
黔西南石漠化区不同群落土壤碳氮磷垂直分异及化学计量特征
2021-12-14 04:49刘子琦喻阳华李开萍蔡路路
西南农业学报 2021年10期
余 逍,刘子琦,喻阳华,李开萍,蔡路路
(贵州师范大学 喀斯特研究院/国家喀斯特石漠化防治工程技术研究中心,贵州 贵阳 550001)
【研究意义】生态化学计量学结合了物理、化学、生物等多学科的基本原理,是研究生态系统能量平衡与碳(C)、氮(N)、磷(P)等多重化学元素平衡的一种理论[1]。C是构成植物体干物质最主要的元素,N在生物合成、光合作用方面起着重要作用,P是核酸和酶的重要组成部分[2]。C、N、P影响着植物生长发育和各种生理代谢过程,而且C与N、P元素循环过程是相互耦合的,三者密切相关[3]。土壤C、N、P化学计量是衡量土壤有机碳(SOC)分解速率、元素限制判断和土壤营养平衡的重要指标[4-5]。从土壤化学计量的角度揭示不同群落土壤生态系统内部C、N、P平衡与循环过程,对于深入认识喀斯特石漠化生态系统具有重要意义。【前人研究进展】目前,诸多学者开展了人工植物群落土壤生态化学计量学的研究。郭其强等[6]研究贵州高原山地马尾松(Pinusmassoniana)人工林下0~30 cm土壤C∶N特征表明,人工林土壤C∶N远低于我国土壤C∶N均值,反映了人工林下物种多样性增加使得土壤矿化有机碳能力增强;刘立斌等[7]分析喀斯特次生林下0~30 cm土壤C、N、P化学计量特征结果表明,喀斯特森林土壤C含量高,C∶N和N∶P高于我国平均水平,N、P元素匮乏;王璐等[8]研究得出,喀斯特高寒干旱区核桃(Juglansregia)人工林0~30 cm土壤C∶N高于刺梨林(Rosaroxburghii),核桃地保肥能力更好,种植核桃有利于改善生态环境;陶慧敏等[9]研究云南喀斯特生境下松林(Pinusyunnanensis)、赤杨林(Alnusjaponica)和侧柏林(Platycladusorientalis)0~10 cm土壤养分化学计量表明,人工林土壤C∶N略低于全球森林土壤均值,表明云南喀斯特地区土壤有机碳矿化、分解能力较好;李善家等[10]认为,内蒙古黑河下游流域的黑果枸杞(Lyciumruthenicum)群落土壤N∶P低于14,植物生长受N限制;杨慧等[11]利用C、N、P化学计量学的内稳态理论,揭示了岩溶生态系统次生林土壤C、N、P化学计量受地貌部位影响显著;陶冶等[12]分析香樟(Cinnamomumcamphora)C、N、P化学计量特征,揭示其生长受N素限制,为香樟可持续经营提供了思路。