
基于SCoT分子标记的黄河鲤抗嗜水气单胞菌不同群体遗传多样性分析
2022-02-03 07:57张岚姜淑泓陈玉函常乐管俊翔林强于淼乔志刚王磊
南方农业学报 2022年10期
张岚,姜淑泓,陈玉函,常乐,管俊翔,林强,于淼,乔志刚,王磊*
(1河南师范大学水产学院/河南省水产动物养殖工程技术研究中心/水产动物疾病控制河南省工程实验室,河南新乡 453007;2中国水产科学研究院珠江水产研究所,广东广州 510380)
0 引言
【研究意义】黄河鲤(Cyprinusy carpio)隶属于鲤形目(Cypriniformes)鲤科(Cyprinidae),主产于黄河流域,是我国重要的淡水经济鱼类,也是我国四大淡水名鱼之一;因其适应性广,肉质鲜嫩,营养价值高且寓意美好而受到广大消费者喜爱,目前已发展成为我国主要的淡水养殖品种之一。近年来,由嗜水气单胞菌(Aeromonas hydrophila)等引起的败血病频发导致黄河鲤大量死亡(Jiang et al.,2022),给黄河鲤养殖业带来巨大经济损失。抗生素等药物的广泛使用极易造成细菌耐药性增加及环境污染等问题(杨先乐和郑宗林,2007)。因此,加快抗病品种培育对保障黄河鲤养殖业健康绿色发展具有重要意义。【前人研究进展】遗传多样性是一个物种群体生存适应和发展进化的前提。在遗传相似性较高的宿主间,病原体传播的可能性更高,而遗传多样性高的群体抵抗疾病暴发的能力更强(Altizer et al.,2003;King and Lively,2012;Gibson,2021;Pérez-González et al.,2021)。在罗氏沼虾(Macrobrachium rosenbergii)(朱其建等,2013;董丁健和戴习林,2020)、尼罗罗非鱼(Oreochromis niloticus)(张栋,2018)、凡纳滨对虾(Litopenaeus vannamei)(方振朋,2019)、黄颡鱼(Pelteobagrus fulvidraco)(张佳佳,2019)中的研究均表明物种遗传多样性对其抗病性能、生长性状、营养价值等方面具有积极作用。也有研究表明,近交过程中遗传多样性降低会从根本上影响种群抵御各种病原体的能力(Amills et al.,2004;Wang et al.,2020)。因此,抗病品种的选育通常伴随着某些基因的杂合及群体遗传多样性的提高(董丁健和戴习林,2020)。目标起始密码子多态性(SCoT)作为一种新型的分子标记(Collard and Mackill,2009),具有操作简单、重复性好、可在物种中通用等优点(Gorji et al.,2011;Korir et al.,2012),特别是作为一种目的基因分子标记,不仅能获得与性状联系紧密的目的基因,还能对性状进行跟踪,非常有利于分子标记辅助育种(Poczai et al.,2013)。目前,SCoT分子标记广泛应用于农作物品种鉴定(Korir et al.,2012)、DNA指纹图谱构 建(Cabo et al.,2014;Hamidi et al.,2014)及遗传多样性分析(Etminan et al.,2016;欧景莉等,2019)等研究领域。【本研究切入点】SCoT分子标记在水生生物中的研究主要集中在群体遗传变异(Marie and Allam,2017;Abu Almaaty et al.,2020)及性别特异性分子标记筛选(Mohamed et al.,2019;Mahgoub et al.,2021)等方面,但针对鱼类抗病育种的研究鲜见报道。【拟解决的关键问题】利用SCoT分子标记对黄河鲤抗嗜水气单胞菌不同群体的遗传多样性及遗传结构进行分析,探讨遗传多样性与其抗嗜水气单胞菌能力的关系,为黄河鲤的抗病育种提供理论依据。
1 材料与方法
1.1 试验材料
供试黄河鲤(平均体质量130 g)购自山东省东阿县绣青水产养殖专业合作社,暂养于室内有效养殖水体1.7 m3的帆布池中,水温控制在(28±2)℃,持续24 h增氧,确认健康、无不良症状后用于后续试验。嗜水气单胞菌由中国水产科学研究院珠江水产研究所提供,4℃保存备用。
1.2 感染处理方法
将嗜水气单胞菌菌液加入100 mL脑心浸液肉汤(BHI)培养基中,经28℃摇床(180 r/min)过夜培养12~18 h后取1 mL菌液进行PCR验证,防止PCR扩增过程中受污染;菌液8000 r/min离心10 min,收集菌体,以无菌PBS进行重悬并稀释至1.23×107CFU/mL。随机挑选300尾黄河鲤放入试验池中,水温保持在(28±2)℃,暂养2 d,禁食。然后按照2 µL/g的剂量对每尾黄河鲤进行腹腔注射嗜水气单胞悬液,腹腔注射后每2 h观察1次,如遇鱼体死亡及时捞出并记录死亡时间,剪取鳍条保存于无水乙醇中。连续7 d无死亡后停止试验并统计数据。
1.3 DNA提取与检测
根据死亡顺序分别选取最先死亡(The first dead population,FP)、最后死亡(The last dead population,LP)及存活(Survival population,SP)的黄河鲤各30尾,按照编号选取相应鳍条,取相同大小(约0.5 cm×0.5 cm)的鳍条剪碎,然后采用动物组织/细胞基因组DNA提取试剂盒(D2100-100T,北京索莱宝科技有限公司),按照试剂盒说明提取基因组DNA。以1.0%琼脂糖凝胶电泳进行检测,通过NanoDrop One/OneC微量核酸蛋白浓度测定仪测定DNA浓度,质量合格的DNA置于-20℃冰箱保存备用。
1.4 SCoT引物筛选及PCR扩增
随机选取4份DNA样品进行SCoT引物初筛选,从80条SCoT引物中筛选多态性好、重复性高、条带清晰的引物(表1)。SCoT-PCR反应体系20.0 μL:2×TaqPCR Master Mix 10.0 μL,SCoT引物1.0 μL,DNA模板20 ng,ddH2O补足至20.0 μL。PCR扩增程序:95℃预变性4 min;95℃40 s,50℃50 s,72℃90 s,进行36个循环;72℃延伸10 min,PCR扩增产物4℃保存。取8.0 µL SCoT-PCR扩增产物经1.8%琼脂糖凝胶电泳后,以BIO-RAD凝胶成像系统检测并拍照保存。

