
松褐天牛繁殖行为及其影响因素研究进展
2022-03-03 09:10郑光楠杨秀好董子舒韦曼丽郑霞林
广西林业科学 2022年1期
郑光楠,杨秀好,董子舒,韦曼丽,郑霞林
(1.广西大学 农学院,广西南宁 530004;2.广西壮族自治区林业科学研究院,广西南宁 530002;3.广西壮族自治区林业有害生物防治检疫站,广西南宁 530028)
松褐天牛(Monochamus alternatus)属鞘翅目(Coleoptera)天牛科(Cerambycidae)墨天牛属,主要危害马尾松(Pinus massoniana)、黑松(P. thunber⁃gii)、雪松(Cedrus deodara)和火炬松(P.taeda)等树种。该虫在中国分布于除黑龙江、内蒙古、北京、宁夏、甘肃、青海和澳门外所有省(市、自治区)[1];在国外分布于日本、朝鲜、韩国、老挝、越南、加拿大、美国、墨西哥、希腊和葡萄牙等国家[1-2]。在中国,该虫1年发生1 ~3 代,以老熟幼虫在树干中越冬[3-5];室内人工饲养时发现,取食马尾松的雌雄成虫平均寿命分别为(63.13 ± 12.33)天和(59.04 ± 12.18)天[6]。该虫还是检疫性有害生物松材线虫(Bursaphelen⁃chus xylophilus)的传播媒介,由其传播的松材线虫病给中国林业产业发展造成了重大经济损失。2017年,中国大陆松材线虫病总成灾面积达85 524 hm2,造成经济损失约195 亿元[7]。鉴于松褐天牛是松材线虫病主要传播媒介,国内外学者对松褐天牛开展了大量研究,包括生物学[8]、生态学[9-10]、行为学[11-14]、生理学[15]、分子生物学[16]和综合防治[17]等。
繁殖是物种延续的前提。研究害虫的繁殖行为,针对其行为采取相应的防治手段、制定针对性的防治策略,可有效控制害虫种群。目前,关于松褐天牛繁殖行为方面的报道包括异性定位[18-19]、交配行为和节律[20-21]及产卵行为[22]等。部分研究结果已成功应用于生产实践,且有效降低了松褐天牛种群数量。樊建庭等[23]研究明确了松褐天牛的聚集性信息素和对其有较强吸引作用的植物源信息素,这两种物质组分复配的引诱剂对该虫有较强的引诱效果;根据松褐天牛成虫对初伐木有较强的趋性这一特性,在林间设置初伐木为诱木,引诱松褐天牛在其上产卵,然后集中销毁诱木,可有效降低该虫的子代种群数量[24]。为系统了解松褐天牛繁殖相关研究,该文对松褐天牛繁殖行为及其影响因素进行综述,旨在为综合利用其繁殖行为开展松褐天牛监测和防控技术研究提供参考。
1 松褐天牛异性定位
松褐天牛首先依靠嗅觉和视觉器官定位受害松树[25],尤其偏好衰弱木和初伐木[19]。与健康松树相比,受害松树会释放更多的α-蒎烯、β-蒎烯和D-柠檬烯,还会释放樟脑烯、β-苯丙氨酸烯、可可烯、长叶烯和β-石竹烯等挥发物,这些挥发物被松褐天牛触角上的毛形感器、锥形感器和耳形感器接收后,通过感器壁上的小孔进入感器淋巴液[26],再通过CSP5、OBP1、OBP9 和OBP10 等气味结合蛋白和化学结合蛋白对这些挥发物进行特异性结合,使松褐天牛成虫能从远距离进行聚集[27-30]。除嗅觉外,松褐天牛也可通过视觉识别衰弱木和濒死松木的颜色[31],进行更进一步的精准定位。当雄虫在受害松树聚集后,其产生的长距离信息素(2-十一烷氧-1-乙醇)与植物挥发物共同作用吸引雌虫聚集[32]。
