
河谷地形对草地贪夜蛾种群迁移的生态学效应
2022-02-21 12:48李永萍徐全元
云南大学学报(自然科学版) 2022年1期
叶 辉,李永萍,冯 丹,徐全元
(1.云南大学 生态与环境学院,云南 昆明 650091;2.云南大学 农学院,云南 昆明 650091;3.云南省林业和草原科学院,云南 昆明 650201;4.西南林业大学 大数据与智能工程学院,云南 昆明 650224)
草地贪夜蛾属世界性重大农业迁飞昆虫,于2019 年首次经云南迁入中国,当年扩散至全国26个省区,发生面积达100 万hm2·次(1 500 万亩·次),是目前云南乃至全国位居首位的玉米害虫,对中国玉米种植业构成巨大威胁[1-3].
迁飞被认为是草地贪夜蛾得以在中国大范围散布、大规模危害的归因[4-6].草地贪夜蛾没有滞育现象,进入秋季因气温下降,随气流向南方温暖地区迁移,以此避免冬季低温胁迫影响;次年春季随气温回升,逐渐从越冬地向北迁移,由此扩大地理分布,造成大范围大规模危害[6-9].草地贪夜蛾迁移活动既具有内在遗传学基础,也受制于外界环境条件[10-11].环境因素如气温气流、地形地貌及寄主等,对草地贪夜蛾迁飞行为具有明确的启动与导向作用[10,12-13].探究环境因素的作用强度及激发机制,有助于揭示并阐释草地贪夜蛾迁移规律[6].
河谷是地表形态的重要地形特征[14].云南河谷数量众多,仅体量较大的河谷就有独龙江河谷、怒江河谷、澜沧江河谷、金沙江河谷、元江河谷、南盘江河谷等[14-15].云南河谷的多样性及广泛性构成了云南独特的河谷景观,对河谷流域自然地理特征、生态环境条件、生物多样性及其演变过程具有决定性影响[15-16].研究表明,河谷地形特征与河谷流域气候特征形成共轭效应,影响着昆虫种群的数量变动模式和空间分布格局[17-18].如桔小实蝇、番石榴实蝇等不少入侵昆虫均是以河谷为通道向内地扩散迁移的,河谷流域也因其适宜的气温环境成为来自热区昆虫的主要栖息地或重要越冬场所[17,19-20].河谷对于外来入侵昆虫在云南的发生与发展具有重要的生态学意义.
本研究拟从自然地理的视角,遵循生态学思想,从河谷流域的综合自然特征入手,探讨草地贪夜蛾在河谷区的迁移扩散规律,探究河谷特殊地形地貌对草地贪夜蛾种群迁移扩散行为的作用机制,以此拓宽和深化对河谷条件下草地贪夜蛾种群发展过程的认知,为阐释云南草地贪夜蛾发生发展过程提供新视野和新思想.
1 河谷对草地贪夜蛾迁移的通道效应
河谷呈条带状延伸,河谷内生态环境条件大体相同或相近,在景观生态学上可称为生态廊道,对生物扩散迁移具有通道作用[15,21].通道现象在怒江河谷最为典型,来自中南半岛经云南西南边境入侵的外来生物,大多沿怒江、澜沧江向内地扩散迁移[18,21].在此,我们以该两江河谷为例分析探讨河谷对草地贪夜蛾迁移的通道效应.
怒江位于云南西部,省内北起丙中洛,流经怒江州、保山市和德宏州,在德宏中山乡进入缅甸,云南省境内全长约650 km[22](图1).怒江分上下游两段,以泸水市为界,怒江上游穿行于碧罗雪山与高黎贡山之间,河谷狭窄,两岸耸峙,岭谷平均高差逾3 000 m;怒江下游河谷渐次开阔,地势趋缓,平均高差仅约500 m[23].
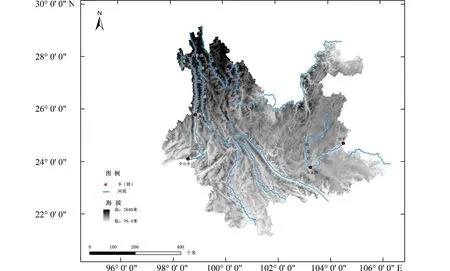
图1 云南省主要河流地理分布Fig.1 Map of distribution of main rivers in Yunnan Province
怒江流域是云南草地贪夜蛾主要发生区之一[1,9].怒江下游冬季气温暖和,玉米四季轮作,是草地贪夜蛾周年发生区;怒江上游地区冬季气温偏低,玉米种植以春夏季为主,草地贪夜蛾不能就地越冬,是草地贪夜蛾季节发生区[9,24].周年发生区与季节发生区大体以泸水古登乡为界.每年3─4 月,草地贪夜蛾随春季气温回升逐渐从周年发生区向怒江上游迁移,途经福贡、贡山,于7 月抵达怒江北界——贡山丙中洛,怒江河谷是滇西南周年发生区草地贪夜蛾向季节发生区的主要迁移通道[24-25].
