
基于EST-SSR 和种子特征的苇状羊茅遗传分析
2022-03-05 06:46王子玥刘凌云常智慧
草业科学 2022年1期
王子玥,刘凌云,刘 曼,常智慧
(北京林业大学草业与草原学院,北京 100083)
苇状羊茅(Festuca arundinacea)是禾本科羊茅属的多年生草本植物,是高度自交不亲和的异源六倍体(2n=6x=42)植物,染色体为PPG1G1G2G2[1]。因其冬季高产,耐粗放管理,成坪效果好,同时具有能储存重金属等优良特性,在牧草生产、草坪绿化、生态修复等方面被应用广泛[2]。
种质资源是作物育种的基础,开展种质资源的遗传背景研究、多样性分析对遗传育种和种质资源收集、鉴定具有重要的意义[3]。与传统评价方法相比,DNA 标记由于受环境影响较小,更加能够反映真实的遗传多样性[4]。SSR 标记根据来源可分为基因组SSR 和表达序列标签SSR (EST-SSR)[5]。ESTSSR 是基于表达序列标签开发微卫星的一种新型分子标记[6]。目前EST-SSR 标记被广泛应用于遗传多样性研究[7],比如研究者[8]采用RAPD 和EST-SSR两种标记探究36 份苇状羊茅的遗传多样性,其中RAPD 和SSR 都显示出高多态性。从株高、分蘖、粗蛋白等农艺性状结合EST-SSR 标记,探究115 份苇状羊茅的遗传差异,发现其中100 份供试材料的遗传多样性水平较低[9]。
种子大小与幼苗竞争能力、休眠、萌发、出土生长率、植物幼苗大小、花的大小、植物寿命以及植物发育过程都密切相关[10]。目前关于植物种子大小遗传规律的研究,主要在黄瓜(Cucumis sativus)、花生(Arachis hypogaea)、大豆(Glycine max)等作物中[11-13]。苇状羊茅相关的遗传分析,多采用分子标记或者将分子标记与生长发育习性、植物学特性、品质性状等指标结合。但针对草种形态的研究鲜有报道。
