
非受体酪氨酸激酶Src在异丙肾上腺素介导脂肪分解中的作用
2022-01-07 04:01程扣辉姜允奇王文景李文奇李子健马鑫冯磊
中国心血管杂志 2021年6期
程扣辉 姜允奇 王文景 李文奇 李子健 马鑫 冯磊
214122 江南大学药学院和无锡医学院(程扣辉、李文奇);100191 北京大学第三医院心内科、血管医学研究所,卫生部心血管分子生物学与调节肽重点实验室,分子心血管学教育部重点实验室,心血管受体研究北京市重点实验室(姜允奇、王文景、李子健);214122 江南大学无锡医学院(马鑫、冯磊)
在能量不足或能量消耗增加时,哺乳动物脂肪组织激活交感神经系统,导致白色脂肪组织中的三酰甘油在一系列脂肪酶如三酰甘油脂肪酶(adipose triglyceride lipase,ATGL)、激素敏感脂肪酶(hormone-sensitive lipase,HSL)和单酰基甘油脂肪酶等的作用下,分解为游离脂肪酸和甘油,从而满足生理需要[1-2]。随着研究的深入,发现脂肪细胞的脂解涉及蛋白质复合物在脂滴表面的组装和拆解。β肾上腺素能受体(β-adrenergic receptor,β-AR)是G蛋白偶联受体家族成员,介导多种肾上腺素和去甲肾上腺素的生理反应[3],肾上腺素与β-AR结合,激活蛋白激酶A(protein kinase A,PKA)通路,导致ATGL磷酸化,并触发脂肪分解[4-6]。除PKA经典通路外,研究还发现儿茶酚胺可通过激活β-AR/Gi/Src通路,促进脂肪分解[7-9]。肉瘤病毒蛋白(sarcoma,Src)是一种非受体酪氨酸激酶,尽管有研究表明在细胞水平抑制Src激酶活性可减轻异丙肾上腺素(isoproterenol,ISO)诱导的脂肪分解,但在整体动物层面上关于Src在AR激活诱导的脂肪分解中的作用还未得到证实。因此,本实验旨在整体动物水平探讨Src在ISO诱导脂肪分解中的作用和机制。
1 材料与方法
1.1 主要试剂