研究表明,草地贪夜蛾在澜沧江流域表现出与在怒江河谷同样的迁移过程,相关研究结果将以另文做详细报道.实际上,怒江与澜沧江都是云南西部地区草地贪夜蛾南北迁移的主要通道,在外来入侵生物向内地传输过程中发挥重要作用[21-25].除此之外,独龙江河谷、元江河谷,以及金沙江和南盘江河谷的部分区段,均是草地贪夜蛾的主要迁移路径,河谷通道效应十分明显[26].云南河谷众多,形态多样,河谷迁移是草地贪夜蛾对河谷形态的生态行为适应的行为特征,河谷通道效应在云南具有显著普遍的生态学意义.
如何理解河谷通道效应的生态学机制呢?首先,两山夹一谷的河谷地貌特征,在地形上阻隔了草地贪夜蛾向河谷两侧扩散,从客观上确定了草地贪夜蛾的行进方向,河谷迁移特别在高山峡谷地区成为草地贪夜蛾行进的无二选择.由此可见,沟槽状地表构型这一河谷地形特征是河谷通道效应的主导因素.其次,云南西部主要受西南和东南季风气流控制,季风气流在纬向河谷内由南向北运行[6,15].河谷气流所形成的动能为草地贪夜蛾迁移提供了驱动力,促进或推动了草地贪夜蛾的北向迁移.研究表明,草地贪夜蛾在正常情况下一次可迁移30 km/d,在有气流驱动作用下,可飞行至480 km之外,气流在草地贪夜蛾迁飞过程中发挥重要作用[10,27].河谷气流一方面加速了草地贪夜蛾的迁移进程,草地贪夜蛾在气流作用下可以飞得更远,同时气流方向也在很大程度上确定了草地贪夜蛾迁移的方向.简言之,河谷地形构成草地贪夜蛾迁移的物理通道,河谷气流为草地贪夜蛾迁移提供了驱动力,两者形成的叠加效应,构成草地贪夜蛾河谷迁移的生态学基础.
从草地贪夜蛾防控看,河谷地形相对狭窄,河谷迁移方向明确,这就为开展草地贪夜蛾防控提供了理想场所[9].河谷防控应作为草地贪夜蛾迁移防控的重点地区,把阻截防控作为河谷防控的基本对策.就怒江河谷而言,草地贪夜蛾防控指导思想是,将草地贪夜蛾控制于怒江下游的周年发生区,在怒江下游至上游区段,也即在从周年发生到季节发生的过渡区域内构建防控阻截区(带),将草地贪夜蛾阻截于向季节发生区迁移的途中,从而把季节发生区内的草地贪夜蛾危害降到最低程度,以此起到事半功倍的防控效果[9,24].
2 河谷对草地贪夜蛾迁移的阻隔或阻滞作用
河谷对于草地贪夜蛾的阻隔主要表现为河谷两侧高山的阻挡作用.阻隔效应与通道效应,是河谷生态学效应中看似相互矛盾,实则相互依存的两个方面.河谷沟槽地形构造一方面为草地贪夜蛾提供了沿河谷迁移的通道,同时也成为了草地贪夜蛾向河谷两侧扩散的屏障.换言之,草地贪夜蛾沿河谷走向迁移,已表明其向河谷两侧扩散受到制约;反过来讲,也正是草地贪夜蛾向河谷两侧扩散受到规束,才出现草地贪夜蛾沿河谷迁移的现象.
