
龙苍沟国家森林公园7种花楸属植物的叶解剖特征及其环境适应性
2022-03-02 06:51朱凯琳李嘉宝
植物研究 2022年2期
朱凯琳 李嘉宝 陈 昕*
(1. 南京林业大学南方现代林业协同创新中心,南京 210037;2. 南京林业大学生物与环境学院,南京 210037)
花楸属(L.)隶属于蔷薇科(Rosaceae)、苹果亚科(Maloideae),该属植物有100~250 多种。花楸属是蔷薇科内分类问题最为复杂的属之一,从属下分类到物种界定均存在着不同观点。花楸属有广义(s.l.)和狭义(s.str.)之分,广义花楸属包含复叶和单叶种类,狭义花楸属包含除以外的复叶种类。我国分类学者多采用广义花楸属的概念,本文亦采用此概念。花楸属植物广布于北半球温带地区,部分种类可延伸到北半球高纬度地带,少数分布至热带亚洲,我国是该属植物的多样性分布中心,其种类在西南山区尤为丰富。花楸属植物春季白花满树,秋季果实累累,其果色丰富,呈红、黄、白、或杂色,不仅是高山风景区秋季重要的色彩组成,还是野生兽类和鸟类重要的食物来源。
叶片是众多高等植物进行同化作用的主要功能器官,其结构特征是物种进化和环境适应的综合表达,具有系统学和生态学意义。一方面,叶解剖结构特征可为物种鉴定和系统演化提供依据;另一方面,叶片受外部环境的影响较为显著,其解剖结构的可塑性变化,可以很好地反映植物在不同生态环境下的适应策略。花楸属植物叶片的外部形态特征在种间和种内均存在较大差异,是造成物种界定困难的重要原因之一。该属植物分布于我国寒温带至亚热带高山,从海拔50 m 至树木线以上,不同种分布的气候带和海拔差异较大。花楸属植物叶解剖结构不仅存在种间差异,还表现出一定的环境适应性变化。目前国内仅有文献提及川西亚高山米亚罗人工林下的陕甘花楸()、山西省西南部中条山区的水榆花楸()以及四川贡嘎山天然林下的灰叶花楸()、西康花楸()、多对花楸()、湖北花楸()6 种花楸属植物叶解剖特征的环境适应性状。另有栽培条件下的花楸树()、黄山花楸()、石灰花楸()面对光照和干旱胁迫时产生的叶结构特征适应变化。分布在同一地区花楸属植物叶解剖性状的种间差异和环境适应性研究鲜见报道。
