
目标树经营对兴安落叶松人工林土壤胞外酶的影响
2021-12-28 08:23于江波王玉娇杨立学张庆江
中南林业科技大学学报 2021年12期
于江波,及 利,刘 月,王玉娇,张 岩,杨立学,张庆江,陈 龙,赵 鑫
(1.东北林业大学 a.林学院;b.森林生态系统可持续经营教育部重点实验室,黑龙江 哈尔滨 150040;2.尚志国有林场管理局小九林场,黑龙江 尚志 150600)
人工林在全球森林资源中扮演着重要角色,自20世纪60年代以来,人工林逐步替代天然林,成为木材的主要供应源,我国已经成为世界上发展人工林最多的国家[1-2]。长期以来,人工林经营过分重视和追求短期生产力与经济利益,偏重于人工更新和同龄纯林培育模式,忽视了森林的自然演替规律、天然更新、森林健康等多目标近自然经营模式,导致纯林经营引起的地力衰退、土壤酸化、环境恶化以及林分生产力下降等问题越来越普遍[3]。目标树经营是以单株林木为对象而进行的一种近自然森林经营作业体系,通过不断的抚育间伐,保持林分的最佳混交状态和目标树最大生长量,保证林分天然更新,促使林分质量不断提高[4-9]。有研究发现目标树经营一方面重新分配林内水、热、光等环境因子,另一方面改变凋落物归还土壤中的基质数量、质量和分解速率,进而提高林地物种多样性和土壤肥力[10]。土壤胞外酶(Extracellular enzyme)广泛存在于土壤中,是由植物根系及土壤微生物分泌到土壤中发挥催化作用的蛋白质。胞外酶在土壤中具有不可替代的关键作用,参与土壤中几乎所有的生物化学反应,将微生物及植物无法吸收的大分子有机物降解为可吸收的小分子物质[11]。
由于研究区的土壤类型、植被类型、区域气候条件及土壤养分状况差异,目标树经营对于土壤养分、胞外酶活性的影响一直没有一致结论[12-15],而且目前国内外目标树经营相关的研究区域多集中在水热条件较好的暖温带、亚热带地区,对寒温带地区树种的研究相对较少[16-18]。兴安落叶松Larix gmelinii是我国大兴安岭地区森林的优势种和建群种,对保持大兴安岭地区生态系统的稳定、保护物种多样性和森林可持续发展都起到非常重要的作用[19]。因此,本研究以目标树经营6 a 后的兴安落叶松人工林为对象,通过分析目标树经营后的土壤养分含量与土壤酶活性的变化以及二者之间的关系,探讨目标树经营在提高人工林土壤肥力中的作用,为加速形成优质高产的人工林提供理论基础和实践支撑。
