
曲靖早泥盆世西屯组软骨鱼(雅致郭氏鱼)鳞片形态学与组织学研究
2021-12-08 08:55彭礼健
曲靖师范学院学报 2021年6期
彭礼健,李 强
(曲靖师范学院 自然历史文化研究中心,云南 曲靖 655011)
0 引 言
早泥盆世是早期脊椎动物各类群演化的关键时期,这一时期早期脊椎动物许多类群在多样性和数量上都呈现爆发式的增长,占据了繁多的生态位.云南曲靖地区早泥盆世地层自下而上分别是西山村组、西屯组、桂家屯组和徐家冲组[1],这套地层是中国早泥盆世地层的经典剖面.其中,西屯组鱼化石尤为丰富,保存了无颌类和有颌类等众多类群的化石,特别是硬骨鱼类一些原始类型的化石,如先驱杨氏鱼、罗氏斑鳞鱼、张氏蝶柱鱼、晨晓弥曼鱼等[2-6],对于研究早期硬骨鱼类的演化具有极为重要的意义.然而,相对于硬骨鱼类的研究来看,该生物群中软骨鱼类群的研究工作尚属薄弱环节,相关研究一直没有系统开展.
软骨鱼类是内部骨骼只有软骨组成的有颌类脊椎动物,其头部为完整的软骨脑颅,身被细小的鳞片[7-11].软骨鱼类最早的的化石记录是奥陶纪时期蒙古鱼类牙和鳞片[12],蒙古鱼因其复杂的多齿状鳞片而为人所知,多见于北美、中国和蒙古[13],这表明软骨鱼在海洋中的辐射早于古生代的其它鱼类(图1).而全球软骨鱼的躯干化石非常稀少,只有两块发现于志留纪,大多发现于早泥盆世[14-18].古生代软骨鱼类的主要发现集中于亚洲中、东部(华南、塔里木、蒙古西部、西伯利亚和图瓦地区等)[19-25]、波罗的海周边[26]、加拿大北部[27-30]、玻利维亚[31]以及澳大利亚[32-33]等地,多为分散保存的鳞片与鳍刺,并有少量的牙齿材料.
软骨鱼自身的内骨骼不易保存,但因其牙齿不断地产生、脱落和替换以及周身覆盖细小鳞片的身体特征,使得牙齿和鳞片大量保存[34-35].由于化石材料的缺失,软骨鱼类化石的真正归属大多存疑,很多无法纳入到相关类群的系统发育分析中.除此之外,软骨鱼类如何由盾皮类逐渐演变而来?软骨鱼类和硬骨鱼类是如何分化的?过去认为的很多软骨鱼的特征是否为有颌类的原始特征,还是软骨鱼的进步特征?这些问题的解决都急需补充更多的软骨鱼化石材料,以进一步完善相关类群的系统发育.
产自云南曲靖早泥盆世西屯组的软骨鱼类—雅致郭氏鱼是为数不多的产自中国的早泥盆世软骨鱼类代表.前期,已有学者对其形态学进行了初步研究[36-37],但对其组织学和生长模式均未做深入探讨.本文利用高精度计算机断层扫描(CT)、三维重建、电镜拍照、组织学切片等多种手段对雅致郭氏鱼进行更详细准确的形态学与组织学研究.