本文以四川龙苍沟国家森林公园内7 种野生花楸属植物为研究对象,对其叶片和中脉的解剖结构特征、可塑性及其与环境因子的相关性进行比较分析,在此基础上探讨了叶解剖结构特征的分类学意义及其环境适应性状,以期更好地理解花楸属植物的种间差异及其环境响应策略,为本属植物的适应性进化研究提供基础资料,同时为本属植物的引种驯化提供参考。
1 材料和方法
1.1 研究区概况
四川雅安荥经县龙苍沟国家森林公园(102°0′~102°9′E,29°28′~29°57′N)地处四川盆地与青藏高原过渡带,面积达1 781 km,海拔处于690~3 666 m,为亚热带季风气候,全年四季分明,降水量充沛;植物资源极为丰富,是花楸属植物分布较为集中的区域之一。
1.2 试验材料
2020 年9 月于龙苍沟国家森林公园采集7 种花楸属植物:毛背花楸()、江南花楸()、大果花楸()、泡吹叶花楸()、多对西康花楸(var.)、晚绣花楸()和梯叶花楸()。每种植物选取3株成年单株(多对西康花楸仅采集到2 株长势良好的成年植株),每个单株采集5 片成熟、无虫害叶片立即浸于70%FAA固定液中备用,单株地理信息见表1。
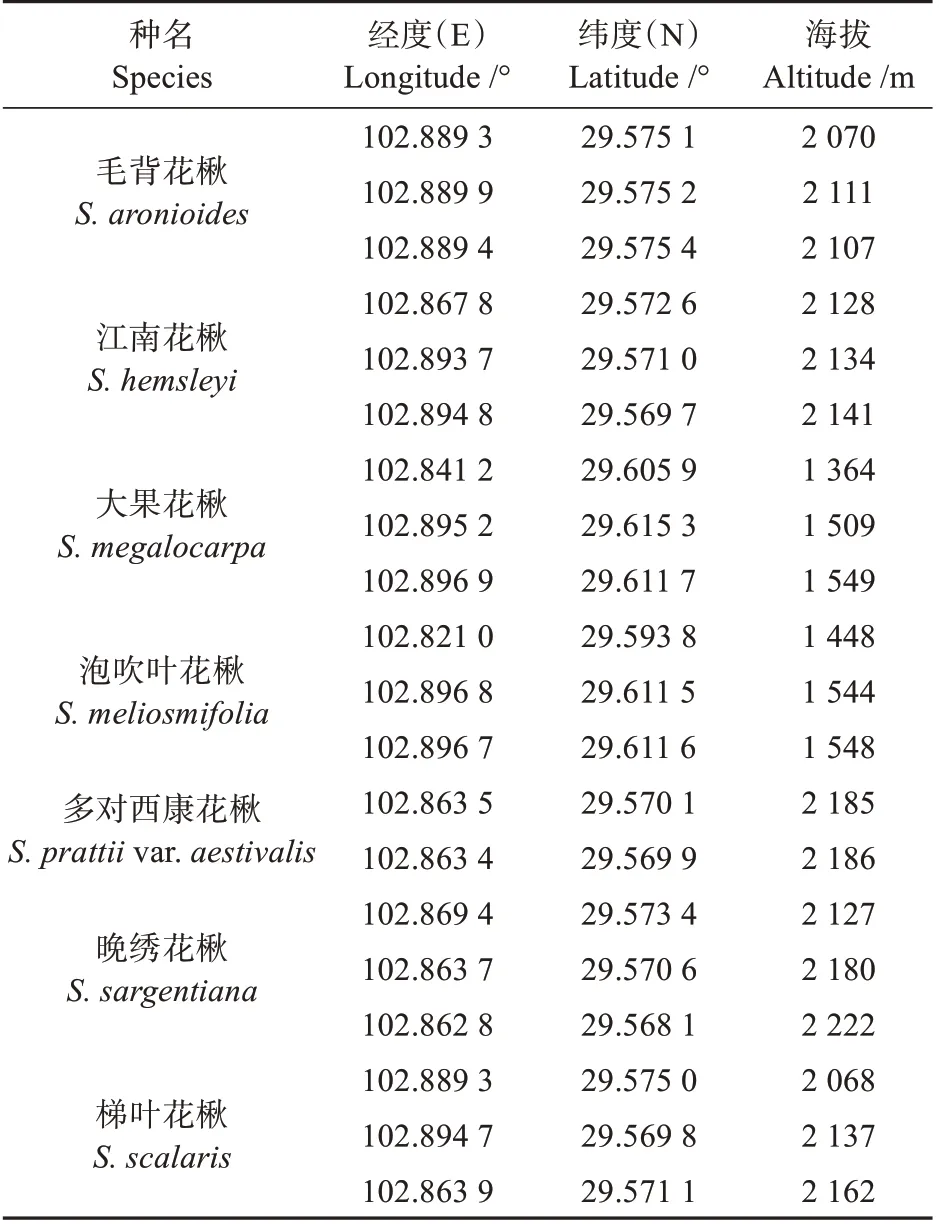
表1 样本来源Table 1 Sources of samples
1.3 试验方法与指标测定
采用常规石蜡切片法,将固定后的叶片沿主脉(距叶基部1/3 处)及叶肉部分分别裁剪成0.3 cm×0.5 cm 的样品,经梯度脱水后,浸蜡并包埋。使用Leica RM 2255自动切片机进行常规石蜡切片制作(切片厚度8µm),番红—固绿双重对染,中性树胶封片。制作完成的样片在Nikon Eclipse Ci-S 显微镜下进行观察,每个物种随机选取3~5 张切片的10 个视野,使用NIS-Element D 4.30 成像软件进行拍摄。使用Image J(https://imagej.net/)软件测量叶解剖结构指标,包括叶片厚度(LT,Leaf thick‐ness)、上表皮厚度(UET,Upper epidermis thick‐ness)、下表皮厚度(LET,Lower epidermis thick‐ness)、栅栏薄壁组织厚度(PT,Palisade parenchy‐ma thickness)、海绵薄壁组织厚度(ST,Spongy pa‐renchyma thickness)和中脉直径(DM,Diameter of midrib)。
根据测量数据计算以下指标:


1.4 环境数据来源与选取
气候数据共包含19 个生物气候变量,均来源于CHELSA(Climatologies at high resolution for the earth’s land surface areas,https://chelsa-climate.org/)气候数据集(1979~2013 年)。使用Arc-GIS 10.6(ESRI,Redland,USA)将数据集内的气候图层转换为GCS_WGS_1984 坐标系,然后利用空间分析工具提取所有样本采集点的气候变量数据。
为避免变量过多而产生多重干扰,将气候变量分为三大类(温度因子、降水因子和水热综合因子),同时使用Pearson 相关性分析与主成分分析(PCA)法对其进行降维处理。在高度相关(>0.9,<0.01)的同类气候因子中,选用主成分贡献值最大的因子作为影响叶解剖结构与种源地环境关系的主导因子(筛选结果见表2)。

表2 气候变量Table 2 Climatic variables
1.5 数据分析
使 用SPSS 26.0 软 件(IBM Corporation,Ar‐monk,New York,USA)对叶片解剖结构观测数据进行单因素方差分析(one-way ANOVA),以检验指标差异性,并使用Duncan 法进行组间多重比较。在R4.0.3(https://www.r-project.org/)软件中运行相关性分析与主成分分析;使用cor 和Hmisc 函数进行相关性分析的运算,以探究叶片各解剖特征之间以及与环境因子的关系;使用prcomp 函数对标准化的解剖结构参数值进行主成分分析的运算,并使用factoextra 程序包将分析数据可视化,以筛选种间差异最大的叶结构指标。
2 结果分析
2.1 花楸属植物叶解剖结构特征
7 种花楸属植物的叶均为典型的背腹叶(dor‐si-ventral leaf),由表皮(epidermis)、叶肉(meso‐phyll)和叶脉(vein)3 部分组成(见图1)。叶片厚度及各结构厚度、栅海比、组织结构紧密度、疏松度与中脉突起度种间差异极显著(见表3,<0.01)。叶片厚度介于108.16~208.21 µm,毛背花楸的叶片最厚,最薄的为大果花楸。
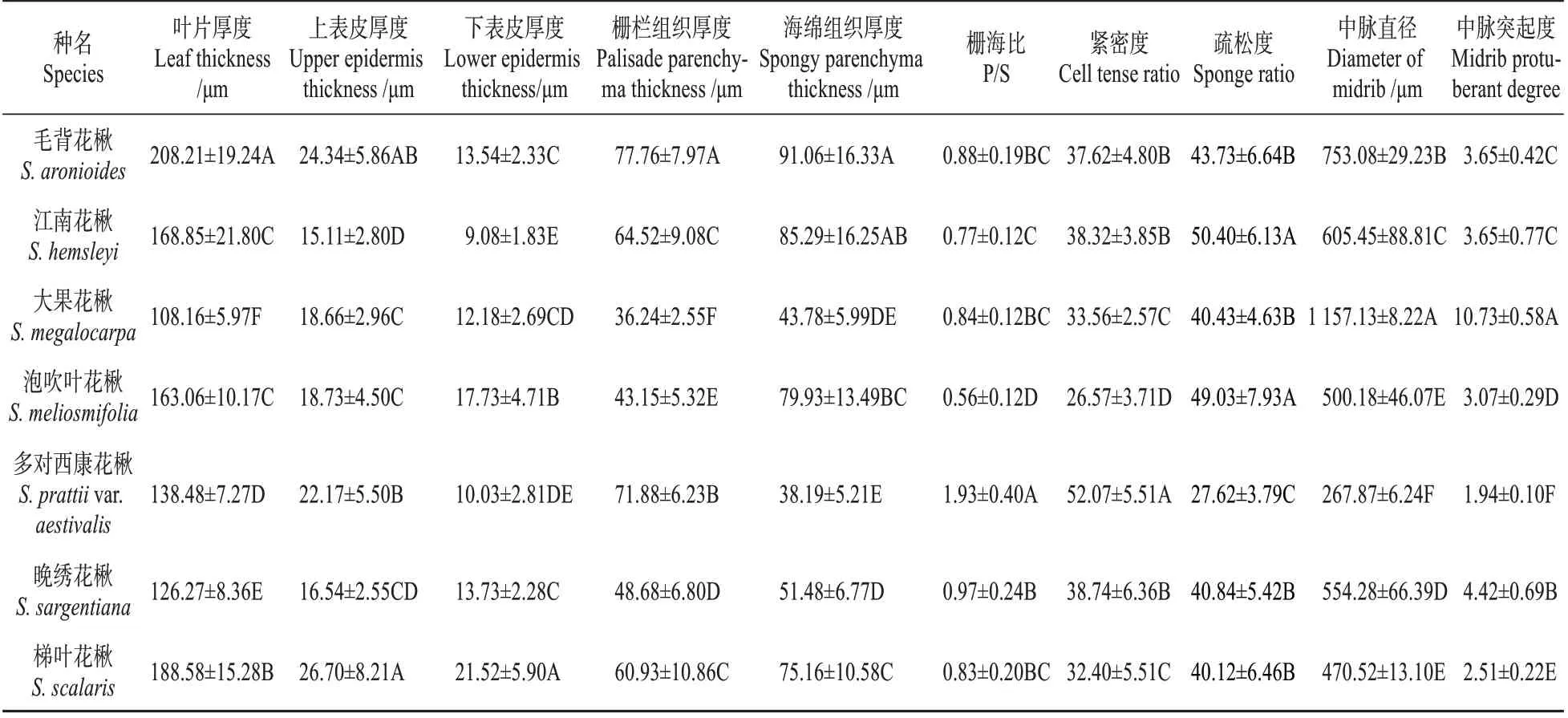