1 材料与方法
1.1 研究区概况
本研究试验地位于大兴安岭松岭林业局绿水林场119 林班。该地区位于大兴安岭林区南端,伊勒呼里山东南坡,嫩江上游左岸,地理坐标为123°29′13″~125°11′10″E,50°09′16″~51°23′48″N,海拔高度介于400~700 m 之间,平均海拔为556 m。年平均温度为-3℃,极端最高温度为38℃,极端最低温度为-48℃。年平均降水量600 mm,主要集中在7月和8月,占全年降水量的48.9%。年均蒸发量1 153 mm,年相对湿度63.5%。年无霜期为100~110 d,植物生长期为90~100 d。林业局作业区内主要土壤为棕色针叶林土和沼泽土等[20]。该地区针叶树种主要有兴安落叶松、樟子松Pinus sylvestris;阔叶树种有白桦Betula platyphylla、蒙古栎Quecus mongolica、黑桦Betula davurica、山杨Populus davidiana、毛赤杨Alnus sibirica等。主要灌木有小叶杜鹃Rhododendron parvifolium、大黄柳Salix raddeana、沼柳S.rosmarinifolia、杜香Ledum palustrevar.angustum N.Busch、越橘Vaccinium vitisidaea、红瑞木Swida alba、山荆子Malus baccata、稠李Padus racemosa、蓝靛果忍冬Lonicera caerulea、刺玫蔷薇Rosa davurica等。
1.2 试验设计与取样方法
试验地设置区域为1988年营造的兴安落叶松人工林,造林密度为2 400 株/hm2。2013年进行首次疏伐,采伐强度在31%,保存株数为1 700株/ hm2。2014年生长季前按照目标树经营措施,在经营样地内选取生长力旺盛、状态良好、干形饱满通直且没有机械损伤和病虫害的林木作为目标树,定牌永久性标记,将目标树胸径的25 倍范围内影响目标树生长的干扰树选定并伐除。