【本研究切入点】以上研究主要集中常态生境或喀斯特高原、山地生境下的人工植被,关于喀斯特石漠化干热河谷生境的人工植物群落土壤化学计量特征研究较少,不利于理解喀斯特石漠化生态系统植物-土壤相互作用的养分调控机制;同时,目前研究主要集中在耕作层,不利于探讨喀斯特石漠化生态系统养分循环过程及系统稳定机制。黔西南干热河谷石漠化区北盘江小流域是典型的纯碳酸盐岩峡谷环境[13],境内鲜有连片土被,该区植物具有石生、高钙、根系深扎的特点。为揭示黔西南干热河谷石漠化区植物群落土壤C、N、P垂直分异及化学计量特征,对耕作层以下土壤予以研究尤为必要。【拟解决的关键问题】选择撂荒后长期种植、经营模式相似、坡向一致的4种人工恢复植物群落进行研究,并以荒地为对照,探讨不同群落的土壤C、N、P空间分布与生态化学计量特征;揭示土壤C、N、P和化学计量的分布规律及其与群落的关系;阐明土壤剖面的养分限制状况,旨在为石漠化治理与植被恢复重建提供理论和实践依据。
1 材料与方法
1.1 研究区概况
研究区位于贵州省安顺市关岭县和黔西南州贞丰县交界处的北盘江小流域峡谷两岸(105°36′30″~105°46′30″E、25°39′13″~25°41′00″N),属珠江流域,地貌组合形态主要有溶沟、漏斗、落水洞、峰丛、峰林、岩溶洼地等,地下水位深切。喀斯特面积45.38 km2,占总面积的87.91%。地貌类型主要为喀斯特峡谷,地势起伏较大,海拔450~1450 m。属亚热带季风性湿润气候,冬季温暖干旱,夏季炎热多雨,年均降雨量1052 mm,主要集中在5—8月,年均气温18.4 ℃,年活动积温为6542.9 ℃。区内以石灰岩和泥灰岩为主要的成土母质,土壤以石灰土为主,平均土层厚度30~40 cm,局部区域可达60 cm以上。
地带性植被为针阔混交林,但受到人为活动的干扰,原生林基本被破坏,现主要以次生林为主。原生植物主要有珍珠荚蒾(Viburnumfoetidum)、铁线莲(Clematisflorida)、芒(Miscanthussinensis)、火棘(Pyracanthafortuneana)等为主的藤、灌、草;人工种植的有花椒(Zanthoxylumbungeanum)、火龙果(Hylocereuspolyrhizus)、金银花(Lonicerajaponica)、玉米(Zeamays)以及香椿(Toonasinensis)、清香木(Pistaciweinmannifolia)、构树(Broussonetiapapyrifera)等经济作物。
1.2 研究方法
基于样点布设代表性和空间均匀性原则,于2018年11月选取4种典型人工恢复植物群落,并以荒地作为对照,共5种类型作为研究对象。在每种类型内,选取海拔、坡向、坡度和土壤类型近似,植被高度、密度相似的植物群落设置样点(表1)。每种类型设置3个10 m×10 m的样方,样方之间的间距>10 m。每个样方挖掘1个剖面,共计15个剖面(5种类型×3个样方)。
