
不同混交比例对杉木和大叶榉幼苗功能性状的影响
2021-11-26 11:36查美琴成向荣虞木奎韩有志
生态学报 2021年21期
查美琴,成向荣,虞木奎,韩有志,汪 成,江 斌
1 中国林业科学研究院亚热带林业研究所 华东沿海防护林生态系统国家定位观测研究站, 杭州 311400 2 山西农业大学林学院, 太谷 030801 3 江西省永丰县官山林场, 吉安 331506
人工林是全球森林资源的重要组成部分,是国家重要的自然资源和战略资源[1]。现有人工林经营主要以纯林为主,导致生产力下降、地力衰退、生物多样性丧失、病虫害严重等许多生态问题[2],严重影响了人工林生态系统的可持续发展。营建人工混交林有助于提高林地生产力、促进养分循环,改善风险管理以及生态可持续性[3]。在森林生态系统中,种间关系作为决定人工混交林经营成败的关键[4],其表现形式主要有竞争、互补效应(竞争减小)和促进作用3种。通常,树种组成和混交比例是影响森林生态系统中种间相互作用关系的重要因素[5- 6]。然而,目前关于人工混交林的研究,主要集中于纯林与混交林树木生长效应、生态系统养分循环、生物生产力和生态经济效益等方面[7],尚缺乏针对人工混交林功能性状特征及其种间与种内变异,以及地上和地下性状相互作用关系的系统研究。
植物功能性状是植物在长期进化过程中适应周围环境而形成的对生态系统功能有一定影响的植物形态、生理或物候特征[8],反映了植物对不同环境的适应及植物内部不同功能之间的生理或进化的权衡,可以将环境变化同植物群落的结构和功能联系起来[9],其变异主要表现在植物主要器官形态和生理性状的差异以及营养元素的分配[10]。近年来,关于树种组成与植物功能性状或生产力之间关系的研究日益受到关注[11]。如Xiang等[12]研究表明,比根长、比根面积和细根生物量在马尾松(Pinusmassoniana)和石栎(Lithocarpusglaber)混交林中略高于各自纯林。Fujii and Kasuya[13]研究表明,在赤松(Pinusdensiflora)和日本扁柏(Chamaecyparisobtusa)混交林中,较小的赤松具有较低的细根生物量、较短的根长和较低的根尖密度,但比根长和比根尖密度高于较大的赤松。这种不同树种组成下树木功能性状的变异是由于参与树种之间相互作用的结果[14]。一般认为竞争减小和促进作用是驱动生物多样性-生态功能正相关的两个主要机制[15]。相反,当混栽树种间发生竞争时,从而至少一种对另一种产生直接或间接抑制的现象(降低增长或死亡率)[16]。然而,树木组成多样性与林分生产力之间的关系取决于立地条件、树种类型以及资源有效性。而目前关于树种组成如何影响树木功能性状的研究较少。此外,在混交林经营中,确定最佳混交比例是决定混交林产量和稳定性的重要因素[17]。如Binkley等[18]研究表明50%柳叶桉(Eucalyptussaligna)+ 50%南洋楹(Falcatariamoluccana)人工混交林地上生物量比柳叶桉纯林高25%。Santos等[19]研究表明,与巨尾桉(Eucalyptusgrandis×Eucalyptusurophylla)纯林相比,50%巨尾桉+ 50%马占相思(Acaciamangium)混交林中巨尾桉高度和胸径更大,但总生物量与巨尾桉纯林相当。混交比例对林分生产力的影响主要取决于树种组成及其种间或种内的相互作用关系。混交比例会改变林分结构,并可能影响地上、地下的树木功能性状。然而,以往研究主要关注混交比例对林分蓄积量或生物量的影响。很少涉及到混交比例对地上和地下功能性状影响的研究。