表1 80条SCoT引物的序列信息Table 1 80 SCoT primers and information of their sequences
1.5 统计分析
SCoT-PCR扩增产物按同一迁移水平下有无条带分别赋值,有条带记为1,无条带记为0,人工读取和统计条带。使用PopGene32对3个黄河鲤群体的所有个体及群体等位基因数(Na)、有效等位基因数(Ne)、Nei's基因多样性指数(H)和Shannon's信息指数(I)进行分析,采用Arlequin 3.5对黄河鲤3个群体进行分子方差变异(AMOVA)分析,通过MEGA 7.0中的非加权组平均法(UPGMA)进行聚类分析,并以Structure 2.3进行遗传结构分析。
2 结果与分析
2.1 不同抗性群体的划分结果
人工感染嗜水气单胞菌后24 h即出现黄河鲤死亡现象,随后黄河鲤死亡率增加,至感染后48 h达峰值,300尾黄河鲤的累积死亡率为40.0%。在人工感染试验过程中最先死亡的视为高感群体(FP),最后死亡的视为低感群体(LP),而人工感染嗜水气单胞菌后1周仍存活的视为抗病群体(SP)。感染后死亡的黄河鲤腹部泛红甚至溃烂,泄殖孔充血,鳃丝出现腐烂现象;解剖发现其腹腔及内脏有出血点,符合黄河鲤感染嗜水气单胞菌的病理学特征。
2.2 SCoT位点多态性及群体遗传多样性分析结果
选取部分DNA模板进行PCR扩增,结果从80条SCoT引物中筛选出18条扩增条带清晰、多态性好的引物(表2)。图1为引物SCoT34在3个黄河鲤群体中的PCR扩增电泳结果。筛选出的18条SCoT引物共扩增出103条条带,范围为3~11条,其中多态性条带97条,平均每条SCoT引物扩增出5.4条多态性片段,范围为2~11条,多态性比率为94.17%。在3个黄河鲤群体中,SP群体的多态性比率最高,为83.50%;其余2个群体的多态性比率分别为79.61%(FP)和81.55%(LP)。3个黄河鲤群体的Na范围为1.7961~1.8155,Ne范围为1.3828~1.4029,H范围为0.2258~0.2467,I范围为0.3453~0.3804。在FP、LP和SP群体中,Na、Ne、H和I均依次递增(表3)。群体间的遗传分化指数(Gst)为0.0972,即90.28%的遗传多样性分布在群体内部,表明黄河鲤群体间的遗传变异小于群体内的遗传变异。