Dulbecco’s modified Eagle’s medium(DMEM)(Gibco公司);胎牛血清(fetal bovine serum,FBS)(北京全式金生物技术有限公司);Bicinchoninic acid(BCA)蛋白定量试剂盒(Thermo scientific公司);DMSO(Sigma公司);ISO(Sigma公司);Src抑制剂吡唑并嘧啶类化合物(4-amino-5-(4-methylphenyl)-7-(t-butyl)pyrazolo[3,4-d]-pyrimidine,PP1)(Sigma公司);p-Src抗体(1∶1 000,Cell signaling公司);t-Src抗体(1∶1 000,Cell signaling公司);β-actin抗体(1∶10 000,Cell signaling公司);HRP标记山羊抗兔IgG二抗(中杉金桥公司);HRP标记山羊抗小鼠IgG二抗(中杉金桥公司);Trizol试剂(Invitrogen公司);qPCR Super mix(北京全式金生物技术有限公司);RT-PCR引物合成(北京生工生物技术公司)。
1.2 实验动物
SPF级雄性10周龄的C57BL/6野生型小鼠购自北京大学医学部实验动物中心及北京维通利华公司,饲养环境由北京大学医学部实验动物中心提供(室温23℃、相对湿度65%、12 h明暗交替)。所有动物均经过北京大学健康科学中心机构护理和使用委员会批准。根据实验需求将小鼠分为对照组、ISO组和ISO+PP1组。ISO组每天背部皮下注射10 mg/kg体重的ISO,连续2周;ISO+PP1组,在此2周内,腹腔注射1.5 mg/kg体重的PP1,每周3次。
1.3 细胞处理
小鼠前体脂肪细胞系3T3-L1使用DMEM培养基(含10% FBS、100 Unit/ml Penicillin和100 μg/ml Streptomycin)在37℃、5% CO2环境中培养。ISO组:1 μM ISO处理;ISO+PP1组:PP1预孵育1 h后用ISO处理;对照组:加入相同体积DMSO。