杉木(Cunninghamialanceolata)是亚热带地区主要的人工林造林树种之一。人工林面积达895万hm2,占中国人工林总面积的19.01%(第八次全国森林资源清查数据)[20]。但大多数杉木人工林仍采用集中连片的纯林经营模式,尤其是杉木连栽导致林分生产力下降、土壤质量衰退以及生态系统功能降低等诸多问题日益突出[21]。大叶榉(Zelkovaschneideriana)生长快,材质优良,是重要的乡土珍贵用材树种[22]。大量研究表明营建杉阔混交林有助于提高和改善林分生产力和稳定性。为此,本文通过控制试验研究杉木和大叶榉幼苗在不同混交比例栽培模式下叶、茎、根功能性状变化及其生物量分配特征,明确2个树种在不同混交比例下生态适应策略,为杉木+大叶榉混交林科学经营提供参考。
1 材料与方法
1.1 试验地概况
本试验地位于浙江省杭州市富阳区虎山(东经119°57′,北纬30°3′)试验基地,该地区具有典型的亚热带湿润季风气候,年平均气温16.4 ℃,年日照时数1816 h,年平均降水量1441.9 mm,无霜期为252 d。
1.2 植物材料及试验设计
本研究选取亚热带地区人工林经营常用树种杉木和大叶榉,采用1年生幼苗进行5种处理(种植)模式:4棵大叶榉幼苗种植于一个塑料花盆中(4Z),1棵杉木和3棵大叶榉(1C3Z),2棵杉木和2棵大叶榉(2C2Z),3棵杉木和1棵大叶榉(3C1Z)和4棵杉木(4C),两个树种幼苗叶、茎、根生长情况初始值见表1。每个处理重复5次,共25盆,盆栽试验在温室大棚中进行。
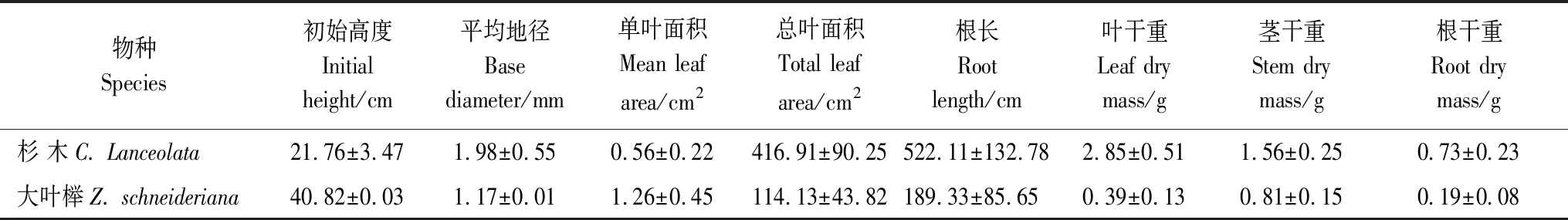
表1 杉木和大叶榉幼苗初始性状和生物量(平均值±标准差)
盆栽土壤取自天然灌木林地,是典型的红黄壤[23],风干后过1 cm×1 cm的筛网(以去除石头和植物残体)。使用前,在土壤中添加20%的泥炭土。将混合后的土壤装入塑料花盆(外茎48 cm,内茎41.5 cm,高31 cm,盆底有4个排水孔)中,每盆土20 kg。土壤全氮采用半微量凯氏法测定,水解性氮采用碱解扩散吸收法,全磷采用硫酸-高氯酸浸提-钼锑抗比色法测定,有效磷采用盐酸和硫酸溶液浸提-钼锑抗比色法测定,速效钾采用乙酸铵浸提-火焰光度法测定,土壤有机质采用重铬酸钾氧化-外加热法测定[24],采用电位法测定土壤pH值[25],土壤初始理化学性质见表2。2019年3月中旬,将一年生杉木和大叶榉幼苗按试验处理后分别种植在每个塑料花盆中。试验期间对两个树种幼苗进行相同的抚育管理, 包括定期清除花盆里的杂草和浇水。

表2 土壤样品基本特征(平均值±标准差)
1.3 光合气体交换测量
