
3个种源池杉幼苗对干旱胁迫的生理响应及抗旱性评价
2022-03-01 18:11覃奎刘秀马道承王凌晖
山东农业科学 2022年1期
覃奎 刘秀 马道承 王凌晖







摘要:本试验探究不同种源池杉幼苗的抗旱性差异,以期为其引种驯化、抗旱种源筛选与培育、消落带植被选择及池杉抗旱性育种提供一定的参考。利用不同浓度PEG-6000与Hoagland的混合液模拟不同程度干旱胁迫,测定干旱胁迫对3个种源池杉幼苗渗透调节物质和抗氧化特性的影响,并利用抗旱指数与主成分分析、隶属函数相结合的方法对其抗旱性进行综合评价。结果表明:3个种源池杉幼苗的丙二醛、可溶性糖、可溶性蛋白含量以及超氧化物歧化酶和抗坏血酸过氧化物酶活性均随着干旱胁迫程度的增大而逐渐升高;浙江安吉和江西分宜种源的叶绿素a、b及总含量均随着干旱胁迫程度的增大而逐渐升高,而广西凭祥种源的含量则呈先下降再上升的趋势;浙江安吉和广西凭祥种源的过氧化物酶活性随着干旱胁迫程度的增大而逐渐升高,而江西分宜种源的则呈先上升再下降的趋势。经综合评价,3个种源池杉幼苗的抗旱性强弱表现为浙江安吉>江西分宜>广西凭祥。
关键词:池杉;幼苗;干旱胁迫;渗透调节物质;抗氧化酶;抗旱性评价
中图分类号:S791.34 文献标识号:A 文章编号:1001-4942(2022)01-0030-08
随着全球气候变暖加剧,极端天气频频发生,干旱已经成为影响植物正常生长发育的主要非生物胁迫因子之一[1]。研究表明,干旱作为一种多维胁迫,可以引起植物表型、生理学、生物化学和分子水平的一系列变化[2],植物对干旱胁迫的适应机制与其形态结构、光合作用、渗透调节、抗氧化酶[3-6]等密切相关,一直是研究的热点。目前对植物抗旱性鉴定的方法主要有田间直接鉴定法、人工控水鉴定法和高渗透溶液模拟干旱鉴定法等[7],其中高渗透溶液模拟干旱鉴定法广泛应用于植物不同品种及种源间的抗旱性研究[8-10]。对植物品种及种源的抗旱性评价方法主要有分级评价法、隶属函数法、抗旱指数法、主成分分析法和灰色关联度法等[11]。近年来,熊仕发[12]、蔡建國[13]等越来越多的学者更倾向于将多种评价方法相结合,以提高抗旱性评价的准确性。
池杉[Taxodium distichum var. imbricatum(Nuttall)Croom]是杉科(Taxodiaceae)落羽杉属(Taxodium)植物,亦称池柏、沼落羽松。其树干通直,材质好,是我国重要的造林、园林树种。因兼具生长快、耐水耐旱、抗风、病虫害少、易繁殖等特点,池杉已成为构建消落带植被的优良适生乔木。目前国内外学者对池杉的研究主要集中于繁育栽培及造林[14]、化学成分[15],另外,在对其水分生理的研究中,耐淹性的研究也较为突出[16,17],但关于其耐旱性的研究目前仍较匮乏,尚缺乏较为成熟的水分耐受性评价体系。在引种驯化及适生性评价方面,不同种源池杉对水分的耐受性仍不甚了然,加之消落带常呈现出周期性的“水淹-落干-水淹”交替的水位变动规律[18],干旱是其遭遇频度较高的生境胁迫之一。因此,本试验以不同种源池杉为研究对象,通过在霍格兰(Hoagland)营养液中添加不同浓度聚乙二醇(PEG),开展不同梯度的干旱胁迫试验,从光合色素含量、渗透调节物质积累、抗氧化酶活性方面分析不同种源池杉对干旱的响应,而后采用抗旱指数、主成分分析及模糊隶属函数相结合的方法对不同种源的池杉进行抗旱性综合评价,筛选出抗旱性较强的种源,以期对其引种驯化、抗旱品种选育、消落带植物选择提供一定的参考。
1 材料与方法
1.1 试验地概况