当雌雄成虫聚集后,雄虫可通过嗅觉和视觉发现雌虫。雌虫产生的短距离信息素和接触信息素有助于雄虫识别雌虫,完成异性定位[18]。研究发现,当雄虫嗅觉和视觉缺失时,抱对成功率显著降低,视觉缺失处理下的抱对成功率显著低于嗅觉缺失处理,可能是因为松褐天牛成虫近距离接触时更依赖视觉定位异性[33-34]。松褐天牛是否存在接触信息素,目前仍存在分歧[35-36]。研究还发现在视觉和嗅觉均缺失的情况下,仍有松褐天牛能成功抱对[33],可能还存在其他尚未发现的异性定位途径。研究发现,部分天牛种类成虫可利用异性发出的声音觅偶[37-38]。松褐天牛成虫是否可利用听觉或其他途径进行异性定位,尚需进一步研究。
2 松褐天牛交配行为及节律
2.1 松褐天牛交配行为
松褐天牛成虫羽化出木后,多取食1 ~3年生的松枝韧皮部补充营养,尤其喜食马尾松当年生嫩枝[39-40];补充营养约10天后,进入交配期。松褐天牛有较长的交配前期,主要是因为成虫羽化后需补充营养至性成熟才能进行交配[41]。松褐天牛成虫羽化出木后15 ~20天为交配高峰期[42]。
松褐天牛成虫完整的交配行为包括相遇抱对、插入输精和配后保护3 个阶段(图1)。雄虫发现雌虫后,会在雌虫附近停留十几分钟,然后突然爬向雌虫抱对交配[43];也有研究发现,部分性成熟雌虫会主动靠近雄虫,寻求交配[44]。当雌雄成虫接触后,雄虫会用前足抱握雌虫,调整身体爬到雌虫体背,用前足与中足抱握雌虫,用上颚咬住雌虫鞘翅以形成能交配姿势,有时雄虫在阻止雌虫反抗的过程中会咬掉其鞘翅[43],出现咬掉雌虫鞘翅这种情况是雄虫的一种交配策略还是其未掌握好力度尚不明确。1对成虫正在交配时,有时会有第2头雄虫出现,与正在交配的雄虫打斗,直至一方落败逃跑[12]。如果第2 头雄虫胜利,则其与该雌虫发生交配;反之,原始配对的雄虫继续与雌虫进行交配。也有研究发现,部分原始配对的雄虫即使“胜利”,也会离开,不继续与雌虫交配[43]。在抱对过程中,有时会出现雌虫强烈反抗和逃离使雄虫不能与之交配的情况;有时会发生错误识别,出现雄虫和雄虫、雌虫和雌虫同性抱对并尝试交配的现象,雄虫同性抱对比例比雌虫高[11];有时甚至会出现与云杉花墨天牛(M.saltu⁃arius)尝试交配的情况[45]。黑腹果蝇(Drosophila me⁃lanogaster)也有同性抱对行为,有过同性抱对行为的雄性在与雌性交配时成功率显著提高[46],推测雄虫在同性性行为中学到了更多求偶经验,提高了其与雌虫的交配成功率[47]。松褐天牛同性抱对及与同属其他昆虫抱对是因错误识别还是为后续能成功交配而开展的练习,亦或是其他原因,尚不清楚。
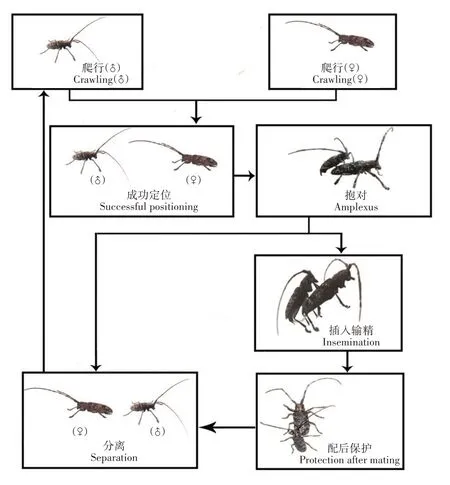
图1 松褐天牛觅偶、求偶和交配行为Fig.1 Behavior of mate location,courtship and mating of M.alternatus
成功抱对后,雄虫腹部末端向下弯曲,外生殖器插入雌虫体内,拉出输精管开始输精。观察发现,松褐天牛成虫在抱对过程中,雌虫可爬动与取食;当雌虫爬动而雄虫又想交配时,雄虫会用颚刮雌虫的鞘翅使其静止,然后将外生殖器插入雌虫阴道完成输精[43]。在输精过程中,雄虫外生殖器由白色透明逐渐变黄再变为橙黄色[43]。在松褐天牛成虫完整交配过程中,雄虫有1 次或多次将外生殖器插入雌虫体内的行为,每次插入时间为3 ~251 s,平均为(57.60±1.363)s[20]。松褐天牛的输精行为有别于其他天牛科昆虫的另一种交配方式“插而不拔”[48],推测松褐天牛多次插入的输精方式有助于其快速输精、躲避同性竞争等风险。