利用Li- 6400XT便携式光合分析仪(Li- 6400XT, Li-Cor, USA)对两个树种的光合气体交换参数进行测定。分别采用12 cm2(2 cm×6 cm)针叶叶室和6 cm2(2 cm×3 cm)标准叶室分别测定杉木和大叶榉光合参数。测定过程中大气温度(Ta,℃)25—28 ℃,相对湿度(RH,%)58%—62%,固定流速为500 μmol/s,叶室内CO2浓度固定在400 μmol/mol,大气CO2浓度(Ca, μmol/mol)由仪器自动记录。。光照强度使用LED红蓝光源,设定为1000 μmol m-1s-1的光合有效辐射(PAR)。所有测量分别于2019年10月15—19日9:00—11:00进行。每个处理中,每盆中的每个树种分别选择1株幼苗,每棵幼苗选择3片相对健康和成熟的叶子进行测量。待光合作用稳定后2 min记录净光合速率(Pn, μmol m-2s-1)、蒸腾速率(Tr, mmol m-2s-1)和气孔导度(Gs, mmol m-2s-1)。瞬时水分利用效率(WUE, μmol/m)采用Pn/Tr计算。
1.4 形态性状的测量
2019年10月下旬对所有处理进行整株收获,按叶、枝干和根系分别放入贴有标签的自封袋中,放入装有冰袋的保温箱内。在每个处理中,对各盆中的每个树种随机选择20片相对健康和成熟的叶片作为一个亚混合样本进行叶面积测量。每个样本的单叶面积(MLA, cm2),总叶面积(TLA, cm2),根长(RL, cm)通过根扫描设备(WinRHIZO, Canada)和软件(Leaf and root image analysis software)进行测定。在鲜重和饱和鲜重称量之后,将所有的叶片、枝干和根系样本小心地放入纸袋中,在60 ℃下烘干至恒定质量,然后测定其干重。比叶面积(SLA, cm2/g)为叶面积/叶干重(LM, g)比值。叶片干物质含量(LDMC, g/g)计算为叶干重/叶饱和鲜重(LSFM, g)比值。比根长(SRL, cm/g)计算为根长/根干重(RM, g)比值[26]。本文中的MLA、TLA、SLA、LDMC、RL、SRL、LM、SM、RM、TMPP、LMR、SMR、RMR和R/S代表每个处理中每个幼苗的平均值。
1.5 数据分析
采用WPS和SPSS 25.0软件对数据进行整理和分析。不同栽培模式下杉木、大叶榉叶、茎、根形态、生理功能性状和生物量及其分配的差异采用SPSS 25.0软件的单因素方差(one way ANOVA)分析,并使用邓肯多重比较法(Duncan′ multiple test)进行不同处理之间差异性检验分析(P<0.05)。不同栽培模式下杉木、大叶榉叶、茎、根功能性状及生物量及其分配的相关分析(Pearson correlation analysis)采用R 3.6.3软件中的“corrplot”包完成[27],并使用R 3.6.3软件中的“ggplot2”, “ggrepel”, “plyr” 和 “vegan”对其进行主成分分析(Principal component analysis,PCA)。
2 结果与分析
2.1 叶片性状变化
杉木单叶面积在不同处理间没有显著差异(0.667),总叶面积随杉木在树种组成中所占比例的降低而减少,比叶面积在混栽模式下显著增大(P<0.05),叶干物质含量在混栽模式下显著减小(P<0.05)(表3)。大叶榉单叶面积在3C1Z处理下最高,总叶面积随大叶榉在树种组成中所占比例的降低而逐渐增大,比叶面积在不同处理间无显著差异(0.758),叶干物质含量在2C2Z处理下显著高于其它3个处理(P<0.05)。