表3 3个黄河鲤群体的遗传多样性水平Table 3 Genetic diversity of 3 C.carpio populations

图1 引物SCoT34在3个黄河鲤群体中的PCR扩增电泳结果Fig.1 PCR amplification electrophoresis of primer SCoT34 in 3 C.carpio populations
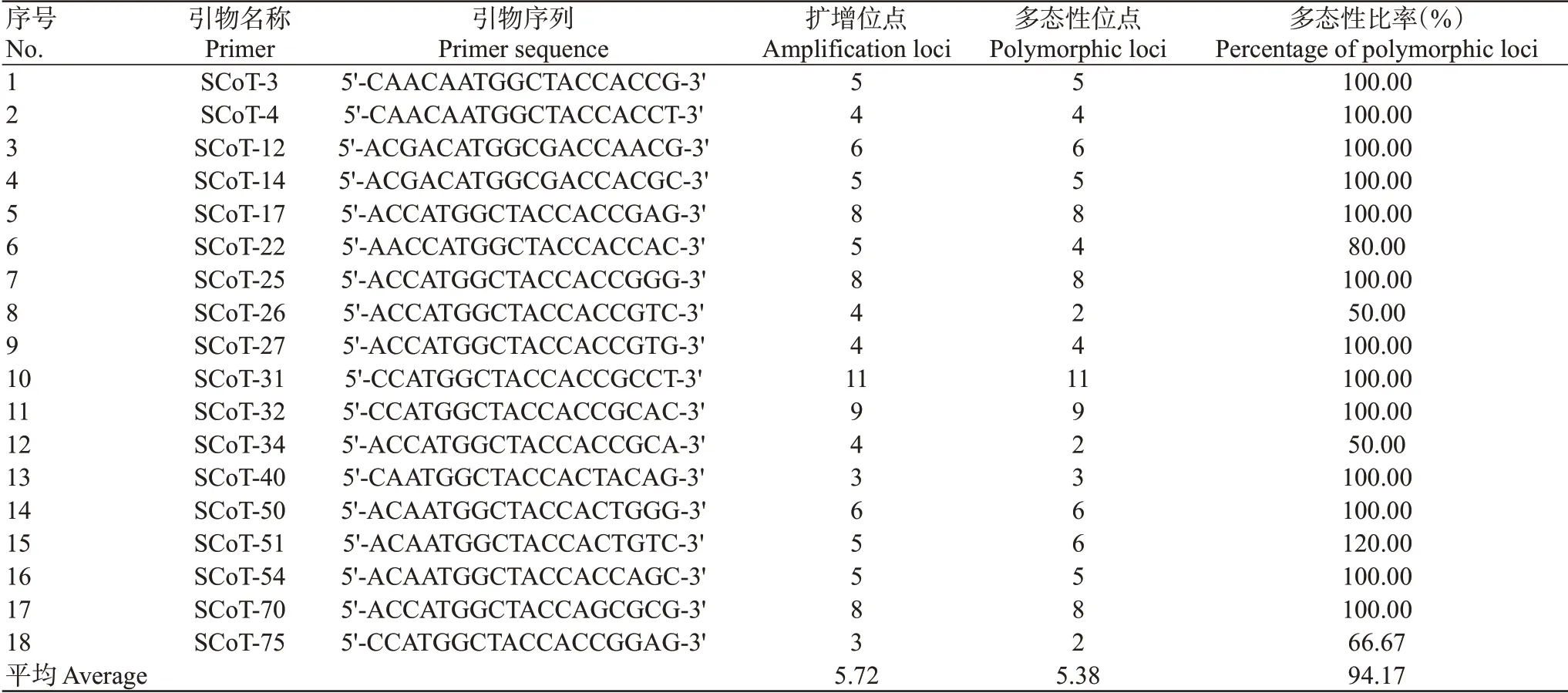
表2 18条SCoT引物的扩增结果Table 2 Amplification results of 18 SCoT primers
2.3 黄河鲤群体遗传分化分析结果
3个黄河鲤群体的AMOVA分析结果显示,群体间和群体内的方差贡献率分别为12.99%和87.01%(表4),表明群体内的遗传变异大于群体间。遗传分化指数(Fst)是衡量群体间遗传分化程度的重要参数(黄小芳等,2020;徐煜等,2021),Fst为0~0.05时表示群体间无遗传分化,Fst为0.05~0.15时表示群体间存在轻度遗传分化,Fst为0.15~0.25表示群体间存在中度遗传分化,Fst大于0.25时表示群体间存在高度遗传分化。3个黄河鲤群体的Fst为0.1299(表4),属于轻度遗传分化;群体间的Nei's遗传距离(Ds)分布在0.0248~0.0835(表5),其中FP群体与SP群体的遗传距离最大。

表4 3个黄河鲤群体的AMOVA分析结果Table 4 AMOVA analysis result of 3 C.carpio populations

表5 3个黄河鲤群体的Nei's遗传距离(对角线下)及遗传相似性(对角线上)Table 5 Nei’s genetic distance(below diagonal)and genetic similarity(above diagonal)of 3 C.carpio populations
2.4 黄河鲤群体遗传结构聚类分析结果
基于Ds的UPGMA聚类分析结果(图2)表明,FP群体和LP群体聚为一支,SP群体单独聚为一支。黄河鲤群体遗传结构分析发现,Mean lnP(K)、ΔK随着亚群数K的增加而变动,K=2.0时,Mean lnP(K)出现最大拐点和ΔK最大值(图3),因此认为K=2.0是最可能的模型,推测3个黄河鲤群体可分为2个亚群(FP和SP)。其中,LP群体由FP群体和SP群体集合而成(图3)。