1.4 蛋白免疫印迹实验
将处理好的3T3-L1细胞加入预冷的细胞裂解液,冰上裂解细胞15 min后,用细胞刮收集匀浆液,BCA蛋白定量法测定蛋白浓度,加入1/4体积的5×loading buffer,100℃煮沸5 min使蛋白质变性。使用10% SDS-PAGE凝胶分离细胞裂解物。将蛋白从凝胶中转移到硝酸纤维膜中,之后用5%的脱脂奶粉室温封闭1 h。将NC膜条带置于特制的抗体孵育盒,加入一抗,置于4℃冷室中240 r/min的摇床上过夜孵育。次日回收一抗,TBST洗膜(10 min,3次),然后用加入辣根过氧化物酶标记的IgG二抗室温孵育1 h,弃去二抗,TBST洗膜(10 min,3次)后即可显影。
1.5 RNA分离和实时反转录聚合酶链反应(RT-qPCR)
使用Trizol试剂(Ambion,Life Technologies,MA,USA)从小鼠附睾脂肪组织和3T3-L1细胞中提取总RNA。紫外分光光度计测定RNA浓度,用反转录试剂盒将1 μg总RNA逆转录合成cDNA,再用SYBR qPCR Master Mix试剂盒在20 μl反应体系下进行实时荧光定量PCR反应检测相关基因mRNA表达。引物序列见表1。以GAPDH作为内参,每个样三个复孔,采用Δ循环阈值(Ct)法处理结果,所有基因表达结果表示为倍数的变化,使用2-ΔΔCt方法计算mRNA的相对表达量。PCR扩增程序如下:其中预变性为95℃,30 s;循环反应为95℃,10 s;60℃,30 s,共40个循环;融解曲线反应为95℃,15 s;60℃,60 s;95℃,15 s。每个循环结束后监测SYBR GreenⅠ荧光发射。
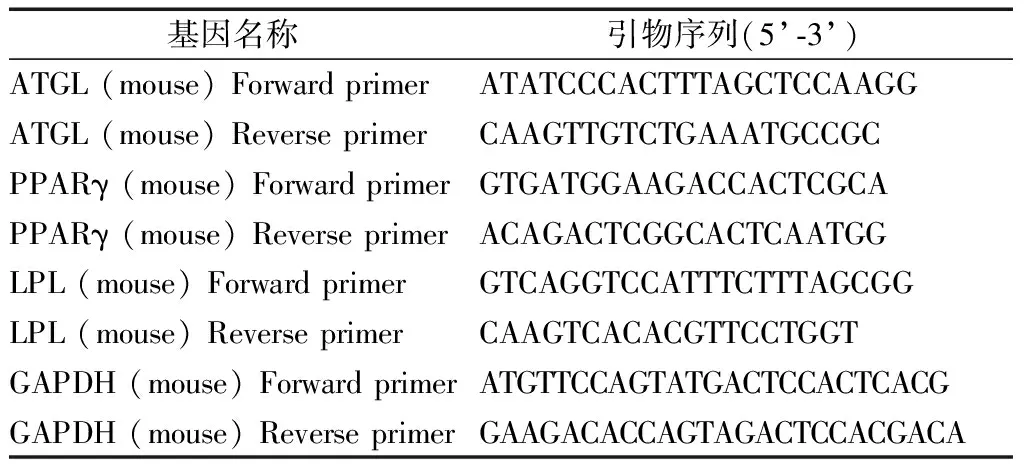
表1 实时荧光定量PCR引物信息
1.6 统计学方法
2 结果
2.1 ISO诱导小鼠附睾脂肪分解