杉木净光合速率、蒸腾速率和气孔导度在混栽处理下显著低于4C处理(P<0.05),而瞬时水分利用效率在不同处理之间差异不显著(0.543)(表4)。大叶榉净光合速率、蒸腾速率、气孔导度和瞬时水分利用效率在不同处理之间均差异显著(P<0.05),其中,净光合速率、蒸腾速率和气孔导度均在2C2Z处理下最大,而瞬时水分利用效率在2C2Z处理下最小。
2.2 根系性状变化
杉木根长和比根长在不同处理之间无显著差异(P=0.408、0.387)(图1)。大叶榉根长随其在树种组成比例的减小呈增大趋势,且3C1Z处理下根长显著高于其它3个处理(P<0.05),比根长在不同处理间差异不显著(P=0.130)。
2.3 不同处理对生物量的影响
杉木叶、茎、根生物量和单株总生物量在混栽模式下均显著低于4C处理(P<0.05),且叶、茎、根生物量和单株总生物量在混栽模式下随杉木在树种组成中所占比例的增加而增加(图2)。大叶榉叶、茎、根生物量和单株总生物量随大叶榉在树种组成比例的增加而逐渐减小,且叶和茎生物量在3C1Z和2C2Z处理下均显著高于1C3Z和4Z处理(P<0.05),而1C3Z和4Z处理间差异不显著(P>0.05)。根生物量和单株总生物量在3C1Z处理下显著高于其它3个处理(P<0.05),而根生物量在2C2Z、1C3Z和4Z 3个处理间差异不显著(P>0.05)。不同栽培模式下各处理总生物量差异显著(P<0.05),2C2Z处理下总生物量最高,4C处理下总生物量最低,而4Z、1C3Z和3C1Z3种处理间总生物量无显著差异(P> 0.05)(图3)。
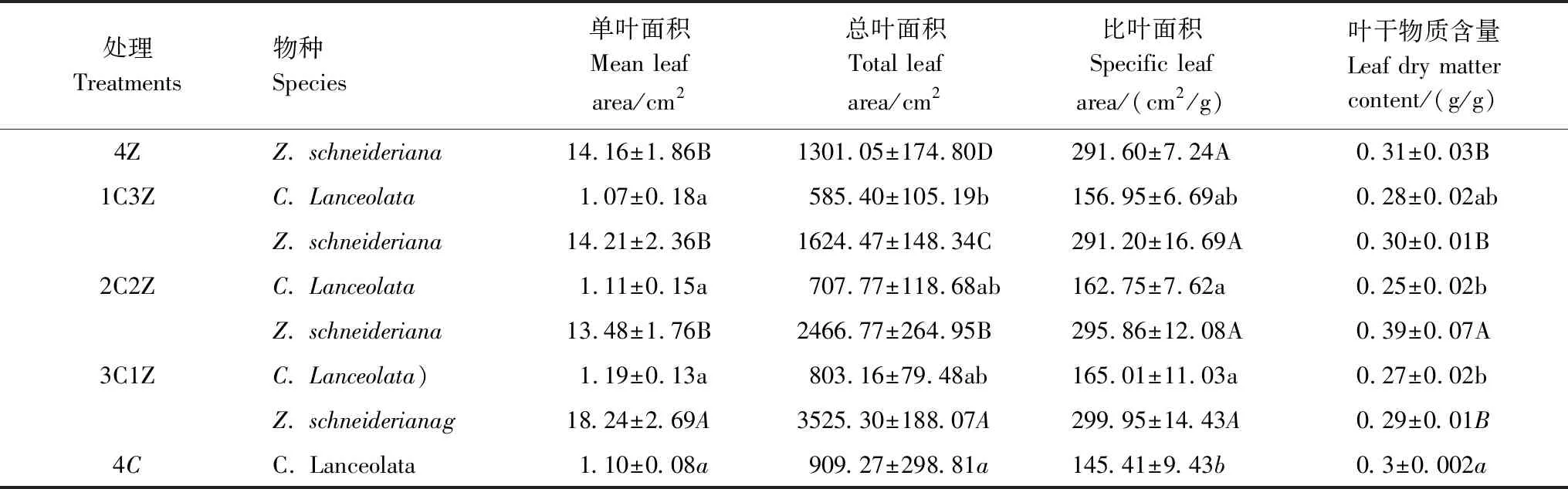
表3 不同处理下杉木和大叶榉叶形态性状的变化

表4 不同处理下杉木和大叶榉叶光合性状的变化
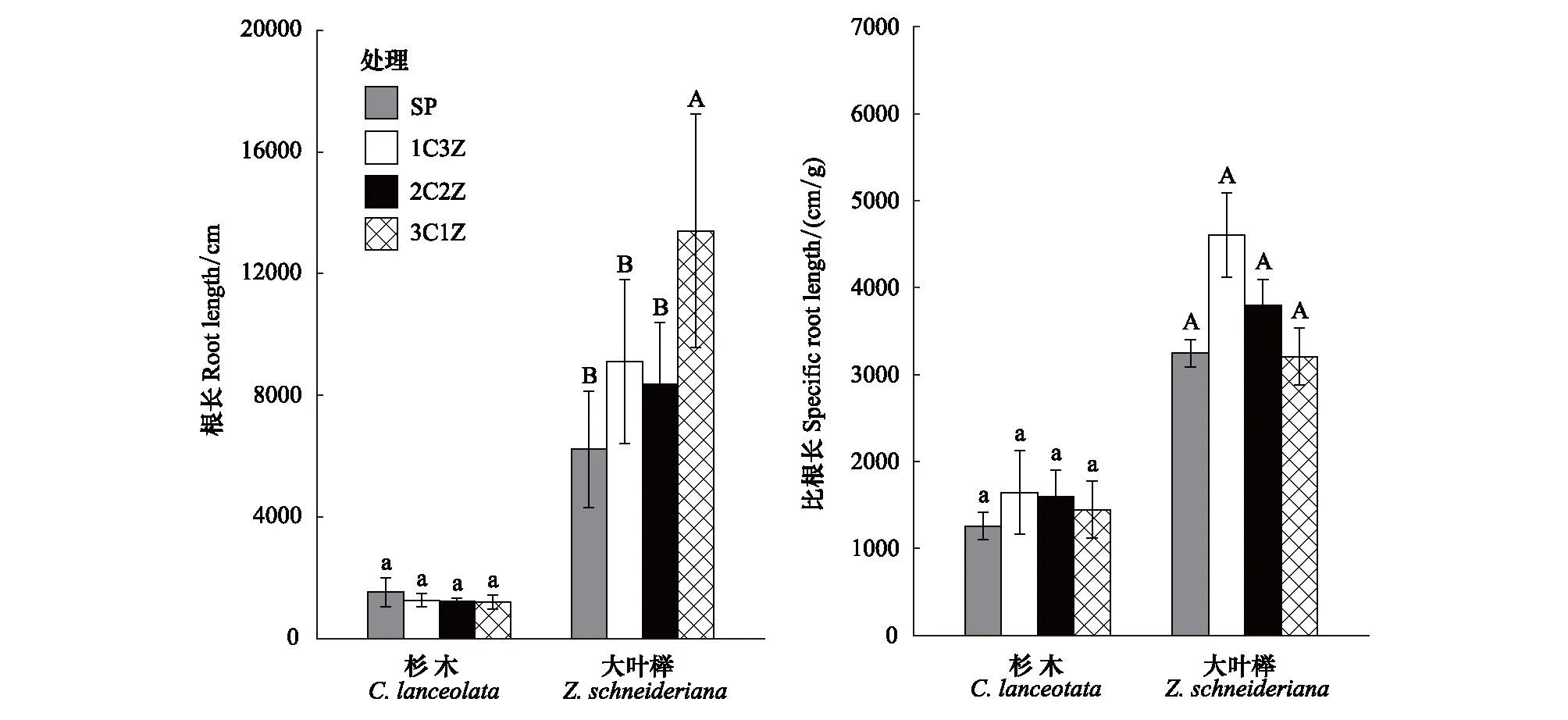
图1 不同处理对杉木和大叶榉根系形态性状的影响Fig.1 Effects of different treatments on root morphological traits of C. lanceolata and Z. schneideriana 误差条表示均值的标准差(n=5);在同一情况下,不同字母表示不同处理间在 P<0.05水平上差异显著,其中杉木的为小写字母,大叶榉的为大写字母;SP:4株杉木或大叶榉幼苗 four C.lanceolata seedlings or four Z.schneideriana seedlings;1C3Z:1株杉木和3株大叶榉幼苗 one C.lanceolata and three Z.schneideriana seedlings;2C2Z:2株杉木和2株大叶榉幼苗 two C.lanceolata and two Z.schneideriana seedlings;3C1Z:3株杉木和1株大叶榉幼苗 three C.lanceolata and one Z.schneideriana seedlings
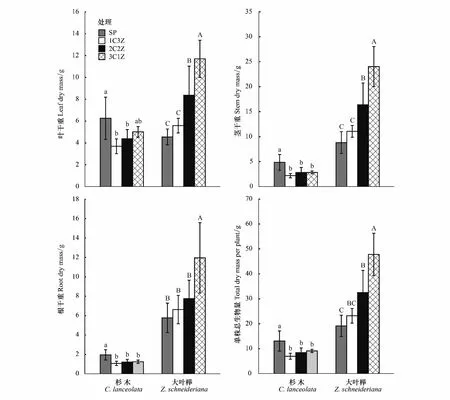
图2 不同处理对杉木和大叶榉叶、茎、根及单株总生物量的影响Fig.2 Effects of different treatments on leaf, stem, root and total biomass per plant of C. lanceolata and Z. schneideriana误差条表示均值的标准差(n=5);在不同字母表示不同处理间在 P<0.05水平上差异显著,其中杉木的为小写字母,大叶榉的为大写字母