雄虫生殖器从雌虫体内最后一次拔出至结束抱对期间为配后保护。在配后保护期间,雄虫一直用前足抱握雌虫的后胸,阻止其他雄虫与该雌虫交配,时长为0.50 ~115.68 min,平均(15.18 ±2.145)min[20]。
2.2 松褐天牛交配节律
松褐天牛在00:00 ~24:00 均有交配行为发生,交配高峰期为18:00 ~24:00[21,43-44](表1)。有研究发现,松褐天牛在10:00 ~14:00 也有一个交配高峰期[44]。松褐天牛交配高峰期存在差异的原因可能是地理环境因素。

表1 松褐天牛昼夜交配节律Tab.1 Diurnal rhythm of mating of M.alternatus
3 松褐天牛产卵行为
松褐天牛出木后约16 天即可产卵[49],卵多产于树干阳面[50]。开始产卵前,松褐天牛雌虫用下颚须、下唇须和触角探触松树树皮表面,寻找合适产卵的位置。在寻找到合适位置后,雌虫用上颚在树皮上咬出1个眼状产卵刻槽,继而旋转身体180°,将产卵器插入刻槽内[43,51],其产卵器受到松树内皮黄烷醇葡糖苷和原花青素等挥发性物质刺激,将卵产在刻槽内[52-53]。雌虫每次产卵1 ~3 粒,以1 粒居多;产卵后,并不立即从刻槽内拔出产卵器,而是先用产卵器分泌胶状物质于刻槽中,再慢慢拔出产卵器[43,51]。雌虫在刻槽内不产卵会形成空刻槽(约占刻槽总数的40% ~50%)[54]。推测松褐天牛雌虫制作空刻槽的原因是既可进一步衰弱树势,有助于子代取食,又能减少其子代被天敌发现的机率,提高种群存活率。雌虫咬出1 个刻槽需10 ~15 min,产1 粒卵需5 ~9 min[43,54]。根据卵在刻槽中的位置可分为纵产、横产和斜产3种[55]。
研究发现,松褐天牛交配后1 ~3 天,雌虫仅咬刻槽并不产卵,使刻槽中的松脂流出[56],减弱松木的防御能力,交配后4 ~6 天开始产卵[51,54]。与体型较小雌虫相比,体型较大雌虫产卵前期更短,且产卵量更多,但所产卵粒较小[8]。雌虫多在夜间产卵,平均每日产卵3 ~6 粒,产卵期约60 天,单雌产卵量40 ~180粒[54]。
4 影响松褐天牛繁殖行为的因素
4.1 非生物因素
4.1.1 温度
松褐天牛最适繁殖温度为25 ~30 ℃[39]。温度偏低时,成虫抱对率和交配率均降低;温度下降至18 ℃时,成虫潜伏不动。低温还会影响雌虫卵巢发育。夜间温度过低(低于15 ℃)时,雌虫卵巢发育减缓且大部分卵不能正常发育[57];在年均气温低于8.2 ℃的地区,松褐天牛不能成功繁殖后代[58]。外界环境为高温低湿时,松褐天牛雌虫咬出的刻槽数量多且有效刻槽所占比例更高;低温高湿时,雌虫咬出的刻槽数量和产卵量均下降[59]。
4.1.2 降雨和气流
降雨和气流会使植物挥发物浓度降低或发生定向转移,影响昆虫对化学信息物质的接收[60]。在松褐天牛中也发现类似现象。降雨时,松褐天牛的成虫交配行为减少,雌虫产卵量明显下降,可能是降雨影响信息素传递和成虫自身活动[61]。
4.1.3 辐射
使用40 GY 及以上剂量的60Coγ 射线辐射松褐天牛雌雄成虫,虽然雌雄成虫仍能正常交配,但与健康异性交配后所产的卵均不能正常孵化;随辐射剂量升高,成虫寿命缩短[62-63]。进一步研究发现,射线处理后,松褐天牛体内糖酵解过程效率降低,该途径中底物(葡萄糖和半乳糖)浓度和某些中间产物(苹果酸等)浓度升高,推测辐射破坏了该途径中某些基因,从而无法正常繁殖后代[63]。
4.1.4 杀虫剂
鱼藤酮影响松褐天牛呼吸,且对松褐天牛幼虫有致畸作用[64]。试验表明,在衰弱木或伐倒木上喷刷鱼藤酮制剂,可有效减少松褐天牛在其上产卵[64]。
4.2 生物因素
4.2.1 自身因素