取样时,尽量避开裂缝且样点距离植株>0.5m,以避开人为施肥对样品的影响。去除凋落物层和腐殖质层,自上而下按0~5、5~10、10~15、15~20、20~30、30~40 cm 6个层次分层采样,每一层次按照S型多点取样混合均匀后,采用四分法保留约1 kg鲜样带回实验室。土样带回室内剔除石粒和根系等杂物,铺设于一张A3纸上,风干后采用四分法按对角线均匀选取200 g形成一个待测样品,研磨过筛后测定SOC、TN、TP含量。
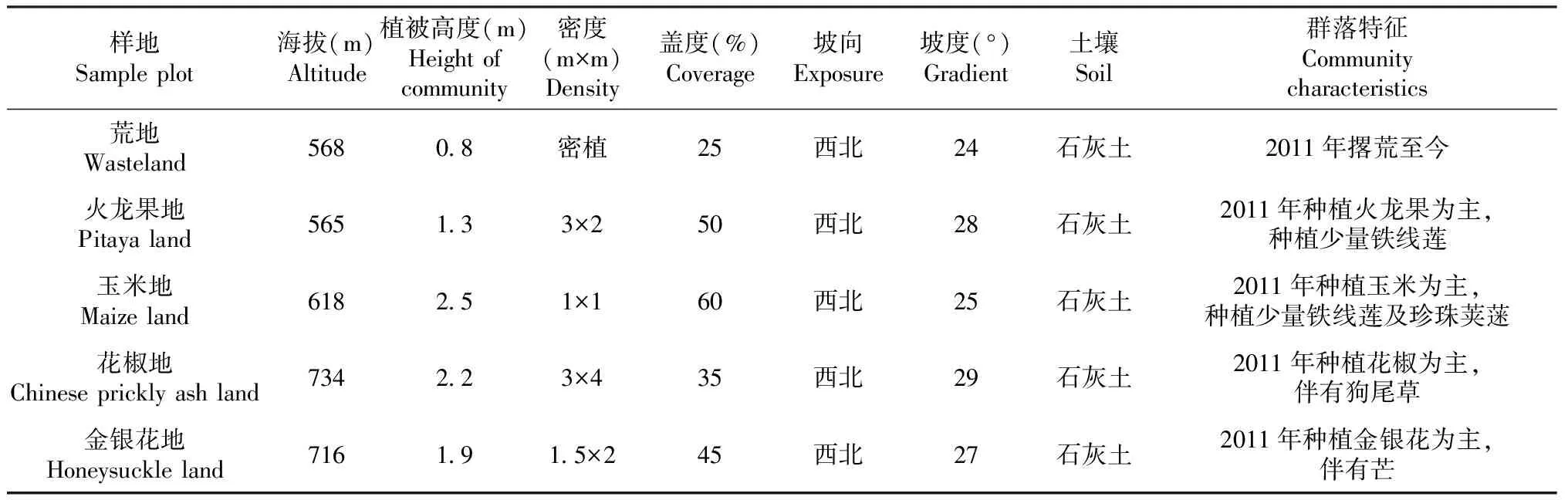
表1 样地群落基本情况
土壤TN采用硫酸消煮-凯氏定氮法测定,TP采用碱熔-钼蓝比色法测定,SOC采用重铬酸钾氧化-外加热法[14]。
1.3 数据处理
试验数据采用Excel 2016和SPSS 22.0软件进行分析处理。C、N、P的化学计量以质量比表示。采用单因素方差分析(One-way ANOVA)法比较不同群落的养分含量及其化学计量差异,用双因素方差分析检验影响土壤养分含量的主要因子。对土壤养分含量及其化学计量之间的相关性进行Pearson相关分析,图、表数据为平均值±标准差。用Origin 9.0软件绘图。
2 结果与分析
2.1 不同群落土壤C、N、P含量的垂直分异特征
从表2和图1综合看出,SOC变幅为11.98~32.91 g/kg,总体随土层深度增加而降低;花椒地和玉米地SOC含量均显著高于同一层次的荒地;各植物群落SOC在0~10与30~40 cm间均存在显著差异。TN含量的变化范围为1.98~4.99 g/kg,总体随土层深度增加而降低;玉米地、花椒地、金银花地的土壤TN含量均高于同一层次的荒地。TP含量的变化区间为0.43~1.69 g/kg,随土层深度增加变化趋势总体降低,但变化差异不显著。土壤TP含量在各群落样地间存在显著差异,玉米地和花椒地的土壤TP含量均显著高于同一层次的荒地。
2.2 土壤C、N、P含量的效应
土壤全量养分受土层深度、植被类型及其交互效应影响而发生变化,效应量大小各不相同,效应量(φ)的范围在0~1,φ<0.1表示小效应,0.1<φ<0.5表示中等效应,φ>0.5表示高效应。双因素方差分析结果(表3)表明,土壤OC、TN受土层深度、植被类型及其交互作用均达极显著水平;土层和植被的交互作用对TP影响不显著,植被、土层对TP影响则分别达极显著和显著水平。SOC效应量排序为土层、植被>交互作用,SOC效应量均达高效应;土壤TN、TP的效应量排序均为植被>土层>交互作用。