表3 花楸属植物的叶片解剖结构参数Table 3 Leaf anatomical parameters of Sorbus species
叶的上、下表皮均由排列紧密的单层细胞构成,上表皮厚度介于15.11~26.70µm,下表皮厚度介于9.08~21.52 µm,上表皮厚度均大于下表皮;梯叶花楸的上、下表皮均为最厚,而江南花楸的表皮厚度均最薄;多对西康花楸(见图1:E1)、晚绣花楸(见图1:F1)和梯叶花楸(见图1:G1)3种复叶类群的下表皮细胞上存在明显的乳突结构。
叶肉分化成栅栏薄壁组织和海绵薄壁组织,栅栏组织邻接在上表皮内侧,由1~2 层长柱状薄壁细胞整齐排列而成;其中毛背花楸、多对西康花楸与梯叶花楸具有明显的2 层栅栏组织(见图1:A1、E1、G1),其余4 种均只含有1 层栅栏组织。多对西康花楸具有最发达的栅栏组织,栅海比为1.93,其叶组织结构紧密度也最大(52.07%);海绵组织邻接在下表皮内侧,由不规则的薄壁细胞构成,具有显著的细胞间隙;泡吹叶花楸具有最发达的海绵组织,细胞间隙较大(见图1:D1),栅海比仅0.56,组织结构疏松度为49.03%。
叶片主脉发达,维管束呈心形,木质部、韧皮部均具有明显的分化,内外相接,是典型的外韧型维管束。维管束周围排列着4~8 层薄壁组织和厚角组织,其中分布着染色较深的异细胞。中脉直径介于267.87~1 157.13 µm,中脉突起度介于1.94~10.73,大果花楸的中脉最为发达且突起度最大(见图1:C2),相对于中脉最不发达且突起度最小的多对西康花楸(见图1:E2),其直径为后者的4.32倍,其中脉突起度为后者的5.53倍。
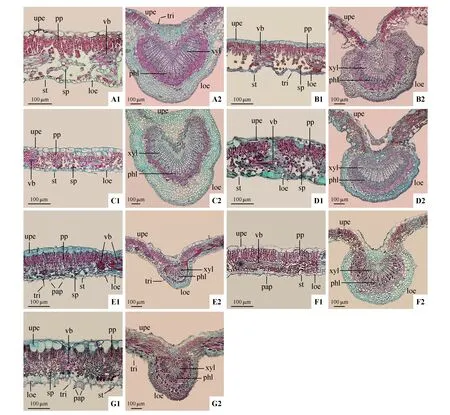
图1 花楸属植物叶片解剖结构(叶肉×20,中脉×10)A1~A2.毛背花楸;B1~B2.江南花楸;C1~C2.大果花楸;D1~D2.泡吹叶花楸;E1~E2.多对西康花楸;F1~F2.晚绣花楸;G1~G2.梯叶花楸;upe.上表皮;pp.栅栏组织;sp.海绵组织;loe.下表皮;st.气孔;tri.毛状体;pap.乳突;vb.维管束;xyl.木质部;phl.