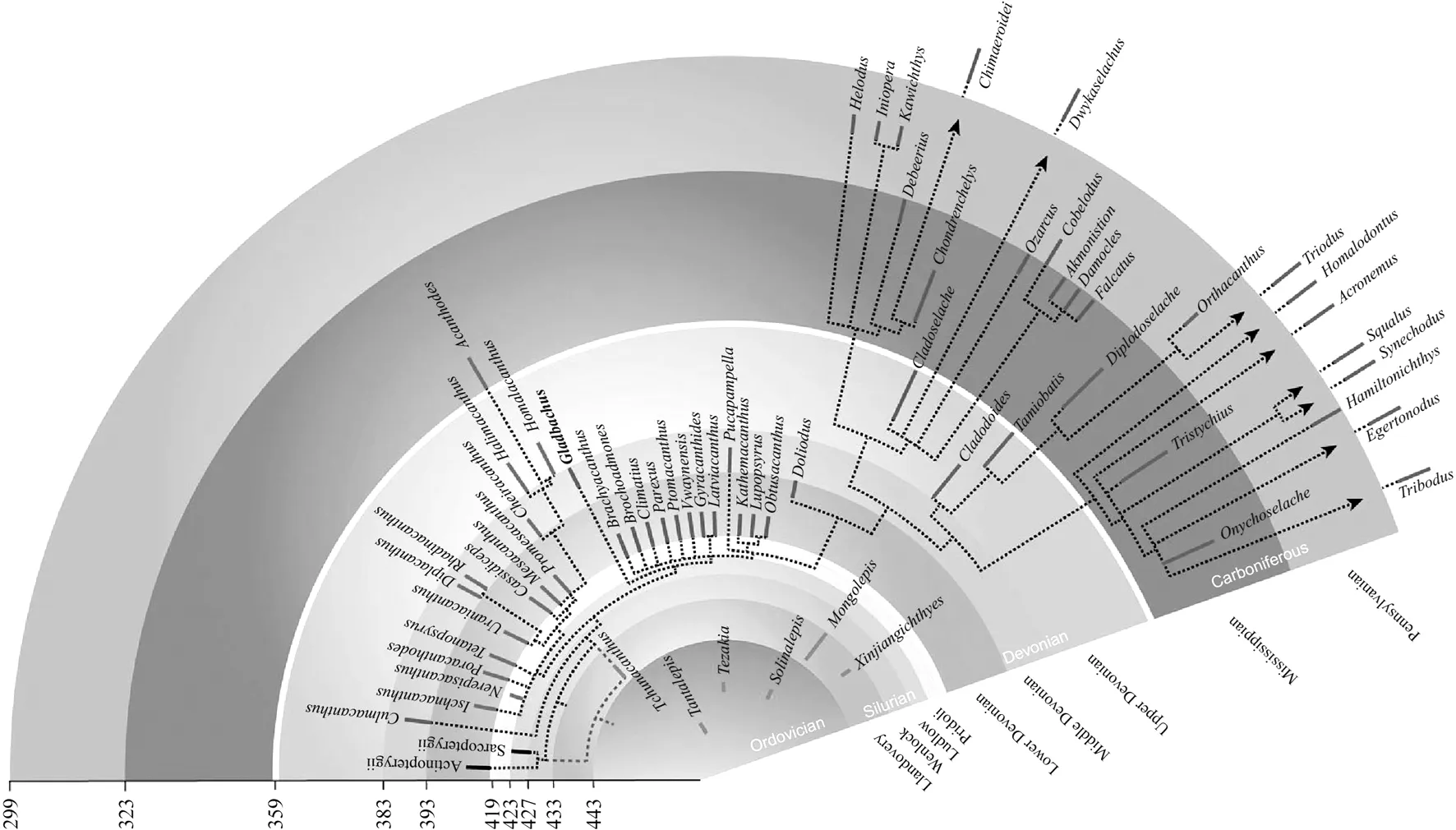
图1 早期软骨鱼类系统发育关系(修改自Michael Coates,2018[19],图中标红部分为已报道的早泥盆世软骨鱼类化石)
1 材料和方法
1.1 研究材料
本文的研究材料为雅致郭氏鱼鳞片化石,均取自云南曲靖泥盆纪西屯组.目前所有鳞片均保存于曲靖师范学院自然历史文化研究中心.
1.2 研究方法
1.2.1 采样及酸处理
将野外获取的岩样进行破碎处理,碎小后的岩样放入塑料容器内,配置3%的醋酸溶液倒入其中没过岩样,待反应至气泡不再产生.取出初次反应的上层清液留作缓冲液,并将反应过的岩样置于缓水流下12 h.反应过水后留下的细小岩样用30目、60目、100目的网筛梯度过滤并分装烘干.将烘干后的样品倒入分液漏斗,加入重液分选1~2 h,利用密度差分离围岩和微体化石.将分选出的微体化石再次冲洗、烘干在体视显微镜下挑选,分类和保存.
1.2.2 CT扫描和三维重建
运用中国科学院古脊椎动物与古人类研究所的高精度CT设备(225 Micro CT)对挑选出的雅致郭氏鱼鳞片进行扫描.扫描后得到的slice数据,导入VGStusio软件进行调整后导成RAW文件通过Mimics Research 18.0软件进行三维重建.重建完成的雅致郭氏鱼类型1鳞片的3D图像见图2.