随着生态建设的推进,苇状羊茅育种的目标越发多元化,其遗传背景与分类工作显得尤为重要。为了解不同来源苇状羊茅种质的亲缘关系,以及开展不同种质资源的科学评价,本研究结合原产地差异、EST-SSR 标记对36 份苇状羊茅进行遗传多样性分析,并根据其种子表型性状进行聚类分析,以期探究36 份不同来源苇状羊茅的遗传差异,进而为苇状羊茅遗传资源的鉴定、核心种质创新和新品种选育提供参考依据。
1 材料与方法
1.1 种质资源的收集
试验共收集到36 份苇状羊茅(25 份野生品种,10 份引进品种,1 份地方品种)为材料,其中35 份由全国畜牧总站提供,另外1 份为商业品种,由北京正道种业有限公司提供(表1)。36 份材料分别来自17 个国家涵盖了欧洲(芬兰、波兰、罗马尼亚、荷兰、法国、摩纳哥、捷克、挪威、保加利亚、南斯拉夫、希腊)、大洋洲(新西兰)、亚洲(中国、哈萨克斯坦)、非洲(突尼斯)、北美洲(加拿大、美国)。其中中国的13 份材料分别来自北京、新疆、江苏、青海、贵州、云南、四川、甘肃8 个地区。
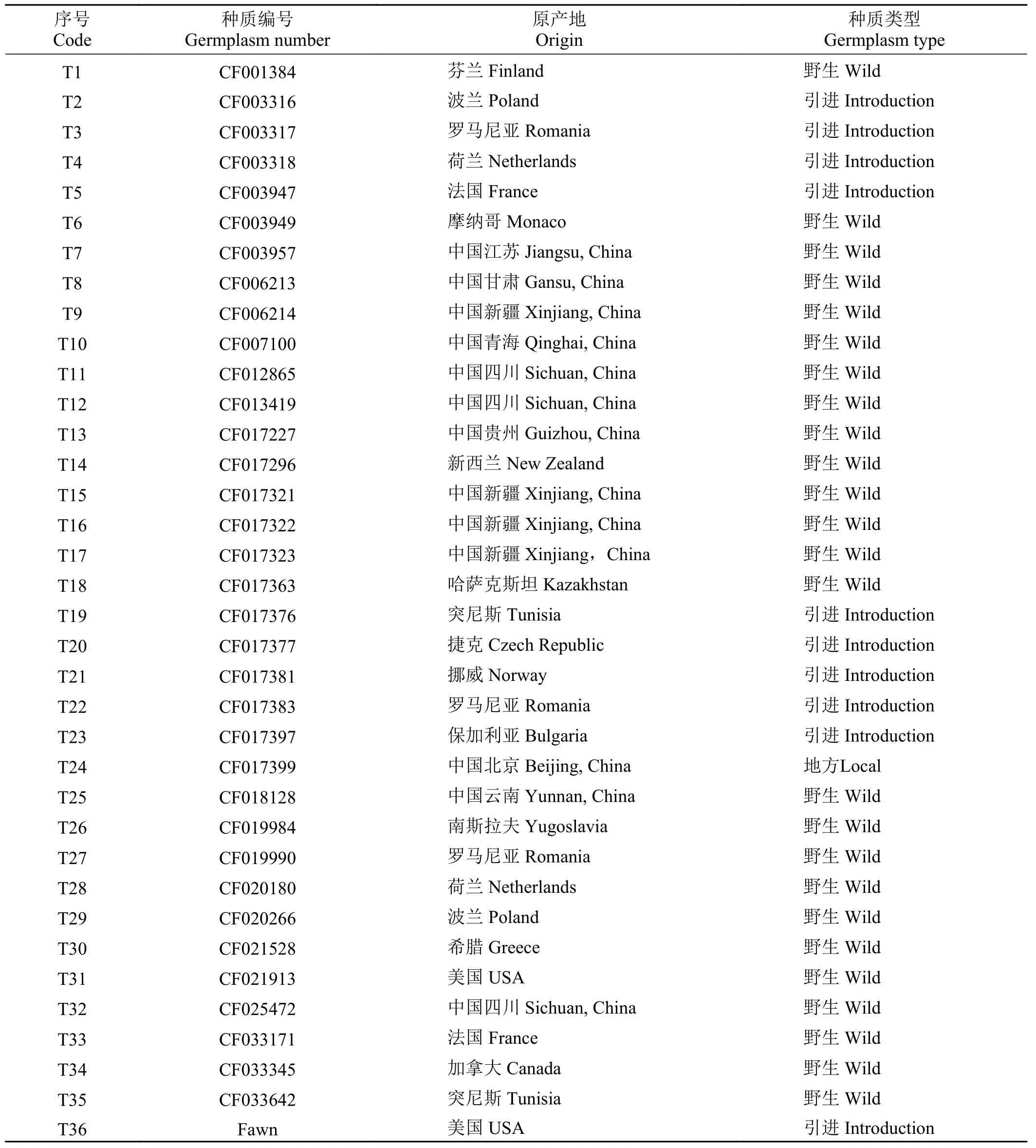
表1 36 份苇状羊茅种质信息表Table 1 Information of 36 tall fescue germplasms
1.2 种子特征的测定
测定方法参考《NYT 2127-2012 牧草种质资源田间评价技术规程》[14]。种子长、宽、厚:以风干后的成熟饱满干籽粒为观测对象,分别随机抽取10 粒干种子,测量最长、最宽、最厚处的距离。千粒重:由于初始种子数量较少,随机选取种子50 粒称重,重复3 次,数据保留两位小数。
1.3 基因组DNA 的提取和EST-SSR 标记
每份供试材料选取15 粒种子,于2020年9月10日在培养箱中进行萌发与预培养[25 ℃/20 ℃,16 h/8 h(光照/黑暗),光强24 000 lx,湿度65%]。3 周后,各挑选10 株以30 cm 固定行间距移栽至三顷园苗圃(40°00′24″N,116°19′60″E)。