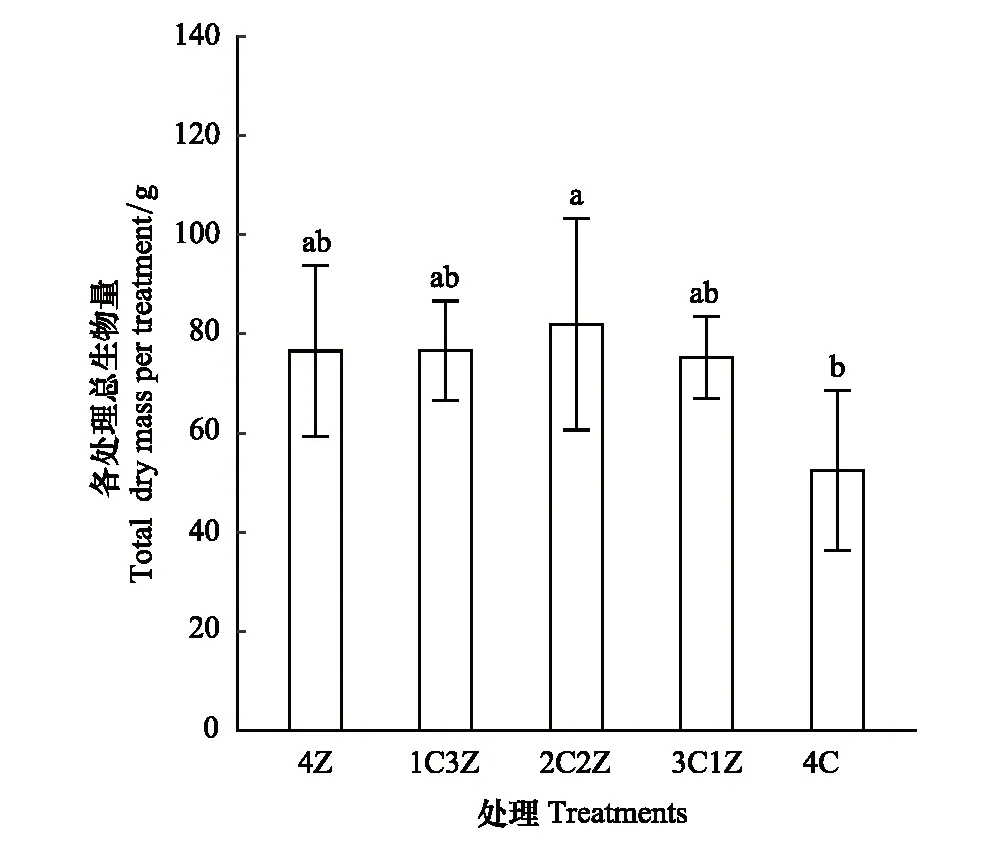
图3 不同处理对杉木和大叶榉各处理总生物量的影响 Fig.3 Effects of different treatments on total biomass per treatment of C. lanceolata and Z. schneideriana 误差条表示均值的标准差(n=5);在同一情况下,不同小写字母表示不同处理间在 P<0.05水平上差异显着;4Z:4株大叶榉幼苗 four Z. schneideriana seedlings;4C:4株杉木幼苗 four C. lanceolata seedlings
不同处理对杉木、大叶榉叶、茎、根生物量比和根冠比影响较大(表5)。杉木叶生物量比、根生物量比和根冠比在不同栽模式之间无显著差异(P=0.943、0.496、0.704),茎生物量比在4C处理下最大,且显著高于1C3Z和3C1Z混栽处理(P<0.05)。大叶榉叶生物比在不同处理间无显著差异(0.809),茎生物量比随大叶榉在树种组成中所占比例的降低而逐渐增大,且在3C1Z和2C2Z处理下显著高于4Z处理(P<0.05),根生物量比和根冠比随大叶榉在树种组成中所占比例的降低而逐渐减少,且根生物量比和根冠比在3C1Z和2C2Z处理下显著低于4Z处理(P<0.05)。
2.4 杉木和大叶榉幼苗叶、茎、根功能性状及生物量分配的相关分析
杉木和大叶榉叶、茎、根功能性状与生物量及其分配间的关系相似(图4)。总叶面积和根长分别与叶生物量、茎生物量、根生物量和单株总生物量呈显著正相关(P<0.05),且总叶面积与根长也分别呈显著正相关性(P<0.05)。但两个树种的功能性状与生物量及其分配间也表现出不同的相关性。杉木净光合速率、蒸腾速率和气孔导度与比叶面积呈显著负相关(P<0.05)。净光合速率和气孔导度与叶生物量、茎生物量、根生物量和单株总生物量呈显著正相关(P<0.05),而比根长与茎生物量、根生物量和单株总生物量呈显著负相关(P<0.05)。大叶榉单叶面积与叶生物量、茎生物量、根生物量和单株总生物量呈显著正相关(P<0.05)。总叶面积和比叶面积与茎生物量比呈显著正相关(P<0.05),而与根生物量比和根冠比呈显著负相关(P<0.05)。
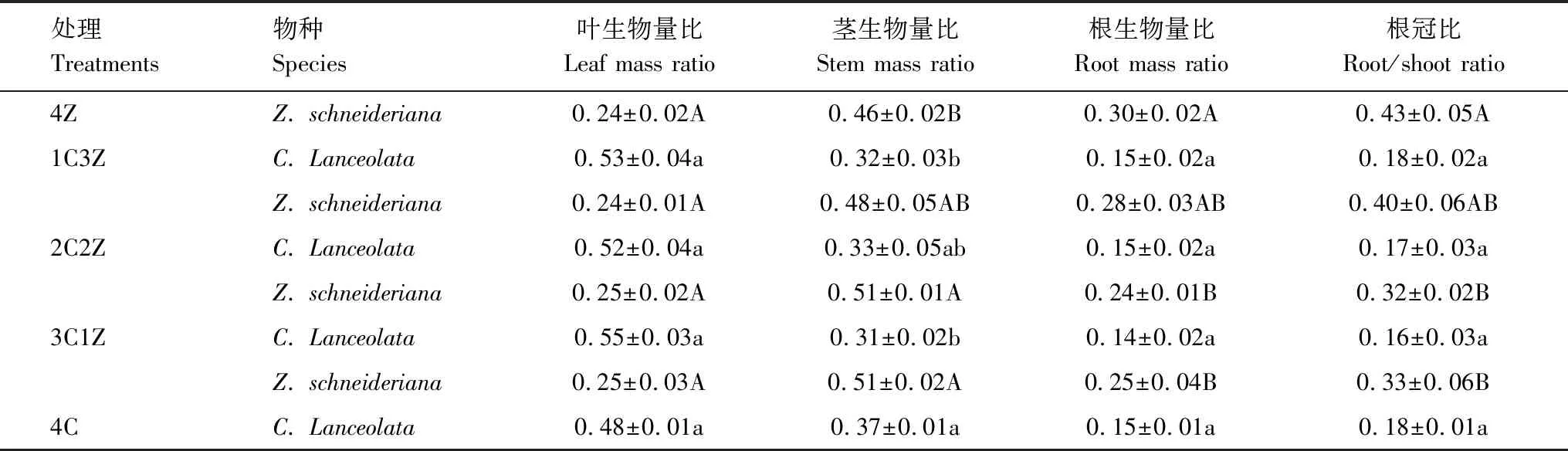
表5 不同处理下杉木、大叶榉叶、茎、根生物量比和根冠比的变化
分别基于不同栽培模式下杉木、大叶榉叶、茎、根功能性状和生物量及其分配的主成分分析(PCA)表明(图5),杉木前2个主成分可以解释杉木功能性状和生物量及其分配变异总方差的95.81%,其中PC1可以解释总方差的80.96%,PC2可以解释总方差的14.85%,总叶面积、比叶面积、根长和比根长贡献较大;大叶榉前2个轴占数据变异性的97.53%,其中PC1和PC2轴分别解释总方差的61.94%和35.59%,也是总叶面积、比叶面积、根长和比根长贡献较大。
3 讨论