在小鼠背部注射ISO 2周后,检测小鼠附睾脂肪重量,发现与对照组相比,ISO组小鼠的附睾脂肪显著减少(P<0.001),但两组小鼠体重无明显差异(图1A~D);苏木精和伊红(HE)染色显示,与对照组相比,ISO组的脂肪细胞大小显著降低(P<0.001)(图1E)。qPCR分析表明,与对照组相比,ISO组的附睾脂肪中脂解相关基因ATGL的mRNA水平显著升高(P<0.05)(图1F);脂肪合成基因过氧化物酶体增殖物激活受体γ(peroxisome proliferators-activated receptorsγ,PPARγ)和脂蛋白脂肪酶(lipoprtein lipase,LPL)的mRNA水平均显著降低(均为P<0.01)(图1G)。实验结果表明,ISO在小鼠体内可促进脂肪分解。
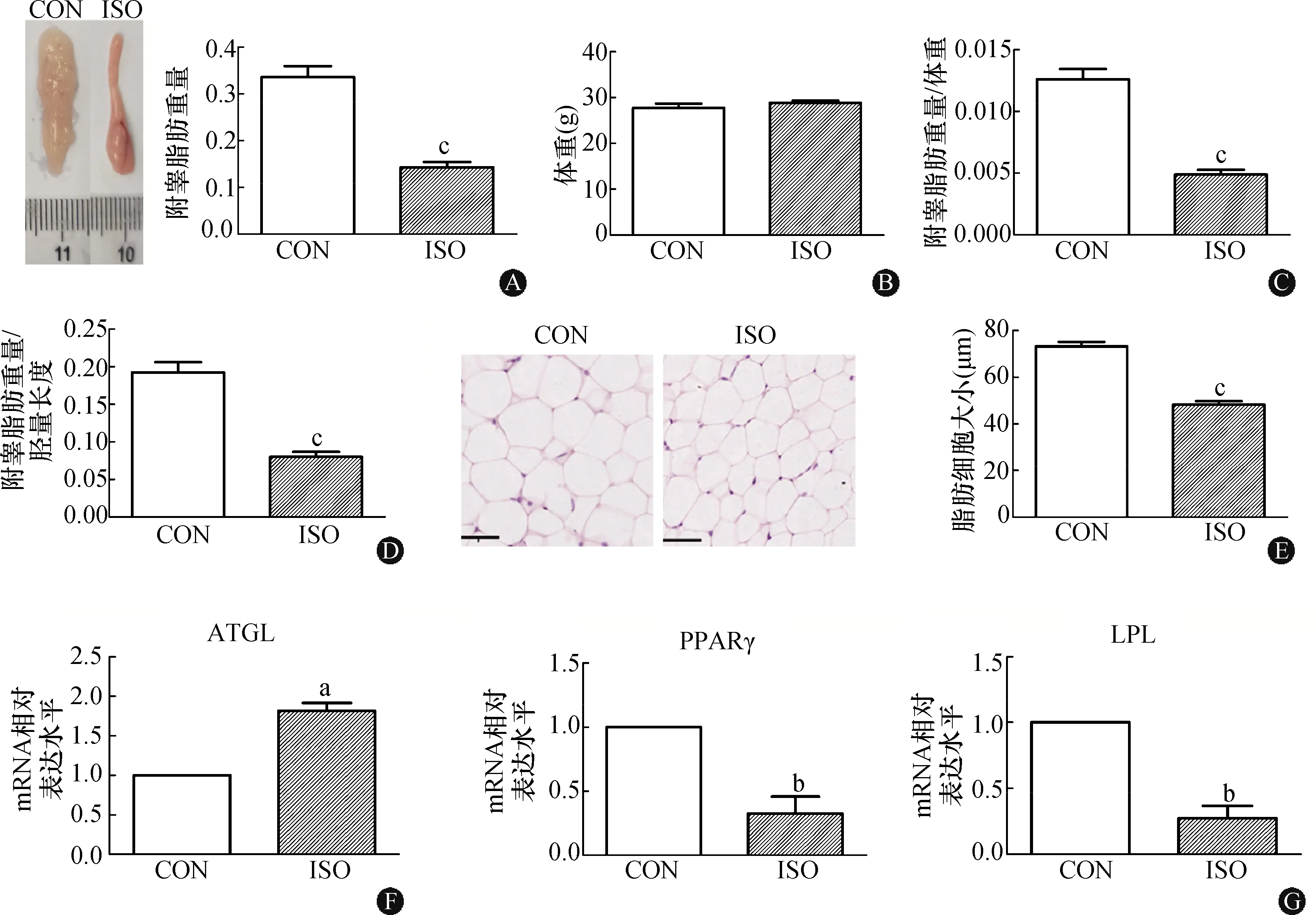
CON:对照组;ISO:异丙肾上腺组;A:小鼠附睾脂肪大体图(比例尺刻度单位:mm)及附睾脂肪重量;B:小鼠体重;C:附睾脂肪重量与体重的比值;D:附睾脂肪重量与胫骨长度的比值;E:小鼠附睾脂肪组织HE染色(标尺:50 μm)及细胞大小统计;F:脂肪分解基因ATGL mRNA的表达水平;G:脂肪合成基因PPARγ和LPL mRNA的表达水平。与对照组相比,aP<0.05,bP<0.01,cP<0.001(n=4)
2.2 ISO刺激促进Src活性
给予脂肪细胞ISO刺激后,蛋白免疫印迹实验显示Src磷酸化显著增加(P<0.05),见图2,提示ISO刺激诱导Src活性增加。

CON:对照组;ISO:异丙肾上腺组;A:蛋白免疫印迹;B:Src磷酸化数据统计分析。与对照组相比,aP<0.05(n=5)
2.3 PP1抑制ISO诱导的Src激活
PP1是Src酪氨酸激酶的高效抑制剂[10]。在给予脂肪细胞PP1预处理后,蛋白免疫印迹分析表明,ISO刺激诱导的Src磷酸化被显著抑制(P<0.01),见图3。
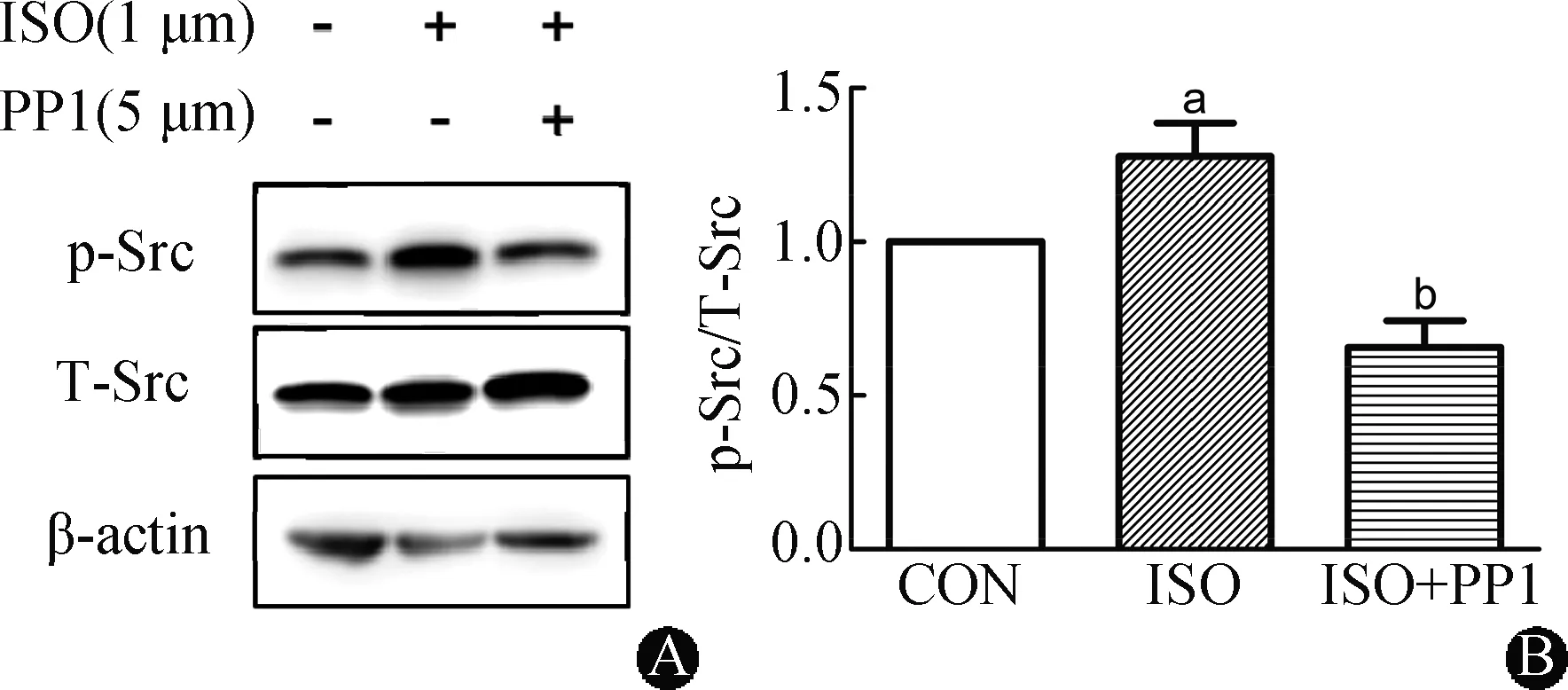