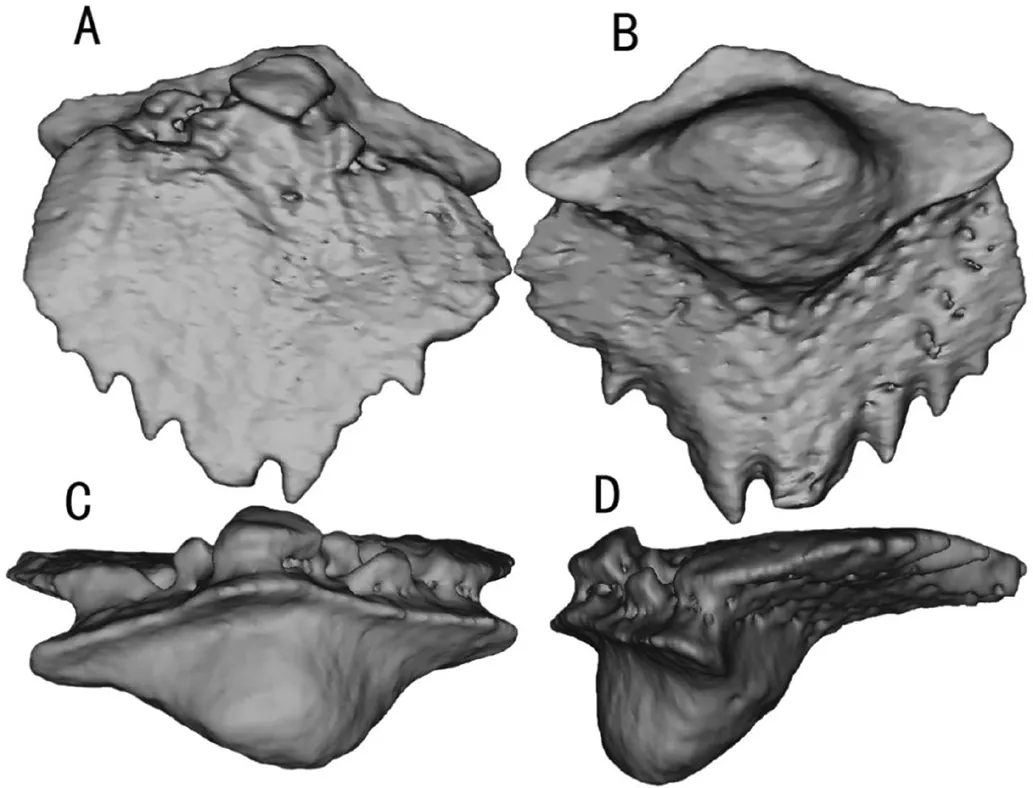
图2 重建完成的雅致郭氏鱼类型1鳞片的3D图像
1.2.3 扫描电镜拍照
扫描电镜(Scanning Electron Microscope简称SEM[38])设备来自中国科学院古脊椎动物与古人类研究所.鳞片化石材料本身是非导电品,为了获得效果更好的图片,进行SEM拍照前需在样片表面涂抹金粉增强导电性.
1.2.4 制作组织学切片
挑选出用于切片制作的鳞片显微拍照并编号.在软胶模具底部先铺一层3 mm厚的环氧树脂,将模具放入真空干燥箱中抽真空,并加热使树脂固化.将鳞片固定在固化树脂中央,再倒入环氧树脂将鳞片完全覆盖,继续抽真空、烘干过程.树脂完全固化后在精密切割机上对鳞片进行分割,将需要观察的一面打磨抛光用树脂粘在单面全磨砂载玻片上,切掉多余部分,继续打磨抛光,整个磨抛过程需借助偏光显微镜观察切片厚度.切片在偏光显微镜下能清晰看到内部组织结构即为完成,借助偏光显微镜完成组织学拍照工作
2 研究内容
2.1 形态学
雅致郭氏鱼的6种类型鳞片的SEM图见图3.

图3 雅致郭氏鱼6种类型鳞片的SEM图
类型1 这些鳞片是对称或近乎对称的.冠部扁平而薄,呈菱形,有几个同心的脊状纹饰,纹饰从上表面前部延伸至后方逐渐消失.冠部的前侧边缘光滑圆润,而冠的后侧边缘有5~10个锯齿.有些鳞片的后外侧边缘呈弱锯齿状,甚至是光滑的,但都有一个或两个突出的后尖.在冠部的前部,有些鳞片具有三角状的次生齿列,在前方重叠.冠部向后延伸超出基部.冠部的下表面中部有几排小孔,从中间到边缘孔径逐渐变小.冠部下表面的后外侧边缘突出,没有小孔.每个鳞片的颈部较低,有许多比冠的下表面孔大的小孔.基部在鳞片的前部,菱形,宽而短.基部的前侧边缘从中部到边缘逐渐变薄,并向前延伸至冠部外侧.基部的后半部分比前半部分厚,中间部分隆起.基部外侧端形成锐角,向外侧伸展.有些鳞片的基部是扁平的,有一个髓腔开口.