叶片是植物进行光合作用的主要场所,也是对外界环境变化最敏感的器官,植物对环境变化的多数响应主要体现在叶性状上,其性状的变异能够反映树种在生态策略上的差异[28]。本研究发现杉木总叶面积和叶干物质含量在混栽模式下显著低于4C处理,而比叶面积显著增大;大叶榉总叶面积随其在树种组成中所占比例的降低而逐渐增大,这表明不同栽培模式对两个树种叶功能性状影响较大。Qin等[11]研究表明,马尾松(Pinusmassoniana)单叶面积和比叶面积在混交林和纯林之间无显著差异,叶干物质含量在混交林中显著高于纯林。Forey等[29]发现山毛榉(Fagussylvatica)叶干物质含量沿树木丰富度的梯度显著降低。此外,李淑英[7]研究发现,杉木+闽楠混交林中闽楠叶面积和比叶面积显著提高,而杉木比叶面积减小。Qin等[30]研究发现白榆(Ulmuspumila)和刺槐(Robiniapseudoacacia)的比叶面积在不同混交比例栽培模式中差异显著,而单叶面积和叶干物质含量无显著差异,且混栽模式能有效提高两个树种叶片的光合能力。这些不同的研究结果表明树种混交对林木叶性状影响的复杂性,取决于树种、林龄、混交比例、立地条件等因素。本研究中,大叶榉生长速率高于杉木,混栽模式生长后期大叶榉枝叶对杉木造成一定程度的侧方遮阴,杉木有效光照显著降低导致其净光合速率显著下降,杉木通过增大比叶面积来增强对光能的捕获效率。尽管杉木比叶面积随混合比例的减少显著增大,但总叶面积逐渐下降,导致整株的受光面积逐渐减少,这将影响杉木在混栽模式下光合产物的累积,进而导致叶片干物质含量下降。这与成向荣等研究结果类似[31]。而大叶榉在混栽模式下较4Z处理相比改善了冠层结构,为大叶榉的生长提供了更多的空间资源,混栽模式下随大叶榉比例的减少,总叶面积呈增大趋势。
根系是植物从土壤中获取水和矿质养分的主要器官,其性状特征能够有效反映植物应对竞争和土壤有效性的适应策略[32]。本研究发现杉木根长在不同处理间无显著变化,而大叶榉根长随大叶榉在树种组成比例的减小呈增大趋势。这表明混栽模式提高了大叶榉根系对土壤资源的利用能力。姚甲宝等[33]研究发现,木荷(Schimasuperba)与杉木混栽时两个树种的根系总长度高于单植。Xiang等[12]研究报道称马尾松和石栎(Lithocarpusglaber)混交林细根根长密度高于纯林。相对于纯林,混交树种的根系可塑性、林龄、树种组成以及根系种间竞争等将对混栽树种根系功能性状产生显著作用。本研究中,混栽模式下杉木、大叶榉根长相对变化的差异主要是由两个树种种间资源竞争能力不同导致的,相对于杉木,大叶榉根系获取土壤资源的能力更强,在土壤资源竞争方面处于优势地位,混栽模式有利于促进其对土壤资源的利用。杉木、大叶榉比根长在不同处理间均没有显著变化,这一方面可能与试验处理中土壤养分和水分供应充足有关,混栽模式下杉木、大叶榉根系不需要维持较高的比根长来增强其对土壤资源的吸收效率[34]。另一方面可能是试验周期太短,两个树种在不同处理间对土壤资源利用的竞争效应尚未体现。