在林隙和林窗内保护天然更新白桦、蒙古栎等乡土树种。在林龄达到45 a 时进行再一次抚育,确定最终目标树,将最终目标树的树冠完全解放,形成自由树冠。终伐前15 a 进行最后一次疏伐,释放空间培养第二代目标树,促进天然更新,如天然更新不够,再进行人为干预。达到目标胸径后,根据地形和林木成熟情况采取带状或群团状采伐。
2019年8月通过初步踏查,分别在经营林分和未经营林分设置3 块30 m×30 m 的样地,样地间距约20 m,并对样地进行每木检尺,样地基本概况见表1。在进行土壤取样时,将表层植物残体清除,在每个样方按0~10、10~20、20~40 cm 3 层分别进行梅花状5 点取样,每个样方的同一个土层分别充分混合,分为2 份样品。一部分土样置于4℃冰箱内,用于测定土壤酶活性及铵态氮和硝态氮含量,另一份土样自然风干后用于土壤其他养分指标测定。

表1 兴安落叶松目标树经营样地基本概况Table 1 Basic situation of the management plot of crop tree of L.gmelinii
1.3 土壤养分的测定
土壤pH 值采用酸度计测定。土壤有机碳、全氮采用碳氮分析仪(Vario MACRO,Elementar Co,Germany)测定,土壤全磷与有效磷采用双酸浸提钼锑抗比色法测定[21]。土壤铵态氮和硝态氮用连续流动分析仪(Tecator FIA Star 5000 Analyzer,Foss Tecator,Sweden)测定含量[22]。土壤微生物量碳氮采用氯仿熏蒸法测定[23]。
1.4 土壤胞外酶活性的测定
土壤胞外酶活性采用微孔板荧光法多功能酶标仪(SynergyH4,BioTek)测定[24],反应底物及底物反应浓度见表2,具体方法为:取1 g 鲜土,用125 mL pH 值为5.0 的50 mmol·L-1醋酸钠缓冲液充分混匀,制备土壤悬浮液,用移液枪吸取200 μL 土壤悬浮液和50 μL 底物(空白为醋酸钠)注入96微孔板内,水解酶在20℃恒温黑暗条件下培养4 h,加入10 μL 1 mol·L-1氢氧化钠终止反应,反应1 min 后使用酶标仪测定荧光值,荧光值在365 nm 波长处激发,450 nm 波长处测定,每个样品测8 个平行。