
基于线粒体D-loop的河南省绵羊品种遗传多态性和遗传关系研究
2021-12-31 06:29韩浩园姚丽娟王惠绘尹慧茹王先宁吴依依李玉法赵金艳贾万里王拥庆
西南农业学报 2021年11期
韩浩园,姚丽娟,王惠绘,尹慧茹,王先宁,吴依依,李玉法,赵金艳,贾万里,李 君,王拥庆,权 凯*
(1.河南牧业经济学院动物科技学院,河南 郑州 450046;2.渑池县农业农村局,河南 三门峡 472400;3.沈丘县农牧科技研发中心,河南 周口 466300)
【研究意义】线粒体DNA(mtDNA)遵循母系起源,作为研究动物起源进化的有效手段,近期已被广泛应用于研究遗传多样性、亲缘关系和遗传分化[1-5]。目前基于线粒体序列对绵羊遗传关系和起源进化研究较为广泛,但中国河南省绵羊品种小尾寒羊、豫西脂尾羊和河南大尾羊之间的遗传关系研究很少,特别是河南大尾羊和豫西脂尾羊群体数量急剧减少,有必要对其遗传资源进行评估,并分析河南省3个绵羊品种间的遗传关系,对于今后3个品种的保护和利用提供遗传学依据。【前人研究进展】绵羊线粒体DNA序列为16.616 kb[6]。早期国外学者对线粒体控制区(D-loop)序列的研究表明绵羊有2个母系起源,其中欧洲的摩佛伦羊(Ovisorientalis)为家养绵羊的母系起源之一,同时排除了羱羊(Caprasibirica)和盘羊(Argali sheep)作为家养绵羊祖先的可能[7],而随后发现家养绵羊可能至少存在3个母系起源[8-10]。中国新疆、内蒙古、西藏、云南、宁夏、山东、甘肃等地区18个地方绵羊品种存在3个独立的母系起源,且绵羊的3个支系可能曾经经历过群体扩张[11-15]。对中国地方绵羊品种mtDNA控制区(D-loop)序列遗传多样性的研究发现:新疆、内蒙古、西藏、云南、宁夏、山东等地区7个地方绵羊品种D-loop区遗传变异丰富[13-15]。中国甘肃绵羊群体mtDNA D-loop遗传多样性均较贫乏,应该对其遗传资源进行保护[14]。【本研究切入点】河南省绵羊品种包括小尾寒羊、豫西脂尾羊和河南大尾羊。小尾寒羊和豫西脂尾羊都属于我国蒙古羊系统[16],是我国绵羊良种。豫西脂尾羊是河南古老的地方绵羊品种,分布在豫西、豫南地区,具有产肉性能好、适应性广、抗病力强、耐粗放饲养等优良特性。小尾寒羊为国家级保护品种,以体型大、生长快、繁殖率高而闻名。河南大尾羊目前濒临绝种,本研究仅发现极少数个体。【拟解决的关键问题】目前对河南省绵羊品种间遗传关系和母系起源尚无系统研究,因此本研究以河南省3个绵羊品种作为研究对象,通过分析mtDNA D-loop区全序列研究其遗传多样性、遗传分化和系统进化关系,揭示品种间的遗传分化及其起源进化,继而了解它们的遗传资源状况,为今后开展河南省绵羊品种的保存、种质特性研究和开发利用等工作提供理论依据。
1 材料与方法
1.1 样本采集
本研究采集23只小尾寒羊(XW)、40只豫西脂尾羊(YX)和4只河南大尾羊(DW)共67个个体的外周血样本。小尾寒羊采自河南雨轩农业科技发展有限公司,豫西脂尾羊采自河南省三门峡市渑池县豫西脂尾羊保种群,依据品种特性和羊场记录采集纯种个体。河南大尾羊采自河南省汝州市汝阳县山区养殖户。利用EDTA抗凝管颈静脉采血5 mL,采集血样置于-20 ℃保存。
1.2 DNA提取和D-loop序列扩增及测序