生物量是评价生态系统结构与功能的重要指标。Bauhus和Schmerbeck[35]对混交林研究发现生物多样性与生产力之间呈显著正相关。另一些研究表明物种丰富的林分生产力高于物种贫乏或单一树种栽培的林分[36- 37]。混交林生产力的提高可能归因于竞争减少和促进作用[38- 39]。如Bauhus等[3]报道称在50%的蓝桉(Eucalyptusglobulus)+ 50%的黑荆(Acaciamearnsii)混栽处理中树干材积和生物量最高。然而,Bouillet等[40]研究发现马占相思在50% Acacia+50% Eucalypts和25% Acacia+100% Eucalypts混栽模式下对巨桉生长无抑制作用,但巨桉在50% Acacia+100% Eucalypts混栽模式下的生物量产量较100% Eucalypts单一树种栽培相比减少了21%。不同混交比例对混栽树种生产力的影响主要取决于特定立地条件下种间竞争和促进过程之间的平衡。本研究中,混栽模式下杉木叶、茎、根及单株总生物量较4C处理相比均显著减少,而大叶榉却随其在树种组成中所占比例的减少而增加。这与两个树种在混栽模式下对地上、地下资源利用和种间竞争有关。生物量分配能够反映植物获取资源的能力。本文中大叶榉茎生物量比在混栽模式下显著增大,而根生物量比在混栽模式下显著减小,这与王振兴等研究结果相似[41],可能是大叶榉为了提高自身竞争力,分配更多的资源供给地上部分来获得更多的光照资源[42]。根冠比反映了植物对环境因子需求和竞争能力,植物的地上生物量较大,表明植物对地上资源具有较强的需求和竞争能力。本研究中,杉木、大叶榉在不同栽培模式下根冠比均<1,这表明杉木、大叶榉地上生物量较大,两个树种均对地上资源具有较强的竞争能力。且大叶榉随其在树种组成中所占比例的降低而对地上资源的需求和竞争越强。相比较而言,大叶榉对地下资源的需求和竞争能力强于杉木,而杉木对光具有较强的竞争能力。

图4 不同处理下杉木、大叶榉叶、茎、根功能性状和生物量分配的相关分析Fig.4 Correlation analysis of leaf, stem, root functional traits and biomass allocation of C. lanceolata and Z. schneideriana among different treatments**表示在 P<0.01水平相关性显著,*表示在 P< 0.05水平相关性显著;n = 5; MLA: 单叶面积 Mean leaf area, TLA: 总叶面积 Total leaf area, SLA: 比叶面积 Specific leaf area, LDMC: 叶干物质含量 Leaf dry matter content, Pn: 净光合速率 Net photosynthetic rate, Tr: 蒸腾速率 Transpiration rate, Gs: 气孔导度 Stomatal conductance, WUE: 瞬时水分利用效率 Instantaneous water use efficiency, RL: 根长 Root length, SRL: 比根长 Specific root length, LM: 叶干重 leaf dry mass, SM: 茎干重 Stem dry mass, RM: 根干重 Root dry mass, TMPP: 单株总生物量 Total dry mass per plant, LMR: 叶生物量比 Leaf mass ratio, SMR: 茎生物量比 Stem mass ratio, RMR: 根生物量比 Root mass ratio, R/S: 根冠比 Root/shoot ratio
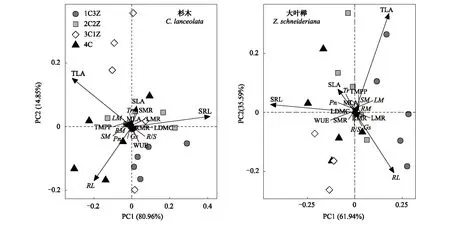