氧化酶活性采用吸光光度计来测定(氧化酶在450 nm 处测定吸光值),将微孔板置于黑暗中20℃孵育18 h。过氧化物酶在多酚氧化酶的基础上,每个孔再添加10 µL 的0.3%双氧水。最后通过土壤干质量和反应时间来计算胞外酶的活力,所测定的土壤胞外酶活性以nmol·g-1h-1为单位来表示。
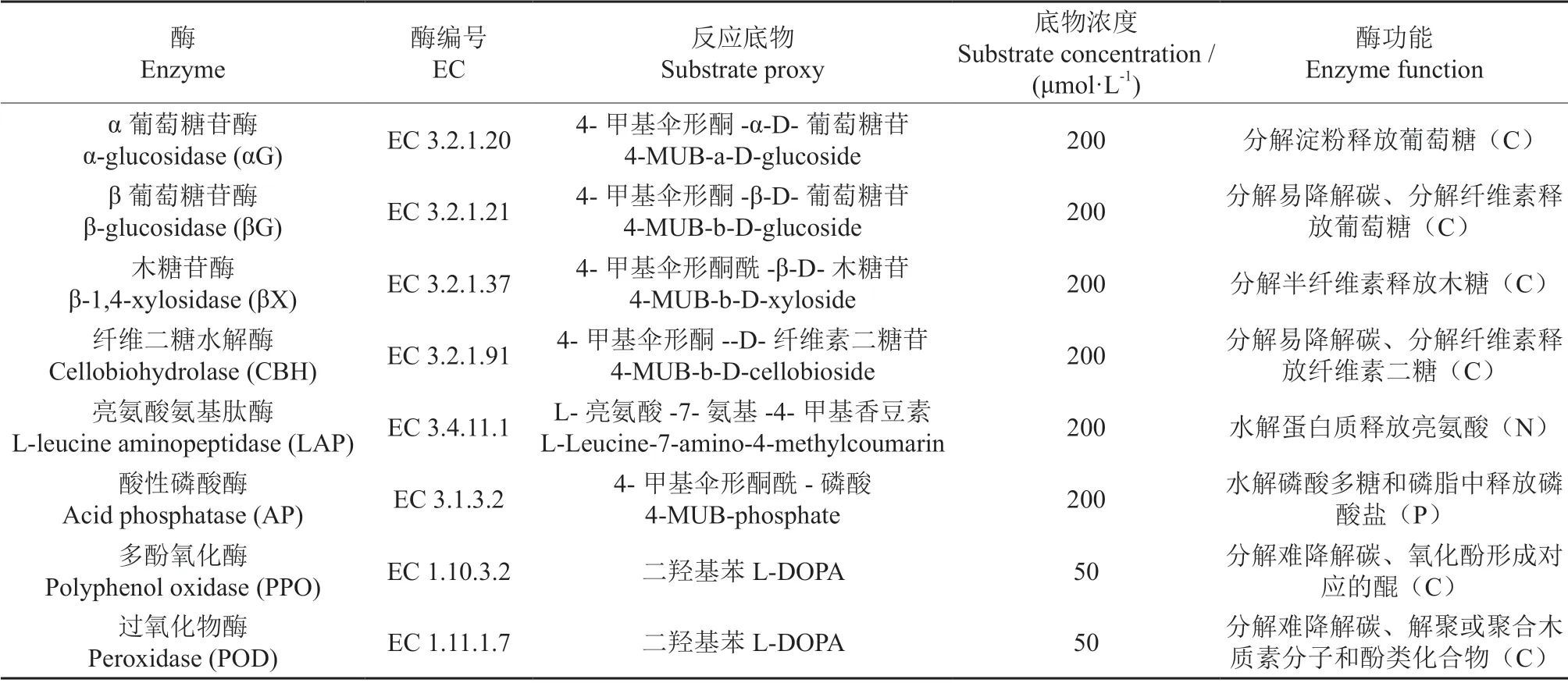
表2 土壤胞外酶的功能及反应底物Table 2 Functions and substrates of the soil extracellular enzymes
1.5 统计分析
利用Excel 2016 和SPSS 22.0 软件进行数据处理。通过单因素(One-way ANOVA)方差分析和最小显著差异法(LSD)分析目标树经营对土壤养分和酶活性的影响,以及不同土层下土壤养分含量和酶活性的差异(α=0.05)。采用Canoco 5.0软件,以土壤酶活性为响应变量,同时以土壤养分指标为解释变量做冗余分析(RDA)。利用SigmaPlot 12.5 软件作图,图表中数据为平均值±标准差。
2 结果与分析
2.1 目标树经营对不同土层土壤养分的影响
与对照相比,经营组中土壤有机碳的含量在0~10 cm 土层中显著增加了18.5%(P<0.05),经营组中的碳氮比在0~10 cm 和10~20 cm 两个土层中分别显著增加了40.5%和21.8%(P<0.05),经营组中的有效磷含量在0~10 cm 土层中显著增加了30.1%(P<0.05),微生物量碳含量在0~10 cm 和10~20 cm 两个土层中分别显著增加了64.2%和37.8%(P<0.05),微生物量氮含量在0~10 cm 土层中显著增加了32.3%(P<0.05)。而经营组中的全氮含量在0~10 cm 土层中显著减少了19.8%(P<0.05)。不同土层的土壤养分差别较大,目标树经营对土壤养分的影响在表层(0~10 cm)最明显,随土层加深,土壤养分含量均降低(表3)。
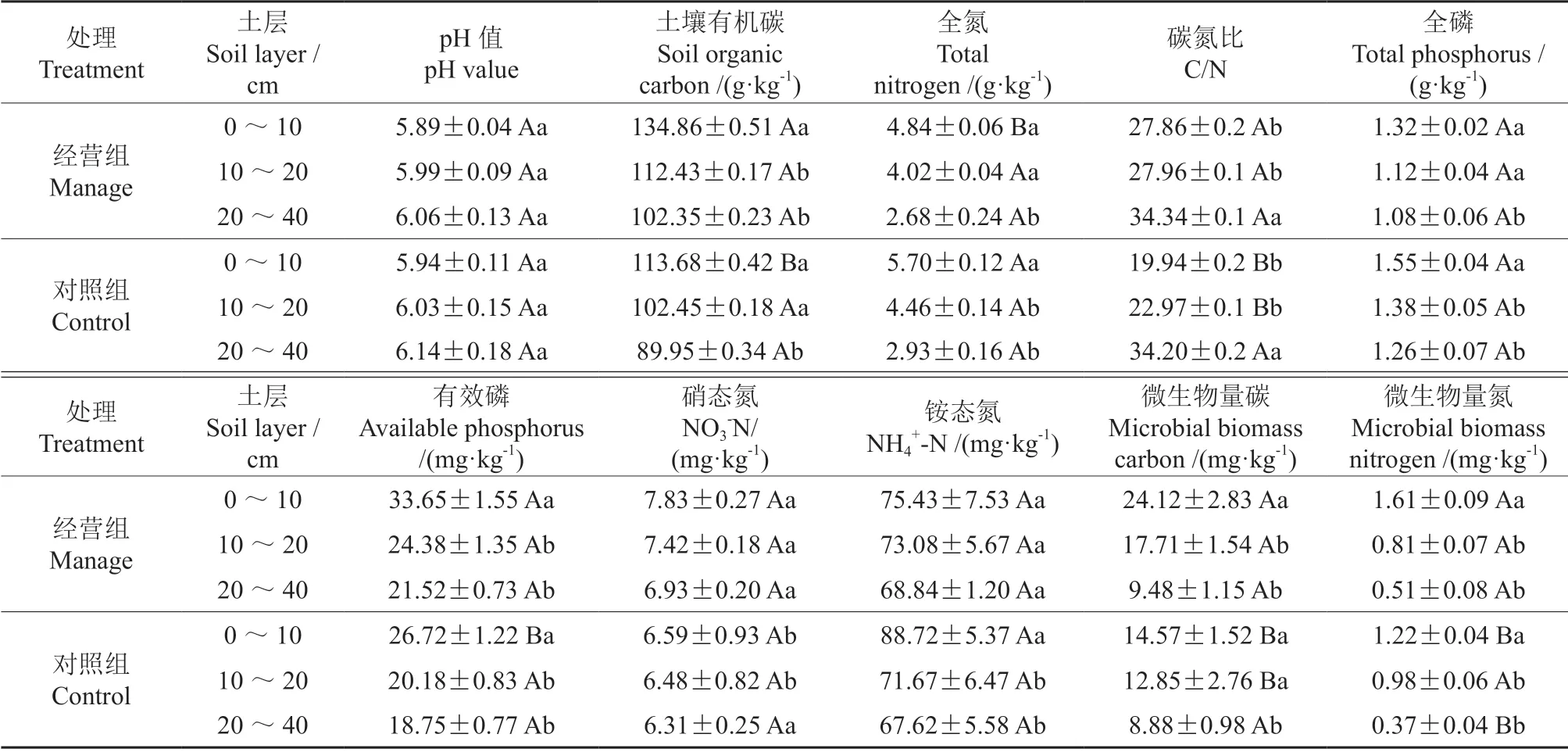