本试验于广西壮族自治区林业科学研究院苗圃基地(108°35′E,22°92′N)进行。该地属于亚热带季风气候,年平均气温21.6℃左右,40.4℃为极端最高温,-2.1℃为极端最低温;年均降水量约为1304.2mm,雨水主要集中在夏季,年平均相对湿度为80%,干湿季分明,夏季潮湿,冬季干燥。试验地全年日照时间较长,雨水充足。
1.2 试验材料
供试植物为浙江安吉(ZJAJ)、江西分宜(JXFY)、广西凭祥(GXPX)3个种源的池杉幼苗。2019年11月分别从浙江安吉县龙山林场、江西分宜县中国林业科学研究院亚林中心、广西凭祥市中国林业科学研究院热带林业实验中心采集3个种源的池杉种子,带回广西壮族自治区林业科学研究院苗圃基地干藏,2020年3月播种于该基地苗圃沙床中,待种子萌发且长势良好后,移入12cm×10cm的育苗杯中,每杯一株,基质为纯椰糠,置露天培育,定时定量浇水,自然光照,待苗高40cm左右时,选取生长一致的幼苗进行试验。
1.3 试验方法
试验在Hoagland营养液中分别添加6个比例梯度的PEG-6000:0(CK)、5%、10%、15%、20%、25%,每处理10个生物学重复,每重复1株幼苗,每种源共需60株幼苗,3个种源合计180株幼苗。2021年6月,将3个种源的池杉幼苗从育苗杯中取出,先用清水洗净附着于根系表面的栽培基质及其他杂质,立即置于Hoagland完全营养液中平衡3天,之后将幼苗根部完全浸入装有不同浓度PEG-6000与Hoagland混合液的烧杯中进行模拟干旱胁迫,CK则将幼苗根部完全浸入装有纯Hoagland完全营养液的烧杯中。处理2天后采样,进行各项生理指标的测定。
1.4 测定指标及方法
选取每处理组中长势一致的植株,采摘相同部位成熟功能叶,剪去主脉,测定各生理指标,每处理重复3次。叶绿素含量采用丙酮-乙醇混合浸提法测定,丙二醛(MDA)含量采用硫代巴比妥酸法测定,可溶性糖含量采用蒽酮比色法测定,可溶性蛋白含量采用考马斯亮蓝-G250染色法测定,超氧化物歧化酶(SOD)活性采用氮蓝四唑光还原法测定,过氧化物酶(POD)活性采用愈创木酚法测定,抗坏血酸过氧化物酶(APX)活性采用紫外分光光度法测定。具体操作均参考李合生[19]的方法。
1.5 数据处理及统计分析
采用MicrosoftWord和Excel2016进行数据处理、分析及图表绘制,用SPSS19.0进行统计分析、差异显著性检验(Waller-Duncan’s法)和主成分分析。
为消除池杉种源间基础性状的差异,参考蔡建国[13]、谢小玉[20]、李素[21]等的方法,引入抗旱指数这一相对指标值对3个种源池杉的抗旱性进行综合评价及分析。公式如下:
2 结果与分析
2.1 干旱胁迫对池杉幼苗叶绿素含量的影响
由图1可知,随着干旱胁迫程度的增大,ZJAJ和JXFY种源的池杉幼苗叶绿素a、叶绿素b、总叶绿素含量逐渐升高,而GXPX种源的各叶绿素含量先下降再上升。与CK相比,ZJAJ和JXFY种源的各叶绿素含量在各浓度处理下均显著增加(P<0.05)。在5% PEG-6000处理下,ZJAJ和JXFY种源的叶绿素a、叶绿素b、总叶绿素含量较CK增幅均最小,且ZJAJ种源的增幅略高于JXFY,分别为37.74%、29.41%、33.33%,但两种源间无显著差异(P>0.05)。在10% PEG-6000处理下,GXPX种源的叶绿素a、叶绿素b、总叶绿素含量较CK降幅最大,分别为35.29%、24.65%、32.71%。在25% PEG-6000处理下,3个种源的叶绿素a、叶绿素b、总叶绿素含量较CK增幅均达到最大值,其中ZJAJ种源的增幅最大,分别为141.51%、85.29%、119.54%;GXPX的增幅最小,分别为75.00%、51.28%、66.36%;3个种源的叶绿素a和总叶绿素含量差异显著(P<0.05)。
2.2 干旱胁迫对池杉幼苗渗透调节物质的影响