韧皮部Fig.1 Leaf anatomical structure of Sorbus spp(.mesophyll×20,midrib×10)A1-A2.S.aronioides;B1-B2.S.hemsleyi;C1-C2.S.megalocarpa;D1-D2.S.meliosmifolia;E1-E2.S.prattii var.aestivalis;F1-F2.S.sargentia⁃na;G1-G2.S.scalaris;upe.Upper epidermis;pp.Palisade parenchyma;sp.Spongy parenchyma;loe.Lower epidermis;st.Stomates;tri.Trichomes;pap.Papilla;vb.Vascular bundle;xyl.Xylem;phl.Phloem;
2.2 叶解剖结构变异系数与可塑性指数
叶解剖结构特征的变异系数与可塑性指数表明(见表4),叶片综合变异均值在10.41%(大果花楸)~16.73%(梯叶花楸),可塑性均值在0.31(大果花楸)~0.46(江南花楸、梯叶花楸),大果花楸叶解剖结构整体可塑性较小,梯叶花楸与江南花楸整体可塑性较大。
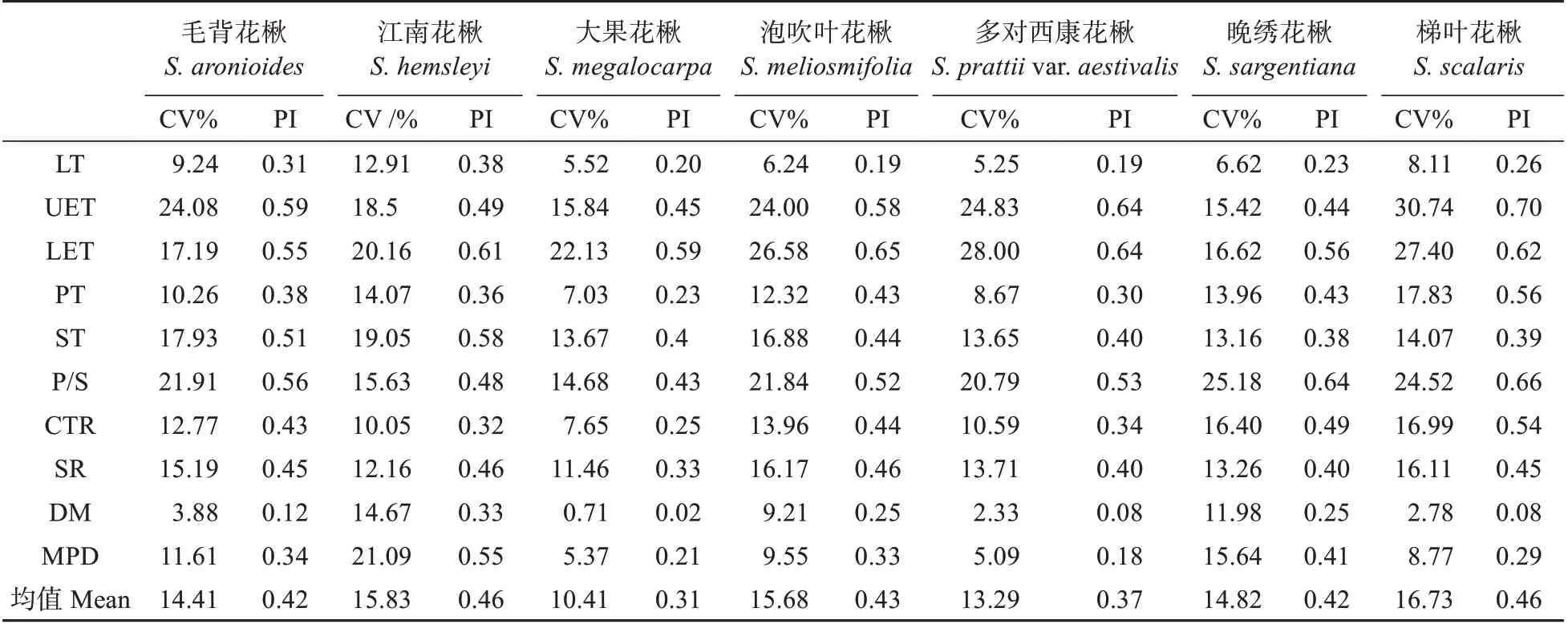
表4 花楸属植物叶解剖结构的变异系数和可塑性指数Table 4 Coefficients of variations and plasticity indexes of leaf anatomical structures of Sorbus spp.
叶各解剖结构性状之间的变异系数与可塑性指数差异较大,上、下表皮厚度的变异程度最大(15.42%~30.74%)、可塑性最强(PI=0.45~0.70),但种间区别较小;作为主要输导结构和支撑结构的主脉,其变异系数为0.71%~14.67%,可塑性指数介于0.02~0.33,整体较低,但种间区别最大。
2.3 叶解剖结构的自相关性和主成分分析
相关性分析结果显示(见表5),各结构参数间存在着一定的自相关性。叶片厚度与上表皮厚度、栅栏组织厚度和海绵组织厚度呈极显著正相关性(<0.01);与叶片厚度相关性最大的海绵组织厚度(=0.858),是影响叶片厚度的重要因素。上、下表皮厚度之间存在正相关性(<0.05),此外,上表皮厚度还与栅栏组织厚度存在正相关(<0.05)。在叶肉组织厚度及组织比例之间,栅栏组织厚度与紧密度存在正相关(<0.01);海绵组织厚度与疏松度存在正相关(<0.01),与栅海比呈显著负相关(<0.01)。在中脉结构指标与各结构指标之间,中脉突起度仅与中脉直径正相关(<0.01),而与叶片厚度、栅栏组织厚度和海绵组织厚度均呈负相关(<0.05)。