表3 兴安落叶松目标树经营后土壤养分的变化†Table 3 Changes in soil nutrients of L.gmelinii crop trees after management
2.2 目标树经营对不同土层土壤胞外酶活性的影响
与对照组相比,经营组中的多酚氧化酶活性在0~10 cm 土层中显著提高了16.8%(P<0.05),β 葡萄糖苷酶活性在0~10 cm 和10~20 cm 两个土层中分别显著降低了45.8%和45.1%(P<0.05),木糖苷酶活性在0~10、10~20 和20~40 cm 3 个土层中分别显著降低了146.8%、220.4%和246.1%(P<0.05)(图1)。
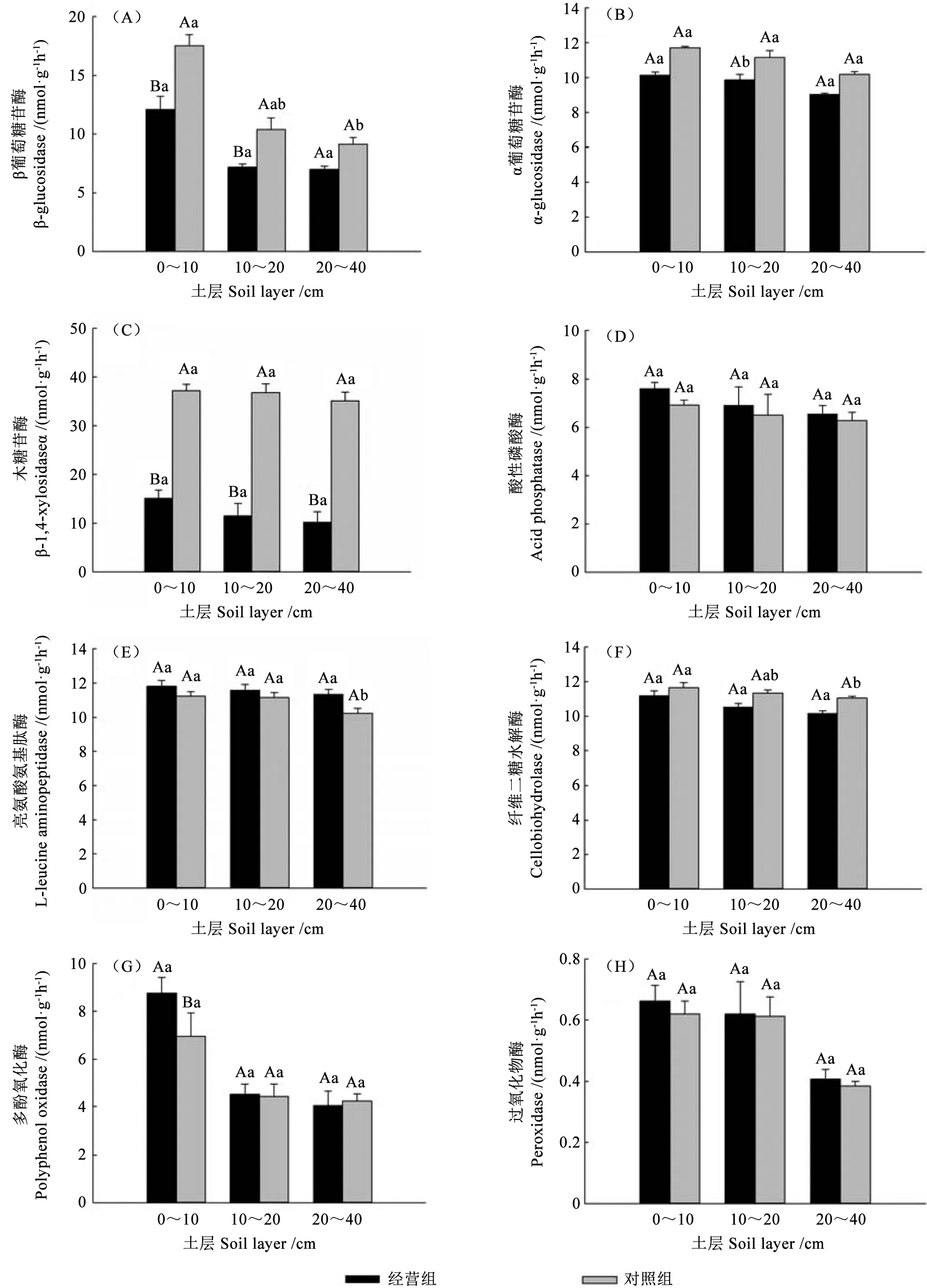
图1 兴安落叶松目标树经营后土壤胞外酶活性的变化Fig.1 Changes of soil extracellular enzyme activity after management of L.gmelinii crop tree
2.3 不同土层土壤酶活性变化的冗余分析
在0~10 cm 土层,第1 轴和第2 轴共解释了土壤酶活性变异程度的84%,第1 轴主要由木糖苷酶和亮氨酸氨基肽酶解释,影响这2 种酶的主要因素是土壤有机碳(P<0.01),且与第1 轴呈明显正相关,解释了该土层土壤酶变异的83.5%,说明土壤有机碳在0~10 cm 土层中是影响土壤酶活性的主要因素,土壤有机碳与木糖苷酶和亮氨酸氨基肽酶均呈正相关。在10~20 cm土层中,第1 轴和第2 轴共解释了土壤酶活性变异程度的91%,第1 轴主要由酸性磷酸酶和β 葡萄糖苷酶解释,影响这2 种酶的主要因素是土壤有机碳(P<0.05),解释了该土层土壤酶变异的95.9%,且与第1 轴呈明显正相关,土壤有机碳与酸性磷酸酶均呈正相关,与β葡萄糖苷酶呈负相关。在20~40 cm 土层,第1 轴和第2 轴共解释了土壤酶活性变异程度的99%,第1 轴主要由酸性磷酸酶和亮氨酸氨基肽酶解释,影响土壤酶的主要因素是全磷(P<0.05),全磷解释了该土层土壤酶变异的37.7%,且与第1 轴呈明显正相关,全磷与亮氨酸氨基肽酶均呈正相关,与酸性磷酸酶呈负相关(图2,表4)。