2.2.1 可溶性糖含量 由图2可知,随着干旱胁迫程度的增大,3个种源的可溶性糖含量均呈逐渐升高的趋势。与CK相比,3个种源的可溶性糖含量除5% PEG-6000处理外,其余各处理均显著升高(P<0.05)。在5% PEG-6000处理下,3个种源的可溶性糖含量较CK增幅均最小,ZJAJ、JXFY、GXPX分别为10.42%、5.66%、4.81%,三者间无显著差异(P>0.05)。在25% PEG-6000处理下,3个种源的可溶性糖含量较CK增幅均最大,GXPX、ZJAJ、JXFY分别为219.23%、117.71%、89.62%,GXPX種源的可溶性糖含量显著高于后两者(P<0.05),而ZJAJ和JXFY种源间的可溶性糖含量无显著差异(P>0.05)。
2.2.2 可溶性蛋白含量 由图3可知,随着干旱胁迫程度的增大,3个种源的可溶性蛋白含量均呈逐渐升高的趋势。与CK相比,ZJAJ种源的可溶性蛋白含量在PEG-6000各浓度处理下均显著高于CK(P<0.05);JXFY种源的可溶性蛋白含量除5% PEG-6000处理外,其余各处理均显著高于CK(P<0.05);GXPX种源的可溶性蛋白含量仅在20%和25% PEG-6000处理下显著高于CK(P<0.05),其余各处理虽高于CK,但无显著差异(P>0.05)。在5% PEG-6000处理下,3个种源的可溶性蛋白含量较CK增幅均最小,ZJAJ、JXFY、GXPX 分别为7.73%、2.52%、1.75%,且三者间差异显著(P<0.05)。在25%PEG-6000处理下,3个种源的可溶性蛋白含量较CK增幅均最大,GXPX、ZJAJ、JXFY分别为60.50%、40.77%、40.48%,GXPX种源的可溶性蛋白含量显著高于后两者(P<0.05),ZJAJ和JXFY间无显著差异(P>0.05)。
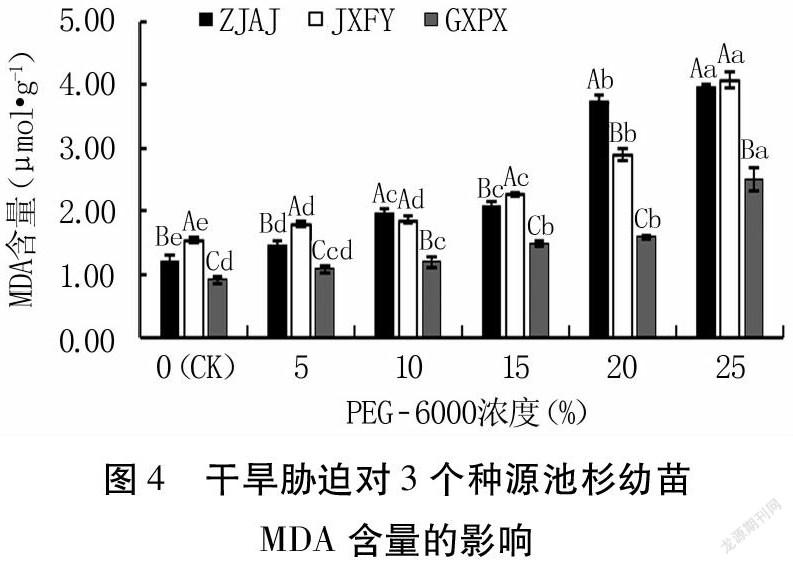
2.3 干旱胁迫对池杉幼苗MDA含量的影响
由图4可知,随着干旱胁迫程度的增大,3个种源的MDA含量均呈逐渐升高的趋势。与CK相比,ZJAJ和JXFY种源的MDA含量在PEG-6000各浓度处理下均显著增加(P<0.05);GXPX种源的MDA含量在5% PEG-6000处理下虽高于CK,但差异不显著(P>0.05),其余各浓度处理下则均显著高于CK(P<0.05)。在5% PEG-6000处理下,3个种源的MDA含量较CK增幅均最小,ZJAJ、GXPX、JXFY分别为20.00%、17.78%、16.34%,且三者间差异显著(P<0.05)。在25%PEG-6000处理下,3个种源的MDA含量较CK增幅均最大,ZJAJ、GXPX、JXFY分别为230.00%、176.67%、166.67%,且GXPX的MDA含量显著低于ZJAJ和JXFY(P<0.05),而ZJAJ和JXFY间无显著差异(P>0.05)。
2.4 干旱胁迫对池杉幼苗抗氧化酶活性的影响
2.4.1 SOD活性 由图5可知,3个种源的SOD活性均随着干旱胁迫程度的增大而逐渐升高。与CK相比,3个种源的SOD活性在PEG-6000各浓度处理下均显著升高(P<0.05),且GXPX种源的增幅均大于其余两个种源。在5% PEG-6000处理下,3个种源的SOD活性较CK增幅均最小,GXPX、ZJAJ、JXFY分别为7.73%、2.52%、1.75%;ZJAJ种源的SOD活性显著高于JXFY(P<0.05),其余两两种源之间无显著差异(P>0.05)。在25% PEG-6000处理下,3个种源的SOD活性较CK增幅均最大,GXPX、JXFY、ZJAJ分别为164.52%、149.02%、118.78%;GXPX种源的SOD活性显著高于ZJAJ和JXFY(P<0.05),而ZJAJ和JXFY间无显著差异(P>0.05)。