2020年11月13日,每份材料随机选择5 棵单株,以相同量生长良好的嫩叶进行混合,液氮充分研磨[15]。采用Omega 试剂盒提取DNA。提取完的DNA 使用Nanodrop 2000 检测浓度,其OD260/OD280均在1.7~2.0,并使用2%琼脂糖电泳凝胶检测纯度。最后将试样浓度稀释至30 ng·μL-1,在3 个离心管中各分装100 μL 液体,-20 ℃保存,用于PCR 反应。
EST-SSR 引物由文献[16]设计的157 条引物中随机挑选20 条(北京睿博兴科生物技术有限公司合成),其中14 条引物扩增效果良好(表2)。10 μL PCR反应体系含有5 μL 2×Taq Master Mix (KT205,北京天根有限公司),0.2 μmol·L-1各引物,2 μL 样品DNA和2.6 μL 无菌水[17]。利用Bio-Rad T100 PCR 仪进行PCR 扩增反应,反应程序94 ℃初变性3 min,94 ℃变性30 s,特定温度退火30 s 和72 ℃延伸60 s,进行34 个循环,在72 ℃延伸10 min。将PCR 产物在8.0%聚丙烯酰胺非变性凝胶上分离并通过银染法显现,使用Bio-Rad 全能型凝胶成像分析系统拍照保存,用于后续分析[18]。

表2 14 对苇状羊茅 EST-SSR 标记信息Table 2 Information of 14 pairs EST-SSR markers in Tall fescue
1.4 数据统计与分析
由于供试材料多为六倍体且取样为多个单株混合取样,标记的一个位点可能在6 个亚基因组中出现多条条带,故无法按照二倍体共显性标记,记录基因型[19]。PCR 扩增产物只能在某一位点上按“有”或“无”进行统计,有条带的标记为“1”,无条带记为“0”。