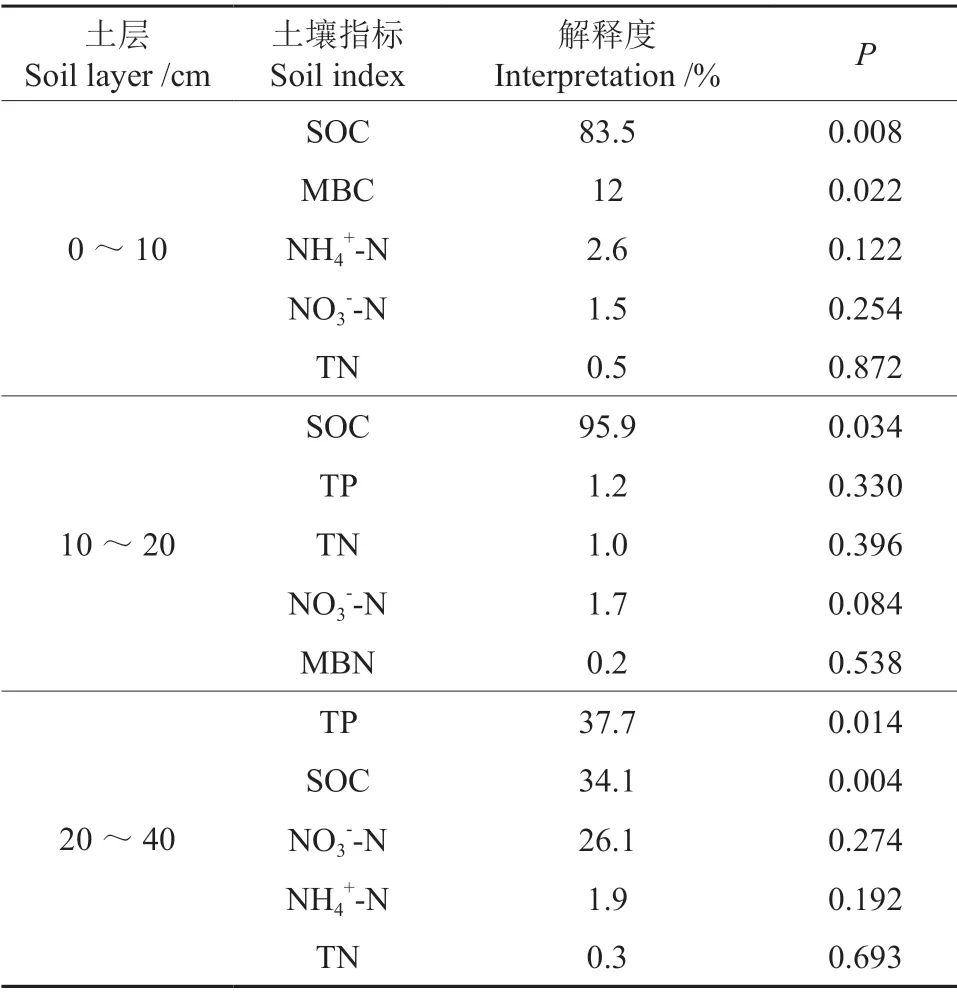
表4 对土壤胞外酶变化有显著影响作用的环境因子相应解释比例Table 4 Corresponding explanatory ratios of environmental factors that have a significant effect on the changes of soil extracellular enzymes
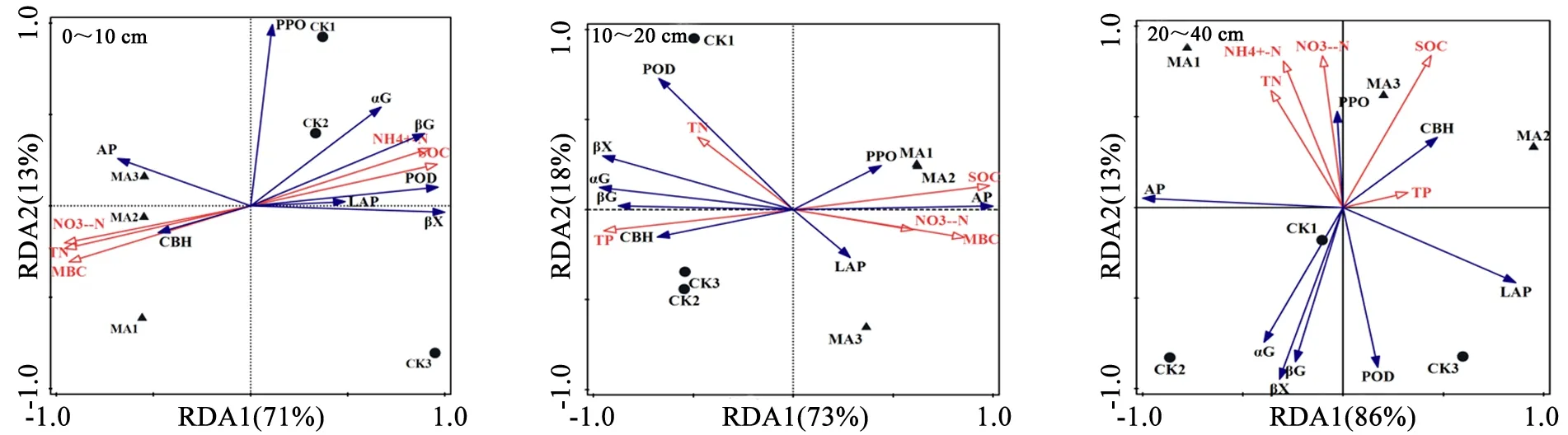
图2 兴安落叶松目标树经营下土壤养分对土壤胞外酶活性影响的冗余分析Fig.2 Redundant analysis of the influence of soil nutrients on soil extracellular enzyme activity under the management of L.gmelinii crop tree
3 讨 论
目标树经营作为近自然林业经营的重要措施之一,在改变林分结构的同时,林下植被、光照、温度以及水分等环境因子的变化,从而引起土壤表层的凋落物分解、土壤有机质形成、土壤生物化学过程和养分循环等变化[25]。本研究结果表明,目标树经营后土壤有机碳、有效磷和微生物量碳氮的含量显著增加,土壤全氮含量显著降低。土壤有机碳作为土壤肥力的物质基础,其含量的增加主要是源于兴安落叶松人工林内前期实验样地修建时产生更多的植物残体以及目标树经营后各样地内的林下植被多样性增加[26]。赵朝辉等[27]在研究间伐对杉木林内土壤理化性质的影响时发现,间伐改变了土壤温度、湿度等条件,加速了死地被物的分解,从而改善了土壤理化性质,提高了土壤有机质的含量,这与本研究的结果一致。土壤中全氮含量显著降低,而有效磷养分显著增多,这与李旭等[14]对云冷杉人工林间伐的研究结果一致。主要原因是在生长季期间,目标树经营后样地内有着更加丰富的植被群落,因此土壤中氮磷的含量必须以更快的速度矿化成植物可利用的形式,随后由更高的根生物量吸收利用,使得土壤全氮和全磷含量减少,速效养分增多,以满足植被在生长季对养分的需求[10]。