河谷对于草地贪夜蛾的通道效应与河谷阻隔作用,在如同怒江河谷、澜沧江河谷这类纵向河谷中十分明显.前面讲到,草地贪夜蛾以怒江河谷和澜沧江河谷为主要迁移路线,该两河谷相向并行,相距最近距离仅有19 km,但两河谷种群的迁移进度并不尽一致.调查发现澜沧江流域中排乡3 月18 日首次见虫,比同纬度怒江的福贡上帕镇早20 d左右,说明澜沧江流域草地贪夜蛾迁移进度明显要快于其在怒江流域.河谷草地贪夜蛾在相距极近的两河内迁移进度差异,说明其在两河谷的迁移过程相互独立,没有或极少有相互渗透或交流,暗示两河谷种群受到山脉所阻隔,河谷阻隔作用显然可见[21].
河谷阻隔作用不仅出现在纵向岭谷区的怒江、澜沧江等河谷区,也反映在元江河谷以及金沙江、南盘江等河谷的部分区段,这些河谷或河谷区段均呈南北走向,与草地贪夜蛾迁移路线一致[28].
阻隔作用除了存在于南北向伸展的河谷中,也出现在东西向延伸的河谷内.在云南东部的南盘江从开远小龙潭到罗平区段,河谷大体呈东西向走向,江面海拔平均为1 000 m,江南为喀斯特丘陵地貌,平均海拔为700~1 500 m;江北为滇东高原,平均海拔为2 000 m,山谷高差显著,平均为1 700 m 左右(图1).该区域3—8 月的主控气流为东南季风,草地贪夜蛾迁移由南向北进行[1].由此,从方位上看,南盘江北边曲靖的草地贪夜蛾易于从南边的红河州过来,但对当地过去几年监测数据发现,南盘江北边与曲靖相邻区域的草地贪夜蛾发生量较少,出现时间也较晚.实地调查分析发现,草地贪夜蛾进入南盘江河谷后,面临北边云岭山脉南段余脉,如菌子山、白腊山等阻挡,迁移种群需要逐渐从海拔为700 m 左右河谷面,向海拔为2 000 m 左右高原面爬升,谷坡高山阻滞了种群的迁移进度,也消耗了迁移种群数量.所以,正是受到南盘江北面山脉的阻滞作用,草地贪夜蛾在从红河州跨越南盘江北上曲靖的过程中,种群量减少,迁移速度趋缓.
河谷对迁移种群的阻滞作用在很大程度上是由于河谷高差造成的.河谷高差愈大,对迁移过程的影响就愈大.由于云南河谷众多,山高谷深,谷岸高差现象极为普遍[14,21,29-30].河谷对草地贪夜蛾迁移阻隔或阻滞的现象广泛存在,但作用强弱因地而异,在一些地方较为显著,而在另一些地方或流域相对较弱[25,28].
进一步分析认为,河谷阻隔不仅限于山脉的物理屏障作用,还包括河谷气流、气温、寄主种植等多种因素.气流沿河谷走向运行,对草地贪夜蛾迁移现成定向驱动力,势必影响到草地贪夜蛾向河谷两侧移动.河谷气温随两岸海拔增加而下降,低温条件将影响草地贪夜蛾的迁飞行为.在没有寄主玉米种植的情况下,草地贪夜蛾种群将因食物缺失难以继代迁移.这些因素从客观上可视为具有不同生态学内涵的屏障,表现出不同程度的阻隔效应.从这一意义上讲,河谷阻隔或阻滞作用是河谷条件下形成或衍生的具有生态阻隔作用的多种因素的综合反映.
需要指出,河谷对草地贪夜蛾迁移的阻隔或阻滞作用,是针对迁移种群环境因素所讲的,是影响草地贪夜蛾迁移的外因.草地贪夜蛾迁移还涉及到迁移种群大小,迁飞高度等内在因素.河谷的阻隔或阻滞作用是迁移种群外部和内部两方面共同作用的结果.有关迁移种群内因部分不在本文讨论之内,不予深入讨论.
河谷对草地贪夜蛾种群迁移的影响既有完全阻隔,也表现有迟滞的阻滞效应,阻隔或阻滞的不同反映了阻挡作用的大小.在草地贪夜蛾区域防控实践中,可以把河谷阻隔阻滞作用作为生态防控因素加以利用.
3 河谷对草地贪夜蛾种群的汇聚效应
这里讲的汇聚效应,是指草地贪夜蛾在河谷的聚集现象.在云南,草地贪夜蛾河谷聚集现象较为普遍,尤其在冬季更为明显[9,12].进入秋季之后,随气温下降,草地贪夜蛾逐渐由高海拔向低海拔移动,由高纬度向低纬度地区迁移,趋向冬季气候温暖的地方[4,9].河谷区域海拔较低,热量充沛,往往是草地贪夜蛾越冬的理想栖息地[9,13].研究发现,怒江河谷、澜沧江河谷、元江河谷,以及金沙江河谷等,都是冬季草地贪夜蛾汇聚的地方[12].