种子特征数据采用SPSS 23 进行方差分析,R 4.04 绘制方差分析图和K 均值聚类图;遗传距离和聚类分析采用NTSYS 软件分析[20];遗传信息中,观测等位基因数(Na)、有效等位基因数(Ne)、基因多样性指数(H)、Shannon 信息指数(I)采用POPGEN 32 计算[21];引物的多态性信息含量(PIC)采用公式:

式中:PICi为标记i的多态信息量;fi为第i种等位基因占总基因数的比率;1 -fi是缺少的基因频率[22]。
2 结果与分析
2.1 EST-SSR 条带分析
使用表2 中20 条引物对36 份苇状羊茅进行了EST-SSR 标记的扩增。结果表明,14 对引物共扩增140 条带,多态性条带总数为133 条,多态性条带百分率95%;平均每对引物扩增出10 个条带,14 对引物所扩增出的多态条带数在8 个(NFA021)~13 个(NFA035)之间,平均多态条带数为9.5 个(图1)。36份参试苇状羊茅资源的多态信息Na、Ne、H和I平均值分别为1.966、1.446、0.271 和0.420 (表3)。说明各苇状羊茅样品之间的遗传多样性处于较高水平。计算其遗传距离与遗传相似性,平均遗传距离为0.576,结果发现各样品之间的遗传多样性处于较高水平,遗传基础较宽。PIC值最大为0.346,平均值为0.27,这表明引物具有很好的有效性。
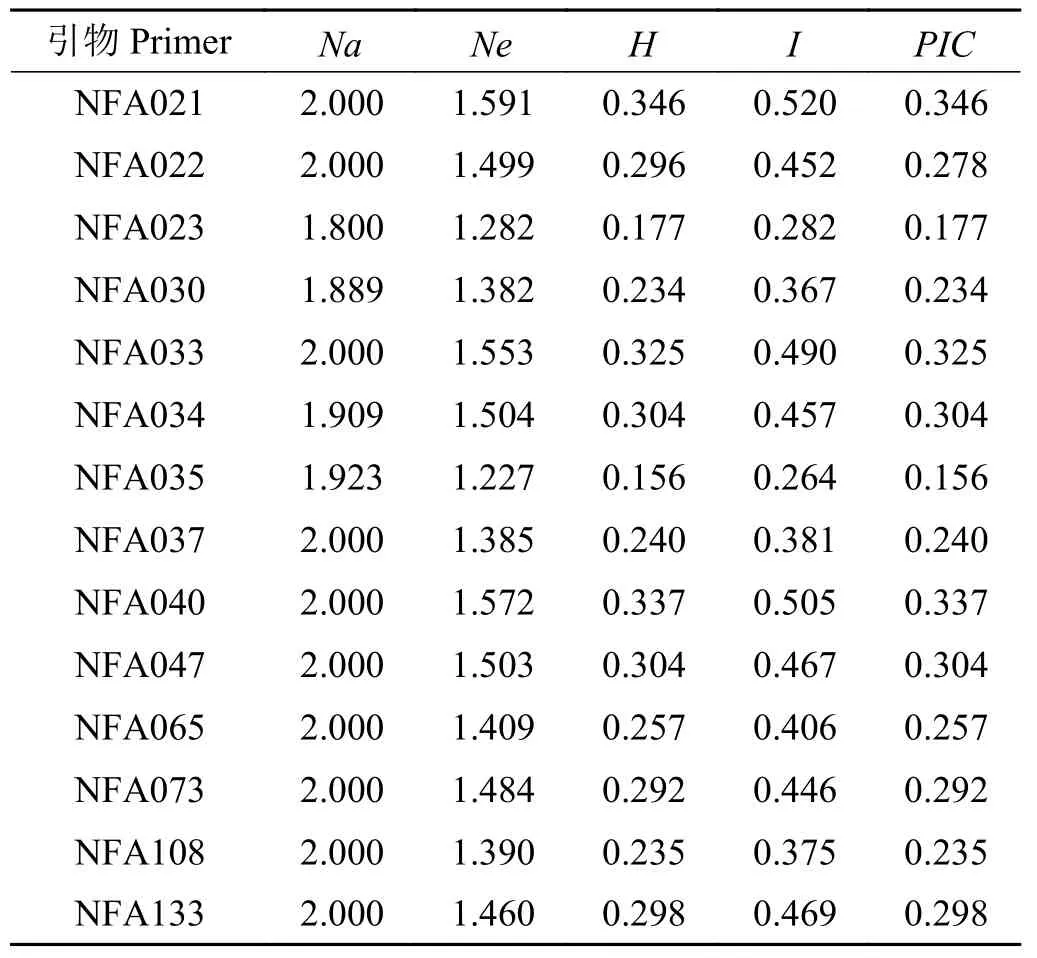
表3 36份苇状羊茅遗传多样性指数Table 3 Genetic diversity index of 36 tall fescue germplasms

图1 引物NFA047 对36 份苇状羊茅种质的扩增条带Figure 1 Amplified bands of 36 tall fescue germplasms by primer NFA047
2.2 UPGMA 聚类分析