微生物量碳氮虽然在土壤养分含量中占比极少,却是土壤有机质中最活跃的组分之一,也是表征土壤微生物数量的常用指标,微生物数量又是评价土壤肥力的重要指标之一[28]。微生物量碳氮含量的增加主要是因为目标树经营后,一方面林下植被多样性提高,土壤养分循环加速,使得微生物群落多样性和数量显著提高;另一方面改变了林下微环境,加速根系分泌,促进凋落物分解,影响土壤中的有机质的数量与质量,最终影响微生物群落[29]。Mendham 等[30]研究表明间伐5 a 后的桉树林中土壤微生物量碳氮显著降低,这与本研究的结果不一致,可能是由于树种、地理位置或间伐时间等不同造成的。
土壤胞外酶活性对生物与非生物环境的变化十分敏感,是土壤系统的物质循环与能量转换的积极参与者,也常被视为是判定土壤质量的重要指标[31]。目标树经营后土壤微环境以及植物多样性的改变有利于土壤微生物活动,从而增强土壤胞外酶活性。本研究发现,不同的土壤胞外酶对目标树经营的响应不同,并非所有胞外酶都受单一影响而以相同的方式产生变化,目标树经营后β葡萄糖苷酶与木糖苷酶活性显著降低,多酚氧化酶活性显著提高。参与土壤易分解有机碳反应的β葡萄糖苷酶与木糖苷酶活性在目标树经营后,均呈显著的降低趋势,这与 Garcia 等[32]有关间伐的研究结果是一致的。有研究表明,土壤中的易分解有机碳主要来自于地下根系和地上凋落物,在目标树经营伐除干扰树后,导致地下根系生物量和地上凋落物减少,使得林分内可利用的易分解有机碳含量减少,所以进一步影响了β 葡萄糖苷酶与木糖苷酶的活性[33]。多酚氧化酶是土壤中主要的木质素降解酶,目标树经营后分解难降解碳的多酚氧化酶活性在表层土壤中呈显著的增加趋势。于立忠等[34]研究发现,间伐显著提高了日本落叶松人工林的多酚氧化酶活性,这与本研究的结果一致。主要原因是经营后林内土壤表层残留了很多木屑和枯枝残渣,增加了土壤中木质素、粗纤维和多酚类顽固性碳的输入,目标树经营后土壤中难分解的碳相对较多,所以使得多酚氧化酶的活性有所提高[26]。