个体大小影响松褐天牛交配。大体型雄虫与小体型雄虫相比,更容易成功定位异性;大体型雄虫与小体型雌虫交配时,常难以成功交配[43]。可能是因为体型差异较大导致雄虫在交配时需要进行的曲腹动作更大,投入的精力更多;或者是因为雄虫在插入雌虫体内时,会带给雌虫超过忍受限度的疼痛感,使得雌虫剧烈反抗,阻止交配的发生[43-44]。
性比会影响松褐天牛繁殖。雄雌比例为2∶1时,雌虫的平均产卵量为107.2 粒;雄雌比例为1∶1时,雌虫的平均产卵量为153.6 粒;前者有效刻槽数低于后者,前者空刻槽数高于后者[65]。
不同交配次数影响雌虫繁殖力和寿命。未交配雌虫寿命显著高于已交配雌虫;只交配1 次雌虫寿命与交配多次雌虫寿命差异不显著[20,66]。雄虫向雌虫体内输入精子和精液等物质后,雌虫会将更多的资源用于繁殖,寿命缩短。在其他昆虫中也发现此类现象,如棉红铃虫(Pectinophom gossypiella)[67]、褛裳夜蛾(Catocala remissa)[68]和茶尺蠖(Ectropis obliqua)[69]。松褐天牛雌雄成虫均存在非专一性的多次交配现象[42,44];交配次数不同,雌虫繁殖力有一定差异。研究发现,多次交配雌虫受精囊内的精子数量一直稳定在12 万个左右,仅交配1 次雌虫受精囊内的精子数量随时间推移不断减少;交配后前3周,仅交配1 次雌虫的产卵量和卵孵化率与多次交配雌虫差异不显著;从第4 周开始,仅交配1 次雌虫的产卵量和卵孵化率急剧下降,多次交配雌虫的产卵量和卵孵化率无显著变化;总体来看,多次交配雌虫的总产卵量(167.087 0 ± 9.187 7)和卵孵化率(94.4 ± 2.99)%均高于仅交配1 次雌虫(113.521 7 ±8.046 8、(83.79±14.53)%)[70]。
4.2.2 寄主植物
不同寄主植物影响松褐天牛产卵量。取食马尾松的松褐天牛雌虫产卵量显著高于取食湿地松(P.elliottii)和雪松的松褐天牛雌虫[6]。同种寄主植物不同年龄枝条影响松褐天牛产卵量。取食马尾松当年生枝条的雌虫日均产卵量显著高于取食2 和3年生枝条的雌虫,且前者寿命更长[40,49]。不同健康状态的寄主植物影响松褐天牛雌虫产卵选择。通常情况下,松褐天牛雌虫优先在衰弱木和伐倒木上产卵,其次为感染松材线虫濒死疫木和枯立木,最后为健康木。松褐天牛雌虫在健康木上咬出的空刻槽所占比例显著高于其在衰弱木、感染松材线虫濒死疫木和枯立木上所占比例[51];雌虫在健康木上的产卵量低且卵不能正常孵化,在伐倒木上的产卵量高且卵孵化率高[71]。相较于未感染松材线虫病的松树,雌虫更喜欢在感染了松材线虫病的松树上产卵[13]。寄主植物直径影响松褐天牛雌虫产卵选择。室内研究发现,松褐天牛雌虫在直径>3 cm 寄主植物上咬出的有效刻槽数及含2 粒卵的比例均显著高于直径<2 cm的寄主植物[55]。野外调查也发现,雌虫主要选择在直径5 ~14 cm 的寄主植物上产卵。可能是寄主植物直径过小时,不能满足幼虫生长发育所需的空间;而直径过大时,树皮太厚,会影响雌虫制作产卵刻槽。
松褐天牛雌虫在寄主植物上形成的有卵刻槽、产卵分泌物和幼虫虫粪均会影响其产卵选择。松褐天牛雌虫对有产卵分泌物和幼虫虫粪的寄主植物有较强的产卵趋避性[14,72-73]。观察发现,当松褐天牛雌虫遇到未含有产卵分泌物的刻槽时,大多会在此产卵;当刻槽中含有产卵分泌物时,雌虫会在此刻槽附近形成新的刻槽但不产卵[15]。产卵分泌物对松褐天牛产卵场所选择的影响机制尚不明确。松褐天牛幼虫虫粪影响雌虫对产卵场所的选择,可能与其含有的特殊化合物有关,幼虫虫粪中的单萜和丁基化羟基甲苯混合物对雌虫产卵有抑制作用[73]。