对36 份苇状羊茅材料的SSR 分子标记结果进行UPGMA 聚类分析,得到UPGMA 聚类图(图2),其相关性系数为r=0.913,说明聚类结果可靠。在遗传距离为0.63 时,可将36 份苇状羊茅分为主要3 类和1 个混合群体。其中T1 和T13 被聚为一类,T15、T16、T17 和T18 被聚为一类。T8 和T9 的遗传距离相同,遗传距离最高为0.85。当遗传距离为0.46 时T23 被单独聚为一类。

图2 36 份苇状羊茅资源的UPGMA 聚类Figure 2 UPGMA dendrogram of 36 tall fescue germplasms
2.3 种子表型的K-均值聚类
测定36 份供试材料的形态学指标,36 份种子长度在5.39~7.67 mm,宽度在0.87~1.29 mm,厚度在0.33~0.74 mm,千粒重在1.53~3.55 g (表4),四者的变异系数分别为8.72%、10.43%、18.72%和19.18%。

表4 36份苇状羊茅种子特征数据Table 4 Data on the seed characteristics of 36 tall fescue germplasms
采用R (版本4.04)对36 份种子的表型性状测量数据进行聚类分析(图3),在K=4 时,种子特征为短、窄、薄、千粒重小的T6、T22 和T33,3 个品系被聚为一类,这与遗传距离在0.54 时的UPGMA 聚类结果相同。T21 和T3 等品系则因为种子长度长、宽度宽、厚度厚、千粒重大的特点被归为另一大类。
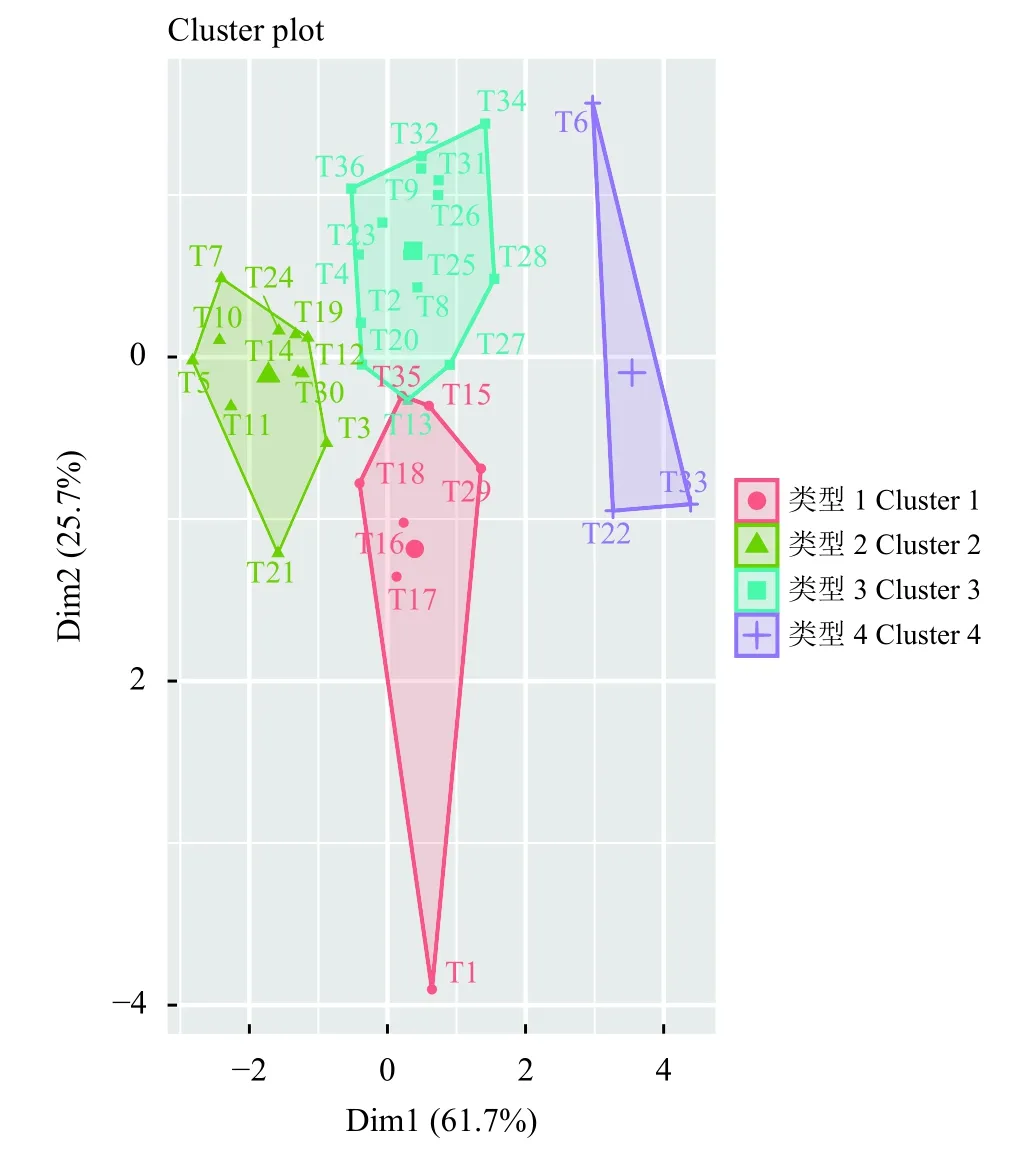
图3 36 份苇状羊茅种子表型的系统聚类分析Figure 3 System cluster analysis of 36 tall fescue germplasms
2.4 不同地理区划的苇状羊茅种子特征平均值、标准差及变异系数