交互作用达显著影响时,需要进行简单效应分析,进一步探究各因素在各水平的影响程度。由表4可知,土层深度和植被类型对SOC交互作用中,不同土层深度均达极显著水平,效应量大小排序为0~5 cm>5~10 cm>10~15 cm>15~20 cm>20~30 cm>30~40 cm;不同植被类型均达到极显著水平,效应量排序为玉米>火龙果、花椒>金银花>荒地。对土壤TN含量的交互作用中,6个土层深度均达到极显著水平,效应量排序为5~10 cm>10~15 cm>0~5 cm>15~20 cm>20~30 cm>30~40 cm;在不同植被类型中,荒地和玉米地的作用达到显著水平,各水平因素的效应量<0.5,其余植被类型均达极显著水平。
2.3 不同群落土壤C、N、P生态化学计量特征
从表5看出,C∶N变化范围为4.64~7.87,各群落土壤剖面上无明显变化规律;同一群落中,除荒地和金银花地0~5、5~10 cm外,各群落土壤C∶N在各土层间无显著差异;不同群落间,火龙果地C∶N均显著高于同一层次的金银花地。C∶P波动范围是11.49~33.98,各群落间,火龙果地、玉米地、荒地的C∶P在15~20 cm处均有增加;金银花地的C∶P在各层次间均不存在显著差异。N∶P变幅为1.91~5.28,在土壤剖面上无明显变化规律。从整体上看,荒地和火龙果地的土壤N∶P显著高于其余群落样地。同一群落中,除花椒地、火龙果地外,其余群落的土壤N∶P在各土层间的差异均未达到显著性水平;不同群落间,花椒地土壤N∶P在各层次中(除0~5 cm外)则为最低。
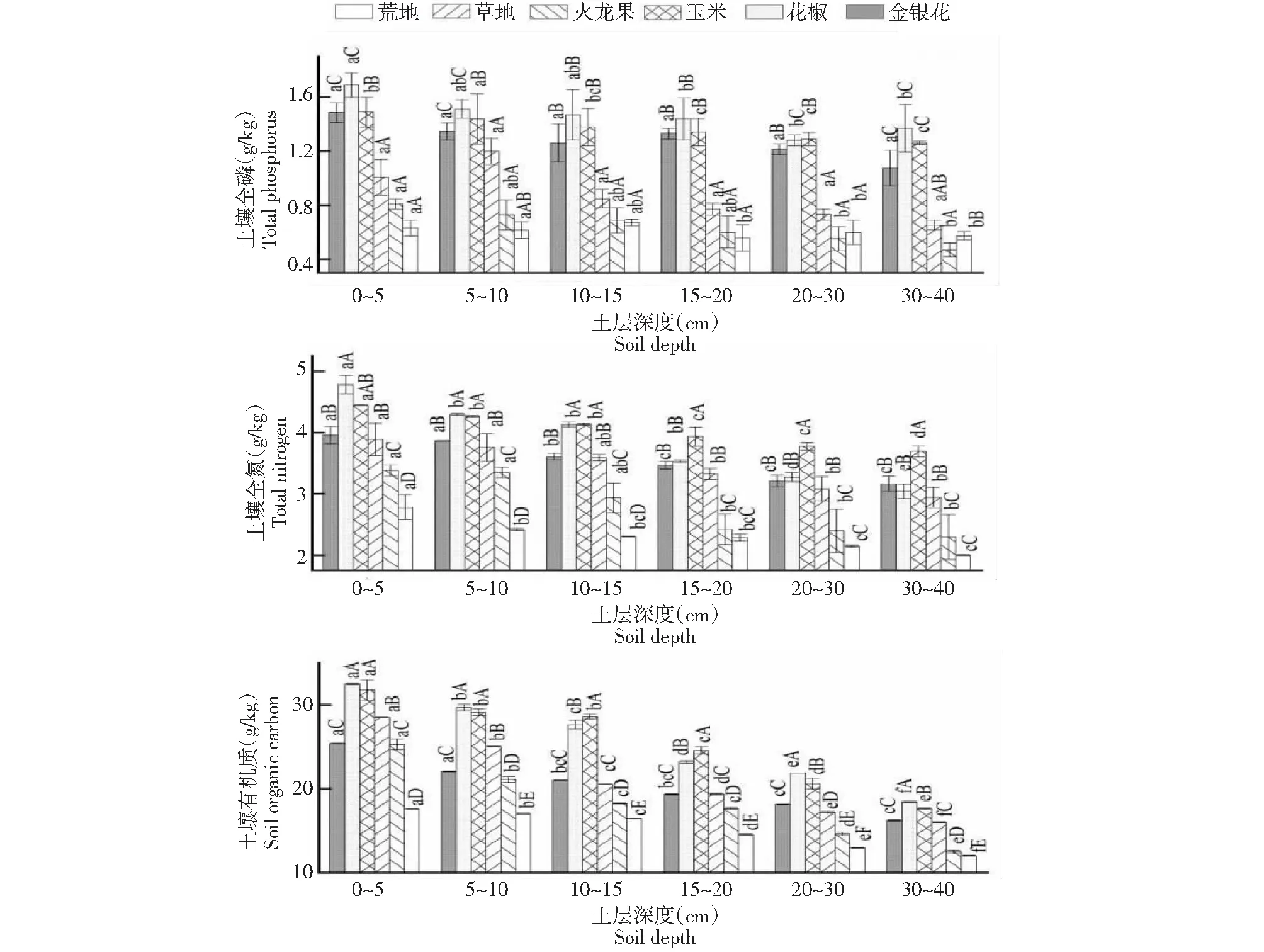
不同小写字母表示不同土层之间差异显著 (P<0.05),不同大写字母表示不同植被之间差异显著 (P<0.05),下同 Different lowercase and capital letters indicate significant difference between different soil depths at P<0.05 level.The capital letters indicate significant difference between different communities P<0.05 level.The same as below图1 不同群落样地各土层的C、N、P含量Fig.1 C,N and P content of different soil layers in different vegetation communities
2.4 土壤C、N、P含量及其化学计量之间的相关性