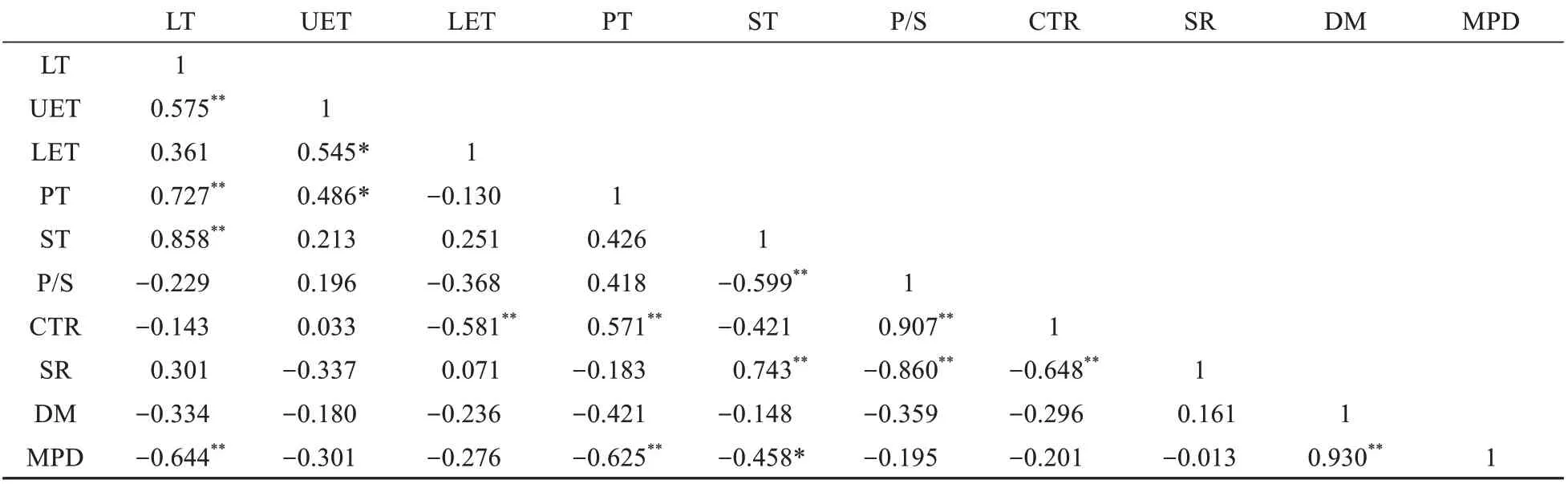
表5 叶解剖结构的相关性分析Table 5 Correlation analysis of anatomical structures of leaves
主成分分析结果显示(见表6),前3 个主成分的累计贡献率达到86.87%,表明这3 个主成分足以表达10 个解剖结构指标的主要信息。其中,第一主成分占36.83%,载荷量最高的为叶片厚度和海绵组织厚度,主要反映叶片总厚度的组成特点;第二主成分占比35.42%,载荷量最高的为栅海比和紧密度,反映了叶肉组织的分布情况;第三主成分占比14.62%,载荷量最高的为上、下表皮厚度,反映的是表皮组织的特点。变量坐标图(见图2)显示了各叶解剖结构所占权重,其中,栅海比是主成分中贡献值最高的变量(0.991),其次为组织结构紧密度(0.933)、海绵组织厚度(0.931)、中脉突起度(0.890)和栅栏组织厚度(0.880)。
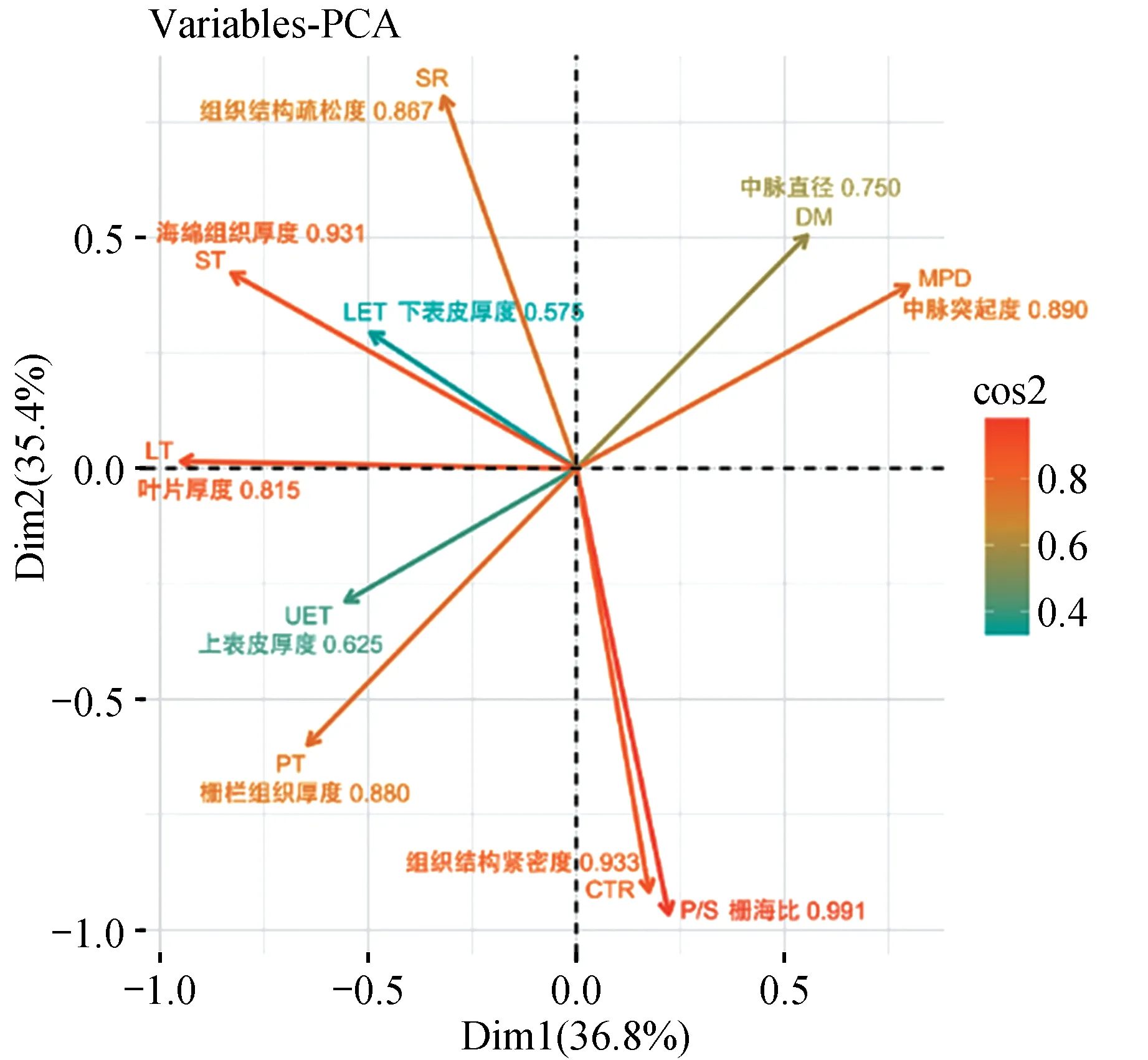
图2 PCA分析生成的叶解剖结构变量坐标图Fig.2 Coordinates map of anatomical structures of leaves generated by PCA analysis

表6 叶解剖结构指标在3个主成分中的载荷系数Table 6 Loading factors of leaf anatomical traits in three principal components
2.4 叶解剖结构与环境因子的相关性分析
叶解剖结构参数与环境因子的相关性显示(见表7),受环境影响最大的是与栅栏组织、海绵组织和中脉密切相关的结构参数。气候因子中,季节性温差与中脉直径、中脉突起度呈显著正相关(<0.01),与栅栏组织厚度、栅海比和紧密度呈负相关(<0.05);年降水量和最暖季降水均与栅栏组织厚度、栅海比和紧密度3个指标存在正相关(<0.05),与中脉直径、中脉突起度存在显著负相关(<0.01);地理因子中,海拔与栅栏组织厚度和紧密度呈现显著正相关(<0.01),而与中脉直径及其突起度呈负相关(<0.05)。