类型2 这些鳞片的冠部呈叶状,几乎对称.冠部的上表面有几个在后面逐渐消失的同心脊,如形态类型1.冠部前侧边缘光滑圆润,后侧边缘微凹,呈锯齿状或光滑.冠部有一个或两个突出的后端,类似于形态类型1.在冠部的前缘有三角状的次生齿列向前嵌套.冠部的下表面中部有几排小孔,从中间到边缘逐渐变小.冠部下部下表面中部有几排小孔,比形态类型1的小孔更稀疏、更小.
类型3 这些鳞片有一个扁平的梯形冠部,冠部上表面有几个从后面逐渐消失的同心脊.冠部的长度大于其宽度,前侧边缘光滑,有次生齿列,而冠部的后侧边缘光滑或有锯齿.冠部向后延伸到基部,形成一个突出的尖端.冠鳞片有一个低而明显的颈部,具一排孔.基部是长方形或梯形的,它们是扁平的或微微膨胀的.与形态类型1和2不同,形态类型3的基部完全被冠部覆盖.没有发现有这种形态的髓腔开口的鳞片.
类型4 这些鳞片有一个不对称的心形冠部,后有一个突出的尖端.冠部上装饰有同心的脊状纹饰.宽度线几乎垂直于长度线,就像形态类型1.前侧缘有一些沟.在冠的下表面中间有成排的小孔.颈部很低,但非常狭长,有一排孔.基部为菱形或梯形,膨大或有横向脊,所述横向脊限于所述基部的前凹部和后凸部.这种形态的鳞片,在基上开有髓腔.
类型5 这些鳞片与形态型3相同,有一个平坦的梯形冠部,上面有几个在后面逐渐消失的同心脊.长前外缘比短前外缘长得多.在前外缘中部可见几个次级齿列.冠部的后外侧边缘有明显的锯齿.冠部下表面中部有稀疏的小孔.颈部低而窄,有一排孔.扁平的基部是长方形的.这种形态比形态类型3的基部更窄.与形态类型3相似,没有发现具有这种形态的鳞片有髓腔开口.
类型6 这些鳞片有一个菱形的扁平冠部,上面有非常模糊的同心脊.冠部前缘是沟状的,而后缘是没有锯齿的.冠的下表面比其他形态类型窄,只有不到三排的小孔.这些鳞片的颈部很低,有一排小孔.菱形的基部扁平或肿胀.基部的长度线与部冠的长度线几乎平行.这类鳞片基部没有髓腔开口.
2.2 组织学
以上六种不同形态学的鳞片具有一致的组织学特征(图4),证明其属于同一个属种.雅致郭氏鱼鳞片冠部由齿质组成,具有垂直于生长线的齿质小管,齿质小管大致平行,有的呈分枝状.齿质中有2~6条生长线,在冠部的后部非常明显.这些生长线大致与冠部的下表面平行.冠部有上升的血管通道,有些是分叉的,延伸到冠后部的齿质中.在冠部的前部,有密集的上升或水平的血管通道.基部由细胞骨组成,含细胞陷窝的分布不均,通常在中部和外部较多.发达的沙普氏纤维很长,呈放射状从上部延伸到基部的外层.

图4 雅致郭氏鱼鳞片纵切组织学图
2.3 生长模式
在以前的研究中,雅致郭氏鱼的鳞片包括幼年、成年和老年三个生长阶段,在大小和形状上各不相同[36].然而,通过对雅致郭氏鱼更多鳞片标本进行更细致的研究,总结出雅致郭氏鱼的生长模式(见图5),发现其生长过程比此前所知道的更加复杂.此外,一些较小的鳞片,具有非常薄的基部或较大的髓腔开口,没有次生齿列,可能代表了真正的幼年鳞片[36].根据鳞片不同的大小、形态和组织学特征,雅致郭氏鱼鳞片的个体发育可分为四个阶段.
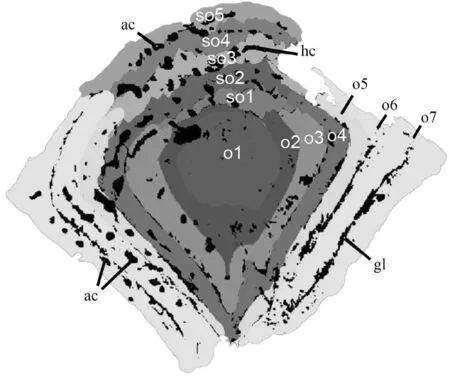
图5 雅致郭氏鱼鳞片生长模式图
阶段1 幼年鳞片:第一阶段鳞片小而薄,冠长0.35~0.60 mm,冠宽0.30~0.60 mm,鳞片深度约0.1 mm.冠部光滑,在前缘没有次级齿列.颈部和基部几乎不存在.