2.4.2 POD活性 由图6可知,随着干旱胁迫程度的增大,ZJAJ和GXPX种源的POD活性均逐渐升高,而JXFY种源的POD活性则呈先升高再降低的趋势。在CK处理下,JXFY种源的POD活性显著高于ZJAJ和GXPX(P<0.05),而ZJAJ和GXPX间无显著差异(P>0.05)。在5% PEG-6000处理下,ZJAJ、JXFY、GXPX种源的POD活性较CK增幅分别为125.85%、109.97%、41.67%,JXFY种源的POD活性显著高于ZJAJ和GXPX(P<0.05),而后两者间无显著差异(P>0.05)。在15% PEG-6000处理下,JXFY种源的POD活性达到峰值,较CK增加171.20%,显著高于其余两个种源。在25% PEG-6000处理下,JXFY种源的POD活性最低,比CK低15.00%,但无显著差异;ZJAJ和GXPX种源的POD活性则达到最大值,分别比CK高464.67%、266.67%;3个种源间POD活性存在显著差异(P<0.05),ZJAJ种源最高,JXFY种源最低。
2.4.3 APX活性 由图7可知,3个种源的APX活性均随着干旱胁迫程度的增大而逐渐升高。与CK相比,ZJAJ和JXFY种源的APX活性在PEG-6000各浓度处理下均显著升高(P<0.05),而GXPX种源在5%和10% PEG-6000处理下升高不显著(P>0.05)。在5% PEG-6000处理下,3个种源的APX活性较CK增幅均最小,ZJAJ、JXFY、GXPX分别比CK高19.58%、16.93%、13.53%,但3个种源间无显著差异(P>0.05)。在25% PEG-6000处理下,3个种源的APX活性较CK增幅均最大,ZJAJ、JXFY、GXPX分别比CK高79.90%、75.40%、55.43%,但3个种源间无显著差异(P>0.05)。
2.5 不同种源池杉抗旱性综合评价
为消除池杉种源间基础性状的差异,通过计算指标的相对值求得抗旱指数(表1),并基于此进行主成分分析,得到2个特征值大于1的主成分,贡献率分别为75.79%、24.21%,累计贡献率达100.00%,保留了原始性状的全部信息(表2)。在主成分1中,叶绿素a、叶绿素b、总叶绿素等具有较大的正载荷,说明主成分1主要与叶片光合色素含量的变化等相关,可称为光和色素因子;在主成分2中,POD、可溶性糖、MDA、SOD等具有较大的正载荷,说明该主成分主要与植物体内的抗氧化酶和渗透调节物质等相关,可称为防御调节因子。经计算得到各种源的抗旱性综合评价值D(表3),据此判断3个种源池杉的抗旱性由大到小依次为ZJAJ>JXFY>GXPX。
3 讨论与结论
叶绿素是植物类囊体膜上色素蛋白复合体的重要组成部分,是光合作用的基础物质和主要影响因子,其含量高低可在一定程度上衡量植株的抗逆性[22]。本研究发现ZJAJ和JXFY种源的叶绿素a、叶绿素b、总叶绿素含量均随着干旱胁迫程度的增大而逐渐升高,这一方面可能与干旱胁迫下叶片的相对含水量下降有关,另一方面也反映了池杉在干旱胁迫下通过增加叶绿素含量来促进光合有机物的积累及渗透调节物质的合成,以更好地适应干旱环境,这与李鑫[23]、高昆[24]等研究干旱胁迫对粉葛[Puerariamontanavar.thomsonii(Bentham)M.R.Almeida]影响的结论相似。
而GXPX种源的叶绿素含量随着干旱胁迫程度的增大呈先降低再逐渐升高的趋势,这可能是该种源抗旱性较弱,轻度干旱胁迫下叶绿素合成受阻,但随着干旱胁迫加剧,植株为维持正常生长,加大了叶绿素合成量,以促进有机物的积累及渗透调节物质的合成,这与区智等[25]对灯台树[Alstoniascholaris(L.)R.Br.]的研究结论相似。