图2 基于Ds构建的3个黄河鲤群体UPGMA聚类树Fig.2 UPGMA clustering tree of 3 C.carpio populations based on Ds

图3 3个黄河鲤群体的遗传结构分析结果Fig.3 Genetic component analysis of 3 C.carpio populations
3 讨论
物种的进化潜力及抵御不良因素的能力取决于种内遗传变异程度(Gibson,2021)。杂合度及群体多态性信息含量越高的生物对环境的适应能力越强,在生长、繁殖和抗逆等方面更具优势。Na和Ne是分析群体遗传多样性的重要指标(任建功等,2021)。本研究中,SP群体的Ne均高于FP群体和LP群体,说明相对于存活群体,死亡群体的遗传多样性有所下降。多态性比率能直观反映群体的多样性程度。3个黄河鲤群体的平均多态性比率为94.17%,具有较高的遗传多态性,尚未受到高强度的人工选择。FP群体、LP群体的多态性比率分别为79.61%和81.55%,SP群体的多态性比率为83.50%,表明存活群体较死亡群体具有更高的遗传多态性。在本研究中,多态性比率、Ne及I等遗传参数均随着黄河鲤群体抗病性的提高而增大,故推测黄河鲤抗嗜水气单胞菌的能力与其遗传多样性呈正相关。朱其建等(2013)基于微卫星分子标记对罗氏沼虾抗病选育群体进行遗传多样性分析,结果显示抗病能力较强的4个群体遗传多样性最丰富。有关蚂蚁(Formica selysi)(Reber et al.,2008)、小麦(Triticum aestivum)(Zhan et al.,2013)等生物群体遗传多样性的研究也表明遗传多样性高的群体抗病力更强。还有研究表明,种群遗传多样性能增强群体遭受疾病或其他自然灾害后的恢复力(Hajjar et al.,2008;Grettenberger and Tooker,2015;Yang et al.,2019)。可见,在面对同样的疾病胁迫时遗传多样性高的群体有望获得更高的产量和收益。
遗传距离能有效反映物种间的遗传变异(梁业松等,2022)。本研究结果表明,FP群体与SP群体间的Ds最大(0.0835),而FP群体与LP群体间的Ds最小(0.0248),说明2个死亡群体间的遗传分化程度相对较低。AMOVA分析结果与群体间的Gst相一致,均表明黄河鲤的遗传变异主要存在于群体内,群体间无明显的遗传分化。基于Ds的UPGMA聚类分析结果显示,FP群体和LP群体聚为一支,SP群体单独聚为一支,也表明2个死亡群体间的亲缘关系较近,其抵抗嗜水气单胞菌的能力相对较弱。利用Structure 2.3对黄河鲤群体的种群遗传结构进行检测,认为K=2.0是最可能的模型,即3个黄河鲤群体可分为2个亚群,即抗病群体(SP)和易感群体(FP和LP)。其中,LP群体由SP群体和FP群体集合而成,表明通过黄河鲤死亡时间划分的群体与通过聚类分析及遗传结构分析得出的群体基本一致。
目前,因嗜水气单胞菌感染引起的败血病在黄河鲤群体中频繁暴发,已成为黄河鲤养殖产业的主要病害之一(Jiang et al.,2016)。遗传同质化群体较遗传多样化群体更易受到病原体感染,即保持群体遗传多样性可减缓疾病对种群造成的危害(King and Lively,2012)。与野生种群相比,养殖鱼类的遗传多样性通常较低,是由于近交或针对某些重要的经济性状进行人工选择所造成。因此,开展黄河鲤抗病育种时应加强对其抗病基因的研究,提高抗病育种亲本选配的目标性,更好地利用抗病基因资源培育抗病新品种(系)。
4 结论
黄河鲤抗嗜水气单胞菌的能力随着群体遗传多样性的增大而增强。因此,在黄河鲤抗病品种(系)选育过程中应保证足够的群体数量,在提高生长、营养等经济性状的同时保证一定的基因杂合度。
猜你喜欢
今日农业(2022年4期)2022-06-01
当代水产(2022年3期)2022-04-26
海洋石油(2021年3期)2021-11-05
非常规油气(2021年1期)2021-04-01
中国现代医药杂志(2020年10期)2020-12-14
世界科学技术-中医药现代化(2020年2期)2020-07-25
中成药(2017年9期)2017-12-19
浙江农业学报(2017年1期)2017-05-17
上海农业学报(2017年3期)2017-04-10
中华老年口腔医学杂志(2016年2期)2017-01-15