采用小量全血基因组DNA快速提取试剂盒(艾德莱生物)提取3个绵羊品种全血DNA,并利用Nanodrop(Thermo Fisher)测量提取DNA浓度和OD260/280值。将合格DNA稀释到20 ng/μL。由生工生物工程(上海)股份有限公司合成绵羊D-loop全序列上游引物F:5′- TCCCTAAGACTCAAGGAAGAAGC-3′和下游引物R:5′- AAGAGTGTGTGCTTGATACCTGC -3′序列[17]。PCR扩增反应体系为25 μL,包括DNA模板(20 ng/μL)1 μL,正、反向引物(10 μmol/L)各0.5 μL,Taq PCR Master Mix 12.5 μL(1 U Taq Polymerase、5 μmol/L dNTP、0.2 mmol/L Tris-HCl、1 mmol/L KCl、0.03 mmol/L MgCl2)和ddH2O 10.5 μL。PCR扩增条件为:95 ℃预变性4 min;94 ℃变性30 s,58 ℃退火60 s,72 ℃延伸60 s,36个循环;72 ℃延伸9 min,4 ℃保存。利用1.5%琼脂糖凝胶电泳对PCR产物进行检测,确认产物大小与目的片段一致,送往生工生物工程(上海)股份有限公司双向测序。
1.3 数据分析
从NCBI上下载五条绵羊序列作为对照:摩佛伦羊(Ovismusimon),登录号为AY091487;羱羊(Ovisammon),登录号为AJ238300;盘羊(Ovisvignei),登录号为AY091490;亚洲型A型和欧洲型B型,登录号为AF039578和AF039577。
利用Lasergene软件包SeqMan软件拼接双向序列。利用MEGA6.0软件比对67只绵羊的D-loop序列(Align by ClustalW)并截齐序列[18],基于Kimura 2-parameter模型构建ML系统进化树,计算3个品种间的遗传距离[19]。利用DnaSP V5软件[20]分析3个绵羊品种D-loop序列的DNA多态性(单倍型多样度Hd、核苷酸多样度Pi等)、基因流和遗传分化(种群间核苷酸平均差异数Kxy、核苷酸歧义度Dxy、固定系数Fst等遗传学参数)、Tajima’s检验和单倍型鉴定。基于单倍型数据,利用NETWORK 10.2.0.0软件(Fluxus Technology Ltd.,Kiel,Germany)构建67只小尾寒羊、豫西脂尾羊和河南大尾羊D-loop单倍型网络图。
2 结果与分析
2.1 河南3个绵羊品种的D-loop序列变异及遗传多样性分析
本研究发现61只绵羊D-loop序列为1376 bp,而2个小尾寒羊(XW15和XW21)存在75 bp插入,长度为1451 bp;4个豫西脂尾羊(YX1、YX14、YX15和YX31)存在75 bp缺失,长度为1301 bp。小尾寒羊多态位点数为132个,核苷酸多样度为0.0230,共定义23个单倍型,单倍型多样度为0.975。豫西脂尾羊多态位点数为129,核苷酸多样度为0.0129,共定义25个单倍型,单倍型多样度为0.936。河南大尾羊多态位点数为55个,核苷酸多样度为0.0270,共定义3个单倍型,单倍型多样度为0.667(表1)。小尾寒羊和豫西脂尾羊单倍型多样度较高,河南大尾羊单倍型多样度最低,可能与河南大尾羊数量稀少有关。3个品种无共享单倍型。
2.2 系统进化树与遗传距离分析