4.2.3 松材线虫
松树感染松材线虫病后,其生长衰弱或死亡,吸引松褐天牛雌虫产卵,在树体中生长的松褐天牛又会携带松材线虫进行传播,形成松材线虫-松褐天牛-松树3 者间的循环[74]。松材线虫影响松褐天牛繁殖。携带有松材线虫的松褐天牛雄虫在交配时会将松材线虫一同输入雌虫受精囊内,使受精囊储存的精子数减少,影响卵的受精[75]。
4.2.4 微生物
松材线虫在进入松树体内时,其伴生真菌或细菌会同时侵入,并与松材线虫共同危害松树。不同种类的伴生真菌影响松褐天牛体内携带松材线虫的数量[76-77]。树体内菌为小孢拟盘多毛孢(Pestaloti⁃opsis microspora)M32 时,松褐天牛体内携带的松材线虫多于树体内菌为松球壳孢(Sphaeropsis sapinea)E11和松球壳孢MHS7.3时,间接影响松褐天牛的精子传输[78]。球孢白僵菌(Beauveria bassiana)也会影响松褐天牛产卵。当松褐天牛感染球孢白僵菌后,雌雄成虫的寿命均显著降低,雌虫日产卵量增加,总产卵量无显著变化[79]。
5 展望
松褐天牛因其传播松材线虫而严重威胁我国乃至整个东亚地区的松林安全。其作为传播松材线虫的媒介昆虫,可在自身补充营养时传播,还可通过繁殖行为传播。两性交配时,雄性可向雌性体内输入松材线虫[45,75],雌性产卵时将松材线虫进一步传播至新的松树上[80]。因此,弄清松褐天牛繁殖与松材线虫传播间的关系和机理,能更有效地控制其种群繁殖,是保护松林和减少林业损失的关键突破口。
研究昆虫繁殖行为有利于害虫种群控制。目前,松褐天牛繁殖行为的一些研究结果已应用于松褐天牛种群动态预测及防控。研发的引诱剂是防治松褐天牛的关键技术之一[23]。目前,投入使用较多的APF-I型引诱剂由性信息素和植物挥发物复配而成[81],在林间有较好的诱捕效果[82]。林间存在伐倒木或衰弱木会显著降低松褐天牛引诱剂诱集效果[9]。针对这一现象,应深入研究规避技术,开发更高效的引诱剂。非寄主植物挥发物对松褐天牛产卵有趋避作用。从非寄主植物中提取出的苯甲醛、香茅醛、薄荷香精油、桉树油、橙花醇、尼古丁和水杨醛均对松褐天牛雌虫产卵有趋避作用,其中尼古丁的趋避作用最显著[83]。在林间,将非寄主植物挥发物与现有引诱剂结合使用,采取“趋避-吸引”策略,是否能更好地控制松褐天牛种群值得深入探索;也可考虑在松林中种植对松褐天牛产卵趋避性较强的植物,降低松褐天牛种群密度。
虽然松褐天牛繁殖方面已有一些研究和报道,仍有很多问题值得深入研究。研究松褐天牛成虫是否利用听觉觅偶,有助于利用声学技术干扰该虫的觅偶和交配[84-85],达到控制其种群的目的。松褐天牛异性定位、两性选择、“同性抱对”的机理及松褐天牛携带松材线虫对其繁殖影响的机制等尚不明确。可通过行为学、分子生物学和化学生态学等学科的技术和手段,对其进行深入研究,不仅有助于明确松褐天牛的繁殖行为及其机理,也有助于制定松褐天牛防控策略。
猜你喜欢
作物研究(2021年6期)2021-12-25
南方林业科学(2021年3期)2021-07-28
甘肃农业科技(2021年6期)2021-07-25
湖南农业大学学报(自然科学版)(2021年2期)2021-05-06
科学导报(2019年35期)2019-09-03
江苏农业科学(2017年7期)2017-05-23
意林(2015年1期)2015-05-14
山东农业科学(2014年9期)2015-01-07
绿色科技(2013年10期)2013-08-28