阶段2 青年鳞片:冠部的生长通过增加新的同心排列的初级齿列,主要分布在冠部的下部、后部和侧面生长.次级齿列出现并向前堆积在前缘.基部向下和周围延伸,有一个延伸的髓腔.基部内表面可见叠状结构,是基部生长的痕迹.在基部的前缘可见血管开口.与此同时,颈部出现并变得突出.基部髓腔开口增大至最大.
阶段3 成年鳞片:鳞片冠部的生长通过增加初级和次级齿列生长.基部的后缘开始向前逐渐变厚,直到髓腔开口关闭.刚开始,扩大部分的基础呈球状,有许多小洞.
阶段4 老年鳞片:鳞片基部继续增厚,其中髓腔逐渐变小,最终消失.所以,这个阶段的鳞片有一个更突起的基底.
3 讨 论
鳞片特征是鉴别和分类早期有颌脊椎动物的有效特征.通过对软骨鱼鳞片组织学的研究发现,鳞片冠部由几层齿质构成,一般来说,软骨鱼鳞片的生长主要有两种模式:叠加生长(新齿质纵向生长,可增加冠部厚度)和平面生长(新齿质向水平方向生长,增加冠部的面积).尽管如此,一些鳞片通过增加新的齿列部分覆盖了前代齿列,这种模式被称为“部分叠加生长”.因此,软骨鱼的鳞片有四种生长模式:叠加生长、部分叠加生长、面积生长和同心面积生长.通过对软骨鱼鳞片同步辐射显微切片,利用齿列的分布重建了它们的三维生长模式.这些鳞片显示了一种复合模式,包括面积生长模式、叠加生长模式和部分叠加生长模式,因为它们具有第一代或后代的多个齿列.根据系统发育分析,鳞片的齿列具有更复杂的生长模式.它的第一代齿列呈线性排列,并与随后几代齿列的面积生长和部分叠加生长模式重叠.第一个齿列几乎完全被第二个齿列所覆盖,并与其他齿列以区域生长模式重叠.几乎所有的棘鱼类都表现出区域生长模式.与软骨鱼鳞片相比,棘鱼的鳞片非常小且厚,表现出一种假想的生长模式(洋葱状).但棘鱼也具有面积生长模式,通过向外侧添加新的齿列和向外侧扩展基部生长.新的系统发育证据根据区域生长模式和某些形态将该物种重新划分为软骨鱼类.所有现存的软骨鱼都有单齿鳞片,但大多数的软骨鱼都有不同生长方式的齿列.雅致郭氏鱼鳞片的三维数据表明,它们具有同心的区域生长模式.这种生长模式在某些其他软骨鱼中也同样存在.
4 展 望
在曲靖早泥盆世西屯组中发现的雅致郭氏鱼鳞片是我国最早报道的软骨鱼微体,对软骨鱼研究有着重要意义.软骨鱼微体化石也是地层研究的重要佐证,在新疆地区早志留红色岩系中发现的丰富软骨鱼微体化石[23]证明志留纪-泥盆纪时期的鱼类主要生活在滨海环境中[36].随着对越来越多含鱼地层的深入研究,研究者发现微体化石不仅存在于陆相的碎屑岩中,更多的是存在于碳酸盐地层中,而碳酸盐地层中富集大量海相化石,这一发现将早期鱼类生活环境拓宽至滨海、浅海和半深海.因而微体化石也成为判断海相-非海相对比的重要依据.未来软骨鱼微体的研究也将倾向于在建立曲靖早泥盆世软骨鱼鳞片详细分类研究的基础上,与不同地区不同时代的软骨鱼进行对比建立更完整的微体化石序列进而建立软骨鱼类生物带.另一方面,软骨鱼作为从奥陶纪发展留存至今的具有完整演化体系的物种,对软骨鱼的研究是了解生命演化的重要证据,所以建立从奥陶纪-志留纪-泥盆纪到石炭纪-二叠纪-三叠纪系统的软骨鱼类微体研究是有必要的.
猜你喜欢
野生动物学报(2022年1期)2022-02-24
作文大王·低年级(2020年4期)2020-04-19
阅读与作文(小学低年级版)(2019年12期)2019-12-26
百家讲坛(2019年16期)2019-04-30
百家讲坛(2019年18期)2019-02-16
百家讲坛(红版)(2017年11期)2018-01-20
热带农业科学(2017年9期)2017-10-23
农村农业农民·B版(2016年7期)2016-10-21