由表6看出,SOC、TN、TP两两之间均存在极显著正相关,表明养分全量之间具有一定的耦合关系。C∶P与C∶N和N∶P间存在极显著正相关,表明土壤生态系统内养分平衡具有较强的正向调控机制。C∶P与TN间存在显著负相关,验证了C、N之间存在互馈效应。SOC与N∶P之间为反向作用效应(相关系数r=-0.446)表明C制约着N、P元素的比例关系。
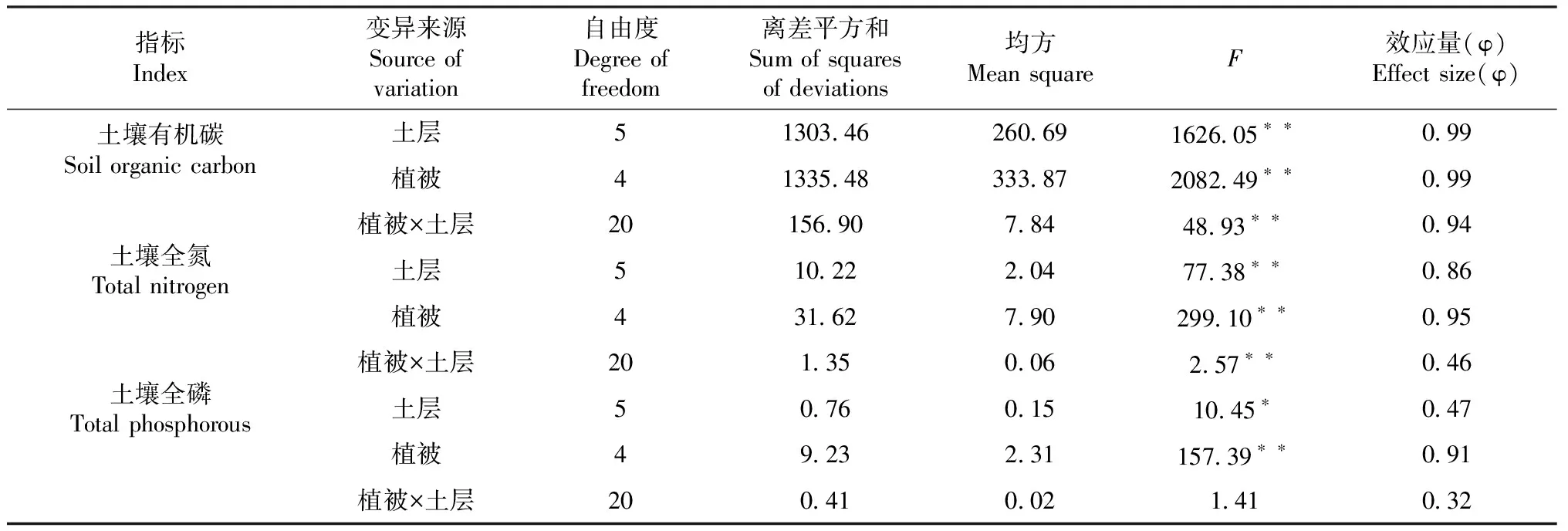
表3 不同土层深度、植被类型及其交互作用的主效应分析结果

表4 交互作用的简单效应检验分析结果
3 讨 论
3.1 不同群落土壤C、N、P含量垂直分异规律
土壤C、N、P是影响养分循环和系统稳定机制的重要化学元素[3]。本研究人工恢复植物群落SOC、TN高于黄土丘陵[15]、喀斯特峰丛洼地[16]、南方山地丘陵[17]、滦河口湿地[18]等区域(表7),原因是研究区地处干热河谷,为亚热带季风性湿润气候[13],降水充沛,水热条件优于上述地区,更有利于动物定居与植物定植,以及腐殖质积累,生物自肥作用强烈[19],但SOC、TN仍低于全国平均水平,表明研究区土壤受N限制的可能性较高。研究区TP含量高于全国平均水平,亦高于其他区域(表7),推测与研究区纯碳酸盐岩有关。Göransson等[5]研究表明,碳酸盐岩能缓慢固定来自基岩中的P,碳酸盐岩越纯,固定P的量越多。

表5 不同植被群落土壤C、N、P化学计量比特征

表6 土壤C、N、P含量与化学计量之间的相关性分析结果
SOC、TN随土层的加深表现由高及低的垂直分布格局,且表层显著高于底层,呈表聚效应。SOC随土层深度增加降低的幅度最大,原因一是表层土壤是根系活动的主要区域,根系分泌物及根际微生物对SOC的积累主要聚集在表层,植物细根通常随土壤深度的增加而减少[6,20],随土层深度增加,根系活动减弱,积累SOC的能力降低;二是凋落物和细根残体主要集中在表层,凋落物和细根残体经土壤微生物和动物分解形成的腐殖质和矿质养分优先在表层积累,然后随水流下渗或者其他介质向下层迁移扩散[8,21],使得SOC从表层到底层逐渐降低。TP在土层间无显著变化,原因是 P是一种沉积型的矿物,主要受土壤母质影响[22],在土壤中迁移速率较低。但不同群落土壤之间的TP含量变异系数达35.71%,原因可能是不同植物根系分泌的有机酸种类存在差异[3,22],导致对母岩中P元素的提取能力各异,影响了TP在各群落间的空间分布。

表7 黔西南石漠化区和其他区域的SOC、TN、TP及化学计量