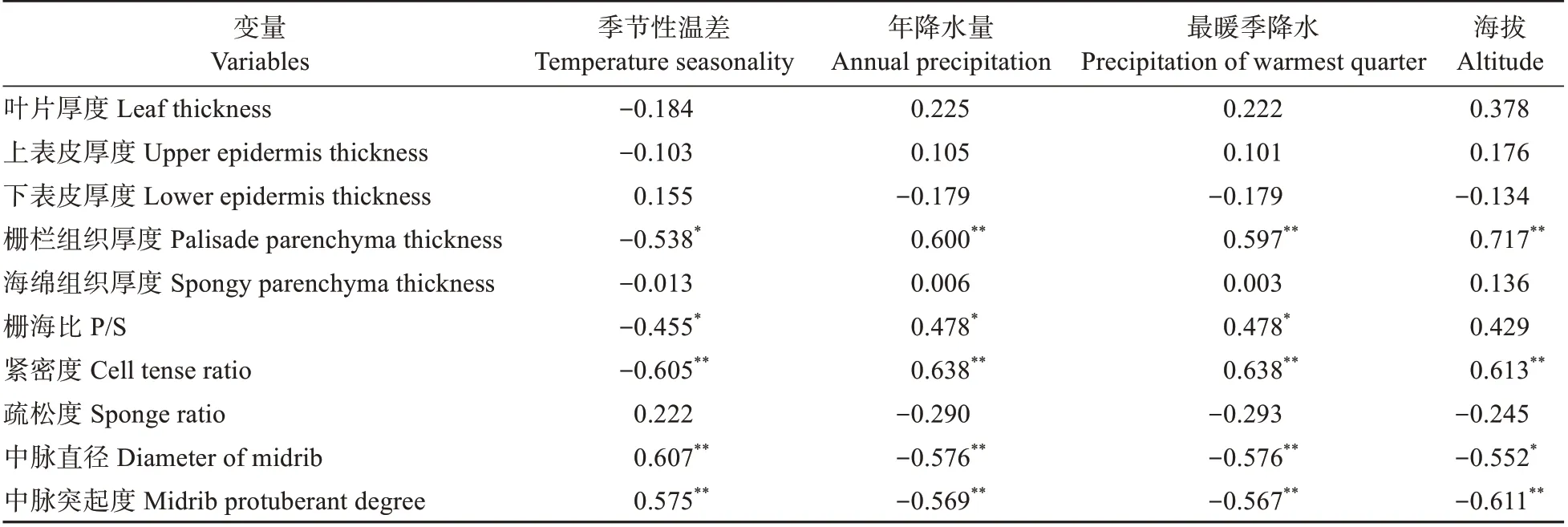
表7 叶片解剖结构指标与环境因子的相关性Table 7 Correlation coefficient between leaf anatomical traits and environmental factors
3 讨论
本研究中所取的花楸属植物均具有典型的背腹叶,从结构上看,叶片厚度的变化主要是由海绵组织厚度变化而引起,叶片上表皮厚度均大于下表皮厚度,与陕甘花楸、水榆花楸、黄山花楸、石灰花楸、欧亚花楸()和棠楸()等花楸属植物的叶特征相同。7 种植物在栅海比、紧密度、海绵组织厚度、中脉突起度和栅栏组织厚度上显示出最大的差异性,因此,栅栏组织、海绵组织和中脉组织是7种花楸属植物中种间差异最显著的解剖结构,可以为将来探讨属下亲缘关系提供解剖学分类依据。此外,仅3种复叶类群(多对西康花楸、晚绣花楸和梯叶花楸)的下表皮具有乳突结构,与郗连连对花楸属复叶组植物叶微形态特征的研究结果一致,单、复叶类群在该结构上的区别对花楸属的系统分类及演化具有重要意义。
在叶解剖结构中,变异系数大于50%的性状被认为是生态适应性状,变异系数较小的被认为是相对稳定的系统演化性状,反映的是物种潜在适应能力。可塑性指数反映了物种抵御环境压力的能力,较高的可塑性水平意味着较强的系统调节能力。本研究中花楸属植物叶结构变异系数较低,各结构相对稳定,可以被认为是演化性状。其中,变异性和可塑性最大的是表皮组织厚度,表皮组织的变化可以增强细胞对光的捕获能力,有利于光辐射穿透叶表皮到达叶肉组织,提高光合效益。龙苍沟国家森林公园分布的花楸属植物多生长于高海拔地区,气温较低且风速较大,影响了植物叶片的液流速度,上表皮较下表皮更厚有利于气体交换,充分加强了叶的碳吸收能力。表皮组织的这些适应性变化是龙苍沟地区花楸属植物对高海拔环境和林下不同光条件的共同响应。变异性和可塑性最小的是中脉直径,中脉是叶片进行水分吸收与贮藏的主要结构,其直径的稳定性较大可能与当地空气湿度大、土壤含水量高等良好的水分条件有关。