基于3个绵羊品种D-loop序列构建系统发育树,ML树结果表明,小尾寒羊、豫西脂尾羊和河南大尾羊聚为三大支(lineage A~C),lineage A大部分为豫西脂尾羊,为亚洲A型;lineage B大部分为小尾寒羊,为欧洲B型,与摩弗伦羊聚为1支;lineage C大部分为小尾寒羊,为中国绵羊特有分支(图1)。羱羊和盘羊为外群。河南大尾羊穿插于小尾寒羊和豫西脂尾羊中,说明河南大尾羊血统不纯。遗传距离分析表明3个品种间遗传距离均大于0.02(表2),达到亚种间遗传距离,其中,小尾寒羊和豫西脂尾羊间遗传距离最远,河南大尾羊与小尾寒羊遗传距离最近。
2.3 遗传分化和中性检验
应用DnaSP 5.0软件计算反映小尾寒羊和豫西脂尾羊种群间基因差异程度的遗传学参数。结果显示,3个品种间固定系数(Fst)范围为0.0253~0.4465,遗传分化系数(Gst)范围为0.0226~0.0706(表3)。3个品种间核苷酸差异数(Kxy)范围为30.239~42.934,核苷酸歧义度(Dxy)范围为0.0241~0.0342(表3)。河南大尾羊和豫西脂尾羊间Kxy和Dxy最高,小尾寒羊和河南大尾羊间最低,说明3个品种间小尾寒羊和豫西脂尾羊、豫西脂尾羊和河南大尾羊间极度分化,核苷酸差异大;小尾寒羊和河南大尾羊间分化很弱,核苷酸差异最小,与遗传距离结果一致。采用Tajima’s D方法进行中性检验,小尾寒羊和豫西脂尾羊Tajima’s D中性检测为不显著(P> 0.10)。河南大尾羊显著偏离中性(P< 0.05),且Tajima’s D 值为正(表1),说明河南大尾羊近年来经历过瓶颈效应。
2.4 构建单倍型网络图
基于线粒体D-loop序列,对小尾寒羊和豫西脂尾羊进行单倍型网络图构建,共发现50个单倍型和3个单倍群(A、B、C),对应系统发育树lineage A~C,Network图结果表明河南省绵羊存在3个母系起源(图2)。
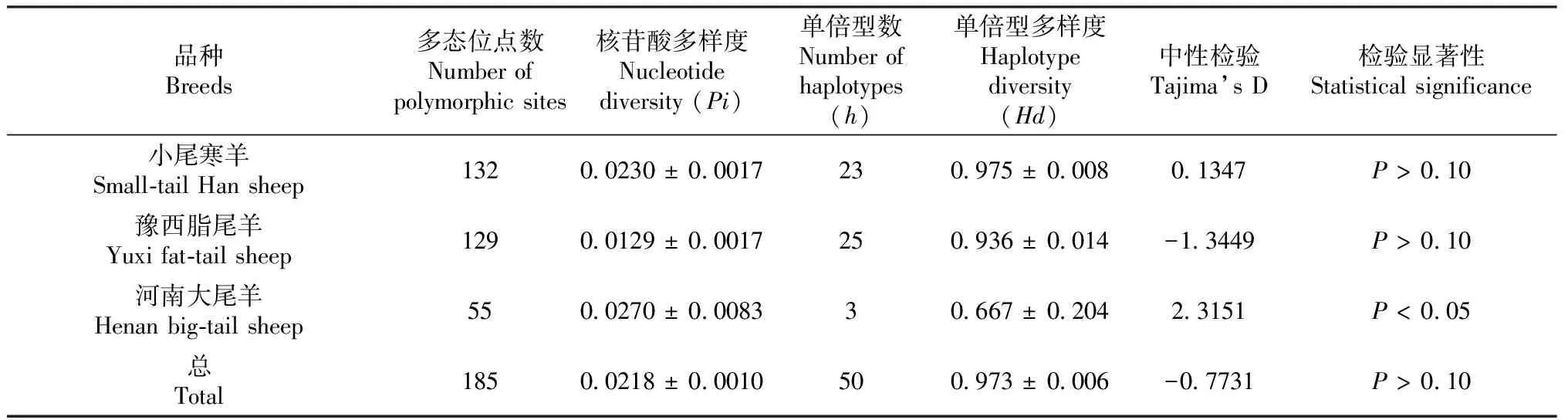
表1 河南绵羊品种遗传多样度

表2 3个河南省绵羊品种间遗传距离
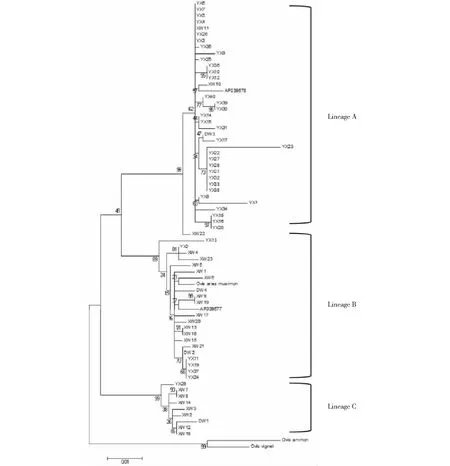
图1 河南省绵羊品种ML系统进化树Fig.1 The ML phylogenetic tree of Henan sheep breeds
3 讨 论
3.1 河南省绵羊品种遗传多样性
本研究以3个河南省绵羊品种小尾寒羊、豫西脂尾羊和河南大尾羊为研究对象,研究了河南省3个绵羊品种遗传多态性和遗传关系。遗传多态性结果显示绵羊线粒体D-loop区的变异除少量的插入/缺失外,6个个体出现了75 bp串联重复序列的重复次数不同,与之前的研究一致[21]。小尾寒羊群体变异位点132个,占9.59%;豫西脂尾羊群体变异位点129个,占9.38%。且2个绵羊品种单倍型较多,分别存在23和25个单倍型,单倍型多样性丰富,核苷酸多样性丰富,属于高单倍型多样性和高核苷酸多样性的遗传多样性模式。这与牛华锋等[22]的研究结果一致。小尾寒羊和豫西脂尾羊的遗传变异必定受到养殖和选育的影响,在群体连续繁殖过程中,群体内积累了较多的遗传变异,说明选育过程中的基础群体种质较优,遗传多样性较高,遗传改良育种保留了具有优良性状的个体,发挥种质优势[23],证明小尾寒羊和豫西脂尾羊具有较高的进化潜力和环境适应力,是优质的河南省绵羊品种,且没有出现种质衰退的现象[24-25]。河南大尾羊多态位点数为55个,较小尾寒羊与豫西脂尾羊少,河南大尾羊单倍型多样度及定义单倍型数目(n=2)为3个品种中最低的,可能与河南大尾羊样品数量少有关。目前河南大尾羊和豫西脂尾羊个体数很少,因此开展河南大尾羊及豫西脂尾羊的遗传多样性研究及品种种质资源保护是非常必要的。