人工恢复植物群落的SOC、TN、TP含量均高于荒地,原因是荒地的植被稀疏,较低的盖度致使降雨、地表径流直接作用于坡面土壤,导致其地表侵蚀加强,造成养分的迁移[11,21],加速了C、N、P等元素的分解与流失,加之荒地的物种组成、结构与层次较人工恢复植物群落单一,凋落物向土壤的养分归还量低于人工恢复植物群落,致使其土壤全量养分低于人工恢复植物群落。
3.2 土层深度、植被类型交互影响作用
土壤养分含量受植被类型、土层深度显著或极显著影响,效应类型为植被类型>土层深度>交互作用,说明植被类型是影响土壤养分分异的主要因子,其次是土层深度,最后为二者的交互效应,这是因为不同植被覆盖类型、凋落物归还形式及归还数量存在差异,使其土壤养分贮藏潜力存在本质差异[21,26]。而土层深度间存在差异,主要是土壤中水盐运动、物理沉积、渗透以及根系效应等作用引起[20,27],因此土层深度虽然对土壤全量养分影响显著,但却低于植被类型。对交互作用进一步分析发现,植被类型中,人工恢复植物群落的效应均大于荒地,表明人工恢复植物群落固存土壤养分能力较强,以0~5、5~10 cm土层的作用最强,20~30 cm的最小,原因是表层土壤与地表直接接触,累积新鲜土壤OC、TN、TP[7],作用显著,而底层土壤由地表向下逐层降解减少,积累能力变弱,因此形成不同差异。
3.3 不同群落土壤C、N、P化学计量特征及其生态学暗示
C、N、P是按照不同比率存在于生物体中的3种生源元素,其间比率的变化决定着生物和生态系统的主要特性[4]。土壤C∶N影响着微生物分解SOC的速率,从而影响植物生长发育[25]。在一定范围内,C∶N低则有利于微生物对SOC的矿化,SOC分解、矿化速率较高;反之,则微生物在分解SOC的过程中则易存在元素受限[2,25]。因此,土壤C∶N通常被认为是土壤SOC矿化能力的标志[28]。研究区土壤C∶N均值远低于全国平均水平[25],亦低于黄土丘陵[15]、荒漠草原[23]、鄱阳湖沙化土地[24]等区域(表7),表明研究区SOC分解、矿化速率均较高,这可能与研究区的亚热带季风性湿润气候有关。C∶N在土壤剖面上差异不显著,相较SOC和TN的空间变异性而言,C∶N比值相对稳定,这与前人研究结果一致[5,8,11],反映了结构性元素C、N对外界环境变化的协同响应。
土壤C∶P能表征土壤P素的矿化能力,也是衡量微生物矿化SOC释放P或从环境中吸收固持P潜力的指标[29]。贾宇等[30]研究认为,当土壤C∶P<200时,土壤中微生物的P素表现为净矿化,从而使土壤中的P含量增加。本研究土壤的C∶P<200,表明土壤P含量相对充足,但相较于其他区域(表7),该区较高的C∶P也表明黔西南石漠化区土壤P矿化能力弱,有效性偏低, TP不易转化为有效磷,这与已有研究得出的岩溶地区P有效性偏低的结果一致[11]。该区人工植物群落长期施入P肥,使P富余,影响土壤元素平衡和植物根系对养分的吸收,今后应重视计量施肥。