CON:对照组;ISO:异丙肾上腺组;ISO+PPI:异丙肾上腺+吡唑并嘧啶类化合物组;A:蛋白免疫印迹;B:Src磷酸化数据统计分析。与对照组相比,aP<0.05;与ISO组相比,bP<0.01(n=5)
2.4 ISO诱导小鼠附睾脂肪分解依赖于Src
在小鼠注射ISO的同时注射Src激酶抑制剂PP1后,检测脂肪分解相关指标显示,与ISO组相比,ISO+PP1组小鼠附睾脂肪重量增加(P<0.05),但三组小鼠体重无明显差异(图4A~B);此外,与ISO组相比,ISO+PP1组小鼠的附睾脂肪重量与体重比值(P<0.01)以及与胫骨长度比值(P<0.05)均显著增加(图4C~D)。HE染色显示,与ISO组相比,ISO+PP1组脂肪细胞变大(P<0.001)(图4E)。qPCR数据分析表明,与ISO组相比,ISO+PP1组小鼠附睾脂肪脂解基因ATGLmRNA水平显著降低(P<0.01)(图4F);脂肪合成基因PPARγ(P<0.01)和LPL的mRNA水平升高(图4G)。提示ISO在小鼠体内促进脂肪分解的作用在一定程度上依赖Src激酶活性。
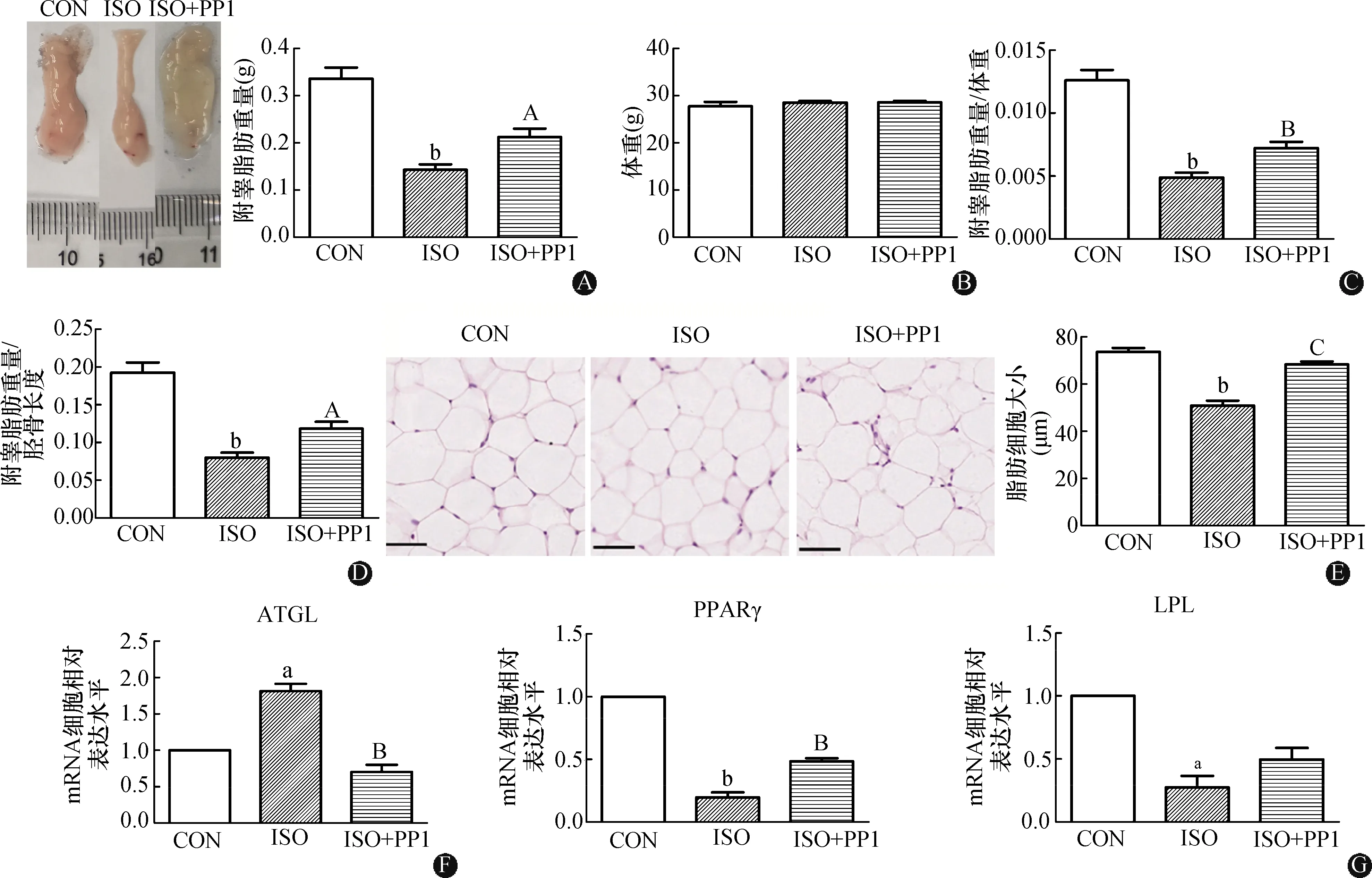
CON:对照组;ISO:异丙肾上腺组;ISO+PPI:异丙肾上腺+吡唑并嘧啶类化合物组;A:小鼠附睾脂肪大体图(比例尺刻度单位:mm)及附睾脂肪重量;B:小鼠体重;C:附睾脂肪重量与体重的比值;D:附睾脂肪重量与胫骨长度的比值;E:小鼠附睾脂肪组织HE染色(标尺:50 μm)及细胞大小统计;F:脂肪分解基因ATGL mRNA的表达水平;G:脂肪合成基因PPARγ和LPL mRNA的表达水平。与对照组相比,aP<0.01,bP<0.001;与ISO组相比,AP<0.05,BP<0.01,CP<0.001(n=4)
3 讨论
