源(如TN90 或NC55)作为供体亲本带来的负面效应。然而,需要指出的是,近年来,国内外学者先后在烟草上发现了多个能够突破eIF4E1基因型抗性的PVY毒株[24-27],于此同时,在烟草中发现了与之对应的新的隐性抗病基因eIF(iso)4E-T[28]。因此,今后如果能够针对两个隐性抗病基因eIF4E1和eIF(iso)4E-T进行烟草种质资源抗PVY 优异等位变异的挖掘和聚合育种,势必可以创制出赋予烟草对绝大多数PVY 病毒株系抗性的新品种[27]。
回交育种对带有个别不良性状的品种改良是一种较好的育种途径,而分子标记辅助选择(MAS)因其对目标基因的快速准确选择为回交育种提供了有效工具[29]。由于eIF4E1基因是隐性抗病基因,在以选择杂合个体为目标的回交转育过程中,以扩增eIF4E1野生型等位基因为目标的显性分子标记不能有效识别含有eIF4E1突变型等位基因的杂合个体,仍需通过逐代测交或自交的方法鉴定基因型,不尽完美。鉴于共显性的分子标记允许在回交转育杂合体阶段进行隐性基因的鉴定,无需进行测交或自交检验,节省选育时间,本研究对烟草种质资源抗PVY 表型和基因型进行鉴定,筛选到多个新的抗病资源,并获得了各个抗病资源eIF4E1基因突变序列信息,为建立烟草PVY 隐性抗病基因eIF4E1的共显性分子标记提供了可能,为提高烟草PVY 抗病育种选择效率,加快品种改良进程奠定了基础。
4 结论
本研究从900 多份烟草种质资源中筛选出19 份高抗PVY 资源,其中14 份为中国烟草地方品种资源。通过抗性基因等位性测验,初步推断19 份抗源的PVYN抗性由隐性抗病基因eIF4E1所控制。通过序列分析,确定19 份抗源中eIF4E1基因存在4 种突变类型,包括单碱基插入、替换、基因片段部分或完全缺失。这些结果为烟草PVY 抗病育种中抗病材料的选择提供了依据,也为烟草抗PVY 共显性分子标记的开发和应用提供了可能,从而有利于提高育种效率,加快育种进程。
猜你喜欢
今日农业(2022年4期)2022-06-01
智慧健康(2021年17期)2021-07-30
空间科学学报(2021年1期)2021-05-22
国际检验医学杂志(2021年7期)2021-04-15
中国产前诊断杂志(电子版)(2020年1期)2020-05-21
环境保护与循环经济(2017年5期)2018-01-22
浙江农业学报(2017年1期)2017-05-17
现代检验医学杂志(2016年5期)2016-08-20
新农业(2016年20期)2016-08-16
中国果菜(2016年9期)2016-03-01