图5 不同处理下杉木、大叶榉叶、茎、根功能性状和生物量分配的主成分分析Fig.5 Principal component analysis (PCA) of leaf, stem, root functional traits and biomass allocation of C. lanceolata and Z. schneideriana among different treatments
植物各功能性状不是独立发挥作用的,而是通过一系列性状组合的调整和平衡以适应特定生境[43],甚至是不同群落间、群落内乃至功能群内的种内与种间功能性状均表现出普遍的相关性与变异性[44]。本研究中杉木、大叶榉总叶面积和根长与叶生物量、茎生物量、根生物量和单株总生物量分别呈显著正相关性。这表明生物量的变化取决于地上和地下资源的获取。通常,较大的总叶面积可以拦截更多的光或更有效地转换拦截辐射。同时,根长越大,越完整地占用土壤空间和获取土壤资源。相反,总叶面积和根长越小,其对地上的光和/或地下土壤体积探测能力越弱。因此,本研究中,混栽模式下种间竞争产生取决于两个树种功能性状可塑性的差异,且这种功能性状可塑性的差异对混栽模式下两个树种生物量生产的影响起决定性作用,亦是混栽树种大叶榉生长竞争优势形成的原因之一。从总体上看,杉木、大叶榉叶、茎、根功能性状和生物量及其分配之间的显著正相关性表现出较高的多性状协同适应特征,但不同栽培模式下的变化特征也存在一定的差异。Kerkhoff等[45]研究表明,植物叶-根功能性状间的相关性与植物所在生境的自然环境及植物性状的表型可塑性等密切相关。所以,生长在同一栽培模式下杉木、大叶榉叶、茎、根功能性状和生物量分配之间的权衡关系均会存在差异。PCA排序结果显示了不同栽培模式下两个树种叶、茎、根功能性状和生物量及其分配的变化特征具有一致性。
4 结论
本文探讨了杉木、大叶榉叶、茎、根功能性状在不同栽培模式下的变异特征,研究发现不同混交比例对两个树种的叶、茎、根功能性状的影响不同。这表明植物功能性状的变化不仅受非生物环境的影响,而且与树种混交比例密切相关。与相应单一树种栽培相比,杉木、大叶榉在混栽处理中生物量的变化部分原因可能是由两个树种功能性状可塑性差异造成的,混栽模式下两个树种对地上光和地下土壤资源的种间竞争可能是其生物量生产的主要驱动因素之一。因此,根据经营目标,确定适宜的混交比例对人工林的可持续经营具有重要意义。本文仅通过模拟试验开展了两个树种幼苗期功能性状随混交比例变化的研究,树种组成对林木生长和功能性状的长期影响还有待进一步研究。
猜你喜欢
小小说月刊·下半月(2021年8期)2021-09-05
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
农民致富之友(2020年8期)2020-05-11
新疆农业科学(2020年1期)2020-02-14
动漫界·幼教365(大班)(2018年3期)2018-05-14
中央民族大学学报(自然科学版)(2017年4期)2017-06-11
湖南大学学报·自然科学版(2016年12期)2017-05-12
绿色科技(2016年15期)2016-10-11
江苏农业科学(2016年4期)2016-06-14
中国麻业科学(2015年5期)2015-12-28