河谷汇聚现象是草地贪夜蛾对河谷环境条件适宜性选择的结果,体现了河谷环境对草地贪夜蛾行为活动的生态学效应.其首先是河谷的热量因素.云南位于低纬高原区,低纬度较之高纬度可以获取更多热量,而高原较之低海拔地区气温明显偏低.例如,昆明位于北纬24.38°~26.36°,年均温为12~22 ℃,而在同纬度的泉州年均温为20~26 ℃[29].正是云南低纬高原区所形成的特定气温条件,使得当地草地贪夜蛾冬季分布北界在纬度上明显低于中国东南沿海地区[12].如果在云南高原面上,海拔随河谷深切而下降,就形成了低纬低海拔区域,河谷气温显著提升.金沙江河谷从元谋至巧家段,平均海拔为700~950 m,冬季最冷月平均气温为9~22 ℃,河谷区气温明显高于区外气温[29].越冬区调查发现,冬季草地贪夜蛾沿河谷延伸呈带状分布,其与河谷热量分布相吻合,反映了河谷热量对草地贪夜蛾河谷分布影响的内在关联[9,30].
河谷热量分布样式取决于河谷所在地理位置及地形地貌形态,从全区域审视形如热岛,其在冬季谷区外气温较低情况下十分明显.河谷热岛为草地贪夜蛾越冬提供了热量,是冬季草地贪夜蛾种群得以在河谷区生存繁衍的主要原因[13].
寄主是草地贪夜蛾河谷汇聚的另一重要因素.云南草地贪夜蛾在生物型上属玉米型,即主要以取食玉米为生,玉米是云南草地贪夜蛾生存发展的前提条件[9].近年大量野外调查表明,由于河谷区热量充沛,当地农户大多有冬季种植玉米的习惯,有的地方如金沙江河谷区的元谋、巧家等地,玉米种植四季不断,为草地贪夜蛾提供了充足延续的食物.河谷热量是河谷区位与地形地貌所致,而玉米种植则是农户利用河谷热量增加土地收益的农事活动,是河谷生态条件的衍生.如果说河谷热量为草地贪夜蛾汇聚提供了热量保障,河谷冬季玉米种植则为冬季草地贪夜蛾提供了食物基础.河谷热量与河谷玉米解释为草地贪夜蛾得以在河谷越冬的关键原因,河谷也因此成为草地贪夜蛾的冬季庇护地.
调查表明,金沙江河谷区是草地贪夜蛾周年发生区在云南高原的冬季发生区飞地,也成为云南北部地区草地贪夜蛾春季发生的主要虫源地.由于该区域位于云南北部季节发生区内,春季扩散较之源于南部周年发生区的草地贪夜蛾出现时间早,使得在该区域开展春季草地贪夜蛾防控更加困难.
河谷防控是云南草地贪夜蛾区域防控的重要组成,应根据草地贪夜蛾河谷汇聚的特点,采取相应的防控对策.基于河谷区草地贪夜蛾冬季汇聚、春季扩散的情况,防控工作应在冬春季同步展开.冬季防控以围歼为主,主要是针对草地贪夜蛾冬季种群聚集于河谷区的特点,采用压缩草地贪夜蛾冬季分布区域,降低草地贪夜蛾冬季种群数量的防控策略.春季防控应将围歼防控与阻截防控相结合,旨在阻截草地贪夜蛾从越冬地向河谷区外扩散迁移,减少草地贪夜蛾向谷外的扩张危害.冬季防控与春季防控相统筹,围歼防控与阻截防控相结合,将草地贪夜蛾控制于河谷区域内,减少或避免其向谷区外扩散,把草地贪夜蛾危害降低到最小程度,以降低谷区及谷外区域玉米受害损失,应作为该类河谷防控的指导思想和关键举措.
猜你喜欢
江西农业学报(2022年4期)2022-06-28
云南画报(2021年4期)2021-07-22
云南画报(2020年12期)2021-01-18
云南画报(2020年11期)2020-12-14
诗选刊(2020年10期)2020-10-27
科技资讯(2018年1期)2018-06-05
植物保护(2018年3期)2018-05-14
青年文学家(2011年2期)2011-06-08
中国青年(1984年2期)1984-08-20