在正常情况下,哺乳动物脂肪组织能够微调一系列神经内分泌信号,精确地适应三酰甘油合成(脂肪生成)和分解(脂肪分解)之间的平衡,以满足生理需要[11-12]。脂肪细胞脂质分解过程中三酰甘油贮存和动员的控制因子是脂质积累的重要调控者,一旦某些调控因子功能失效,脂解作用发生紊乱,则能够导致机体脂肪酸利用失常,诱发肥胖、胰岛素抵抗和糖尿病等疾病[13-14]。脂肪细胞分解产生的游离脂肪酸是外周组织能量需求时重要燃料;此外,释放的游离脂肪酸也可调节葡萄糖和胰岛素的活性和产生,参与胰岛素抵抗的发生发展[15-16]。因此,脂肪细胞脂质分解不仅调节能量平衡和脂肪分布,其对于代谢稳态也是至关重要的。
脂肪分解过程ATGL水解三酰甘油成二酰基甘油,开始脂解过程;HSL催化二酰基甘油生成单酰甘油;单酰基甘油脂肪酶最终会水解单酰甘油转化成脂肪酸和甘油。2004年,ATGL首次被描述为三酰甘油水解酶,其特异性水解三酰甘油,对其他脂质底物的活性非常有限。研究表明,ATGL在脂肪组织中高度表达;同时,它也在心脏、骨骼肌、肝脏和其他组织中表达[17-18]。ATGL活性受磷酸化修饰的调控,研究发现AMP活化蛋白激酶使ATGL Ser406位点磷酸化,增加了ATGL的活性,从而增加脂肪分解[19]。其次,HSL是最早发现的脂解酶,其活性对儿茶酚胺、促肾上腺皮质激素和胰高血糖素等激素敏感[20]。与ATGL类似,HSL在脂肪组织中高表达,在肌肉、睾丸、类固醇组织和胰岛等其他组织中均可检测到其表达[21]。儿茶酚胺是脂解的主要正向调节因子,通过与β-AR结合,引起AR激活,AR属于G蛋白偶联蛋白受体家族。AR激活引起偶联的G蛋白亚基被释放,根据G蛋白的亚型激活(Gs亚基)或抑制(Gi亚基)腺苷酸环化酶活性[7]。激活的腺苷酸环化酶可增加细胞质中环磷酸腺苷(cyclic adenosine monophosphate,cAMP)水平,随后导致蛋白激酶A(protein kinase A,PKA,也称为cAMP依赖蛋白激酶)的激活,从而使HSL磷酸化并强烈激活脂解[3-6]。