3.2 河南省绵羊品种间的遗传关系
本研究基于ML法构建小尾寒羊、豫西脂尾羊和河南大尾羊D-loop序列的系统进化树,可见河南大尾羊穿插在小尾寒羊与豫西脂尾羊之间。本次采样仅采到4只河南大尾羊样本,证明该品种数量少,且血统掺杂了小尾寒羊和豫西脂尾羊血统,纯种河南大尾羊几乎绝种。
遗传距离遗传距离分析表明3个品种间达到亚种间遗传距离,其中,小尾寒羊和豫西脂尾羊间遗传距离最远,河南大尾羊与小尾寒羊遗传距离最近,与遗传分化系数、核苷酸歧异度、核苷酸差异度及系统发育树结果一致。该结果揭示了小尾寒羊、豫西脂尾羊和河南大尾羊间的遗传距离远近。
系统发育树及Network结果表明豫西脂尾羊和小尾寒羊有少数个体互相掺杂,说明3个品种间存在一定的基因交流。20世纪90年代,河南省多地利用小尾寒羊体型大、生长速度快、繁殖率高的品种优势,对当地的豫西脂尾羊进行改良,以提高豫西脂尾羊综合生产性能[26-27],因此造成小尾寒羊和豫西脂尾羊间的基因交流,与本研究的基因流结果一致。
3.3 种群进化与种群扩张历史
关于绵羊的野生祖先,目前还没有一个统一的看法。本研究所测定的河南省家养绵羊3个品种分为3大支系:支系A占所有个体的55.23%,支系B占31.34%,支系C占13.43%,并且亚洲A型和欧洲B型分别与支系A和B聚为一类,有研究表明中国绵羊至少存在三个母系起源[8,14]。且认为第三个分支C数量不多[28],本研究验证了中国绵羊存在有3个独立的母系起源。聚类图显示,摩佛伦羊与其中分支B聚为一类,而羱羊和盘羊单独聚为一类这说明摩佛伦羊对中国家养绵羊的贡献较大,而没有发现羱羊和盘羊对中国绵羊的起源有贡献,这与国内外学者的研究结果一致[6]。
本研究中性检验结果显示,小尾寒羊和豫西脂尾羊的Tajima’s D检验结果不显著,表明该序列在进化上遵循中性模型,这2个绵羊品种在较近的历史上未经历群体扩张及瓶颈效应[29],保持相对稳定的群体规模。但河南大尾羊Tajima’s D检验结果显著,说明河南大尾羊近期经历瓶颈效应,与近期河南大尾羊数量急剧减少现状一致,因此有必要加速对河南大尾羊遗传资源的研究及保护。
4 结 论
本研究基于线粒体D-loop序列,对小尾寒羊、豫西脂尾羊和河南大尾羊遗传多样性及起源进化进行了研究,得出以下结论:河南省小尾寒羊、豫西脂尾羊品种遗传多样性较丰富;小尾寒羊与河南大尾羊遗传关系最近,与豫西脂尾羊关系最远;河南省绵羊品种有3个母系起源,且摩佛伦羊对河南省绵羊起源与进化有更大的贡献。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
今日农业(2022年2期)2022-06-01
情感读本·道德篇(2022年2期)2022-04-12
戏曲研究(2021年2期)2021-03-09
儿童时代·幸福宝宝(2019年11期)2019-12-13
吉林农业·下半月(2017年12期)2017-12-18
现代农业科技(2016年22期)2017-03-24
数学大王·低年级(2016年8期)2016-05-14