应对干旱胁迫,植物可通过自身细胞的渗透调节作用来降低渗透势,从而降低水势,使细胞可以继续从外界吸水维持膨压[26]。可溶性糖、可溶性蛋白是调节植物细胞渗透势的重要物质,能在一定程度上促进细胞内活性氧的清除。本研究中3个种源池杉的可溶性糖、可溶性蛋白含量均随着干旱胁迫程度的增加而逐渐升高,这与赵英等[27]对蟛蜞菊[Sphagneticolacalendulacea(Linnaeus)Pruski]的干旱胁迫研究结果一致。此外,本研究还发现,抗旱性较弱的GXPX种源在重度干旱胁迫下可溶性糖含量显著高于ZJAJ和JXFY,说明其在遭遇重度干旱胁迫下主要通过合成可溶性糖来适应干旱逆境。
MDA为膜脂过氧化产物,其含量可以直接反映细胞膜脂过氧化程度,从而间接反映植物的受害程度[28]。在干旱程度严重情况下,活性氧大量积累,引发细胞膜脂过氧化,从而使细胞膜受到伤害,导致MDA含量上升。本研究中,3个种源池杉的MDA含量均随着干旱胁迫程度的增大而逐渐升高,这与郭米山等[29]对国槐[Styphnolobiumjaponicum(L.)Schott]的研究结论相似。3个池杉种源均在25% PEG-6000处理下MDA含量增幅最大,表明该胁迫程度对细胞膜造成严重伤害,膜脂过氧化加重。
SOD、POD和APX是植物体内重要的抗氧化酶,可有效清除活性氧,减轻水分胁迫诱导的氧化伤害[30]。本研究中随着干旱胁迫程度的增大,ZJAJ和GXPX种源的SOD、POD、APX活性及JXFY种源的SOD和APX活性均升高,但JXFY的POD活性呈先升高再降低的趋势,这与陈梦园[31]对栓皮栎(QuercusvariabilisBlume)的干旱脅迫研究结果一致。说明当干旱胁迫超过一定程度后,活性氧清除机制可能遭到破坏,导致抗氧化酶活性下降,不足以清除活性氧,从而对植物膜系统的保护受限。
植物的抗旱性是由多种因素综合作用形成的[32],仅用某项抗旱指标难以准确、全面地反映其抗旱能力[33]。本研究通过主成分分析和隶属函数法对3个种源池杉的抗旱性进行了综合评价,结果表明ZJAJ抗旱性最强,JXFY次之,GXPX最弱。但造成这种差异的原因还有待从分子水平进一步研究。此外,本研究发现叶绿素含量、POD活性和可溶性糖含量是池杉幼苗在干旱胁迫下较为敏感的指标,这与赵瑾[34]、高晓宁[35]等的研究不尽相同,可能是不同植物对于干旱胁迫的应激机制不同导致。植物抗旱性综合评价是一个复杂的问题,不同研究筛选出的典型抗旱指标是否具有通用性,还有待进一步研究与论证。
综上,不同种源池杉幼苗对不同程度干旱胁迫表现出不同的响应。抗旱性较强的ZJAJ种源幼苗的叶绿素含量、渗透调节物质、抗氧化酶活性和MDA含量随着干旱胁迫程度的增大而逐渐升高,而抗旱性较弱的种源幼苗存在随着干旱胁迫程度增大而出现叶绿素含量和POD活性下降的趋势。经主成分分析和隶属函数法综合评价,3个种源的抗旱性由大到小依次为ZJAJ>JXFY>GXPX,可为抗旱育种应用和消落带植被栽培提供参考。
猜你喜欢
青年文学家(2022年12期)2022-05-18
诗潮(2021年1期)2021-03-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
安徽农学通报(2016年24期)2017-01-12
安徽农学通报(2016年24期)2017-01-12
现代园艺(2016年7期)2017-01-09
山东农业科学(2016年11期)2016-12-17
现代园艺(2016年2期)2016-03-15
河北渔业(2015年10期)2015-10-15