冗余分析的结果表明,土壤有机碳是驱动0~20 cm 土层土壤酶活性变化的主要因子,说明土壤有机质是影响表层土壤酶活性的重要因素,另外酶活性的增强也有利于加剧土壤有机质的矿化作用,土壤中微生物可利用的碳氮养分含量增高,从而加速土壤养分循环和碳周转[35]。凋落物、根系、动植物残体等碳源主要积累于表层土中,所以土壤有机碳在土壤表层对酶活性变化的主导作用更显著[36]。全磷是驱动20~40 cm 土层土壤酶活性变化的主要因子。由此可见,土壤有机碳并不是影响土壤胞外酶活性的唯一因素,全磷也是影响土壤酶活性的重要因子,全磷与土壤有机碳共同影响着土壤中的酶活性。磷作为温带森林生态系统的限制性养分元素之一,随着土壤深度的增加,磷含量会显著下降,且相比于土壤碳氮养分含量较低,导致深层土壤中缺磷现象加剧,此时解磷微生物会通过分泌相关的胞外酶将有机养分转化为供植物根系吸收利用的无机养分。全磷作为土壤中磷的总贮量指标,具有关键作用,为土壤中有机磷向无机磷的转化提供底物,因此全磷可能是决定20~40 cm 土层酶活性的关键因子[37]。在高纬度寒区森林生态系统中,除土壤环境因子外,植被群落多样性、植物根系生物量以及土壤微生物群落等也是驱动土壤胞外酶活性变化的重要因素。因此,下一步我们还需结合这些指标进行研究。
4 结 论
目标树经营对兴安落叶松人工林土壤养分及胞外酶活性都有显著的影响。冗余分析表明,目标树经营引起的土壤养分的变化是影响土壤胞外酶活性变化的主要原因。本研究由于实验时间较短,因此还需长期观测目标树经营对兴安落叶松人工林的影响,以期更好地揭示近自然经营背景下土壤胞外酶与土壤养分之间的作用机制,能够为人工林的可持续经营提供更加全面、科学的参考依据。
猜你喜欢
中草药(2022年20期)2022-11-15
食品科学(2022年20期)2022-10-31
现代食品科技(2022年9期)2022-10-09
农业工程学报(2022年8期)2022-08-08
建材发展导向(2022年10期)2022-07-28
北京大学学报(自然科学版)(2022年3期)2022-06-17
绿色科技(2022年4期)2022-03-21
当代陕西(2020年24期)2020-02-01
当代陕西(2020年24期)2020-02-01
当代陕西(2020年24期)2020-02-01