土壤N∶P通常用于诊断植物生长受N或P限制,指示土壤养分供应水平[29]。本研究中土壤N∶P高于其他地区(表7),这可能是因为湿热地区P淋溶速度较N素流失更强烈[21],因此导致N∶P较高。有研究[5,11]认为,在陆地土壤生态系统中,生物固N量与土壤N∶P呈反比关系,该区土壤较高的N∶P也预示着其生物固N量较其他生态系统低,即被植物吸收有效N能力较低,反映了该区植物生长易受N素限制。Koerselman等[31-32]提出:当植物体N∶P<14时受N限制,N∶P>16时受P限制,而N∶P在14~16时受N和P共同限制。本研究中,4种人工植物群落N∶P均远低于14,指示N亏缺,表明群落植物生长易受N限制,这与长期施入P肥形成P素的过量累积有关。今后应对人工恢复植物群落实施精准配方施肥,避免养分元素浪费,且有利于调控土壤环境。
4 结 论
SOC、TN随土层的加深为呈高及低的垂直分布格局,表聚效应明显。TP在各土层间无明显变化,但不同群落土壤之间的TP含量变异系数达35.71%。植被类型是影响土壤养分分异的主要因子,人工恢复植物群落土壤养分均显著高于荒地,表明适宜的人工恢复群落有利于喀斯特石漠化区土壤养分的恢复。
土壤C、N、P生态化学计量比在0~40 cm土壤剖面上的总体格局为上高下低。C∶N、C∶P、N∶P的变异系数表明C∶P、N∶P属中等变异,C∶N属弱变异。4种人工恢复植物群落N∶P远低于14,指示N亏缺,植物生长易受N限制,今后应对人工恢复植物群落实施计量施肥,避免养分元素浪费。
土壤SOC、TN、TP呈两两极显著正相关,表明全量养分之间关系密切。C∶P与C∶N、N∶P呈极显著正相关,表明土壤生态系统内养分平衡具有较强的正向调控机制;C∶P与TN呈显著负相关,验证了C、N之间存在互馈效应;SOC与N∶P之间为反向作用效应,表明C制约着N、P元素的比例关系。
猜你喜欢
当代水产(2022年8期)2022-09-20
建材发展导向(2022年10期)2022-07-28
——以贵州威宁县为例
测绘地理信息(2022年2期)2022-04-02
昆明医科大学学报(2022年2期)2022-03-29
绿色科技(2022年4期)2022-03-21
现代园艺(2021年19期)2021-12-08
防护林科技(2020年3期)2020-05-13
学校教育研究(2020年7期)2020-04-09
当代陕西(2020年24期)2020-02-01
当代陕西(2020年24期)2020-02-01