叶解剖结构综合变异系数的种间差异也在一定程度上反映了物种的环境适应能力。综合变异系数和可塑性指数均为最高的梯叶花楸,在龙苍沟国家森林公园内分布的海拔梯度最大(1 300~2 300 m),而2 项指标值均最低的大果花楸海拔分布范围最小(1 300~1 500 m)。由此可见,变异性和可塑性较高的植物具有较强的潜在适应能力和克服环境异质性的能力,也往往分布得更为广泛。
种间差异最大的栅栏组织、海绵组织和中脉组织受气候与海拔因子的影响也最大,因此,叶解剖结构性状的差异不仅与自身的生长特性有关,也与环境因子密不可分。当季节性温差变大时,7 种花楸属植物通过降低栅栏组织厚度、提升叶脉突起度来增加叶脉能量分配的方式来适应温差变化、抵御寒冷气候条件,进一步反映了叶脉与叶片各输导组织和同化组织之间相互协调、相互制约的关系。年降水量、最暖季降水的增加以及海拔梯度的升高,导致了水热综合条件的改变,高海拔山区强光照、低温、低CO浓度等环境条件影响了植物的蒸腾作用,导致体内水分及无机盐的运输速率降低,进而影响植物的生长发育。栅栏组织的增厚引起了叶片紧密度的提升,是7种花楸属植物为适应湿润环境而产生的结构变化,与石灰花楸和花楸树的适应策略一致。同时,这一特征变化还提高了植物在高海拔地区的抗寒性及光合效能,花楸属植物叶解剖结构对海拔的适应性变化与刘梦颖等的研究结果一致。
本研究中,7种植物对四川龙苍沟国家森林公园环境产生的适应性变化,与川西地区(贡嘎山和米亚罗林区)其他花楸属植物的生存策略一致。同质环境下,叶片各解剖结构间的协同变化,是该属植物面对环境压力时提高自身生存适应性、协调与生境的合作关系的有效方式。7种花楸属植物的叶解剖特征不仅表现出了显著的种间差异,还很好地诠释了花楸属植物对高海拔环境的响应,对阐释花楸属植物在叶解剖性状上的进化关系及其环境适应性具有重要意义。
猜你喜欢
祝您健康·文摘版(2019年4期)2019-06-11
法治研究(2018年2期)2018-09-10
当代教育(2018年4期)2018-01-23
东方教育(2016年4期)2016-12-14
课程教育研究·学法教法研究(2016年21期)2016-10-20
饮食科学(2016年3期)2016-07-04
小学教学参考(语文)(2016年4期)2016-05-07
第二课堂(课外活动版)(2015年6期)2015-10-21
建筑遗产(2014年1期)2014-10-21
第二课堂(小学版)(2014年4期)2014-08-02