β-AR存在3种亚型,β1-AR、β2-AR、β3-AR,它们在脂肪细胞中共同表达,其中β3-AR在脂肪细胞中表达最丰富,其与异源三聚体G蛋白Gs结合,促进细胞内cAMP的升高和PKA的激活[22]。然而,也有证据表明,这些受体也可与Gi结合导致Src招募,进而促进Src/Erk依赖性的脂肪分解,促进脂肪的进一步分解[23]。
在本研究中,我们通过在小鼠体内皮下连续注射ISO两周后发现,小鼠附睾脂肪重量显著降低,脂肪细胞较对照组显著变小,脂肪分解和合成基因的mRNA水平与脂肪重量变化一致。此外文献研究表明,Src在3T3-L1细胞脂肪分解中起一定作用[7,24],我们在细胞水平上的结果与文献结果一致,证实ISO能增加Src的磷酸化,PP1预处理后Src活性显著降低。同样,在小鼠体内腹腔注射Src激酶抑制剂PP1后发现,小鼠附睾脂肪重量较ISO组显著升高,脂肪细胞显著变大,脂肪分解和合成基因的mRNA水平与脂肪重量变化一致。上述实验结果证实,Src激酶活性在AR激活诱导的脂肪分解中起一定作用。因此,我们研究证实了在动物体内β-AR激动剂ISO诱导脂肪分解依赖Src激酶活性。
本研究存在不足之处:Src在体内通过影响脂解基因如ATGL和HSL去促进脂肪分解的进一步机制还亟待研究。总之,本研究发现Src在ISO诱导的脂肪分解中存在一定作用,未来可通过干预Src激酶活性从而改善肥胖等相关代谢性疾病。
利益冲突:无
猜你喜欢
云南农业大学学报(自然科学)(2022年4期)2022-07-28
畜牧兽医学报(2022年6期)2022-07-07
中国畜牧杂志(2022年4期)2022-04-15
现代泌尿外科杂志(2021年5期)2021-05-27
中华养生保健(2020年9期)2021-01-18
科学24小时(2018年1期)2018-01-10
中学科技(2017年11期)2017-12-26
现代养生·下半月(2016年6期)2016-10-21
科技视界(2015年4期)2015-01-02
科学大众(中学)(2009年8期)2009-09-06