受样本数限制,本研究取亚洲群体、欧洲群体和北美洲群体共计33 份材料,探究不同地理区划的苇状羊茅种子特征平均值、标准差和变异系数(表5)。其中亚洲群体的种子的长、宽、高以及千粒重数据均最大,但是长、宽、高数据在3 个群体内均不存在显著性差异(P>0.05)。欧洲群体的平均千粒重为2.36 g,低于亚洲、北美洲群体,且差异显著(P>0.05)。另外北美群体4 个种子特征的变异系数最小范围为2.04%~5.65%,欧洲群体种子高度和千粒重变异系数最大,分别达到了25.02%和23.35%。
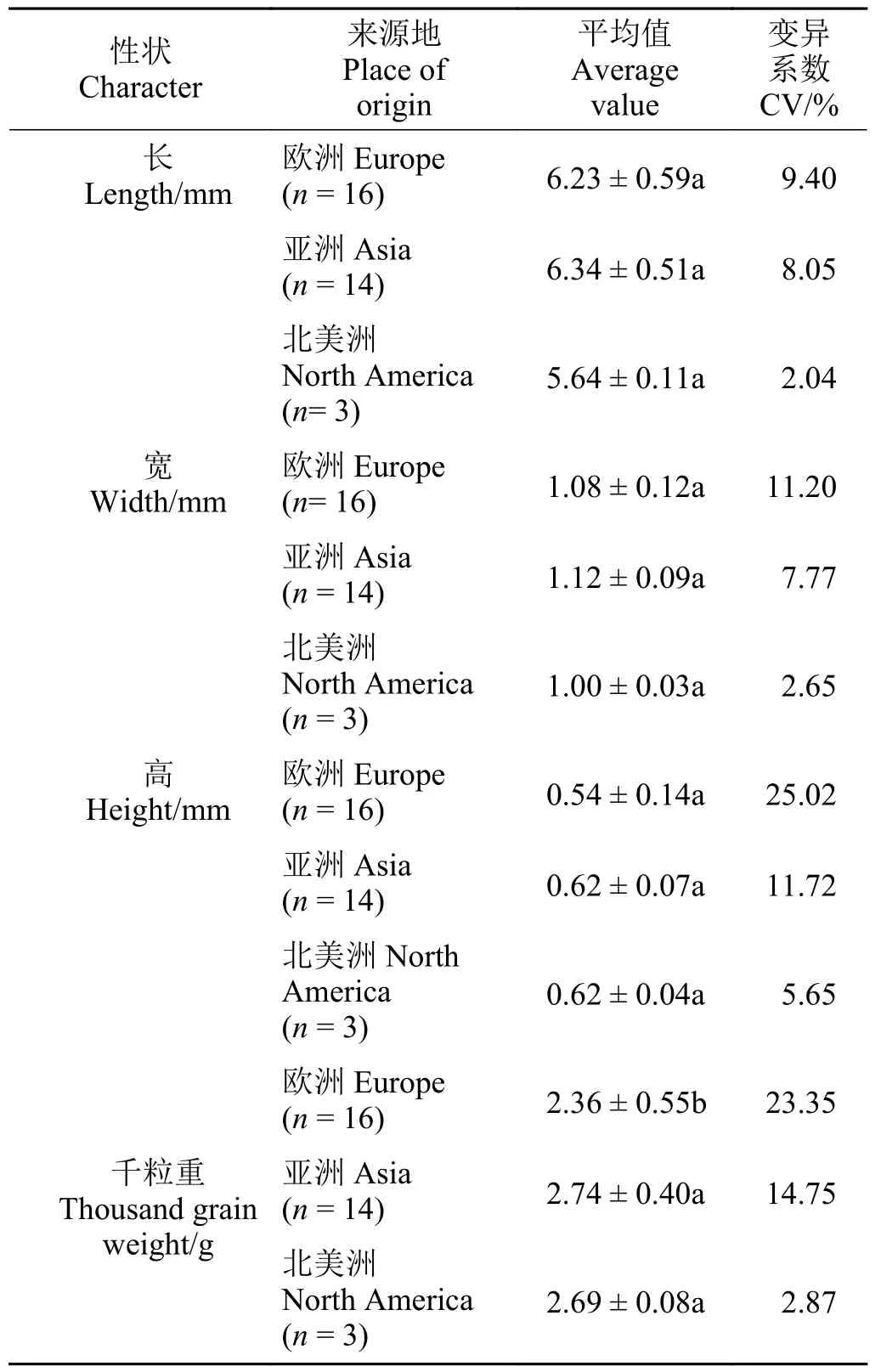
表5 不同区划种子特征上的差异Table 5 Differences in seed characteristics between zones
3 讨论
我国目前育成的草种新品种,品种间差异小、同质化等问题比较突出[23],扩大种质地理来源的跨度以及开展多角度种质资源评价是直接、有效的方法。遗传多样性分析是作物改良计划的一个重要步骤[24]。目前,针对苇状羊茅种质的遗传分析已经做了许多研究,但是却忽略了草种子特征的差异。有研究表明,大粒种子比小粒种子拥有较大的胚乳或子叶,能够为幼苗提供更多的营养物质,在苗期能忍受更大的环境压力,对后代幼苗贡献较大[25]。2019年,刘加文等[23]指出提高国产草种供给质量和效率,保障有力的国产草种有效供给,才能加快发展中国草种,赢得产业发展主动权。种子质量是草种业发展的关键,因此种子特征研究显得尤为重要。本研究以36 份来自17 个国家的种质为对象,从地理区划、EST-SSR 标记、种子特征3 个方面探究种质资源的差异。
EST-SSR 是分析苇状羊茅遗传多样性的有利工具。研究结果显示,36 份种质高度异质,遗传距离0.29~0.85,高于李家丽等[26]、Shahabzadeh 等[27]对不同苇状羊茅品种(系)的分析结果。在Shahabzadeh等[27]的研究中,引物NFA037 具有最高的多态性,但是在本研究中,PIC值最高的是NFA021,而NFA037处于中等水平。这种现象可能是种质来源差异导致的,Shahabzadeh 等[27]的材料全部收集自伊朗,而本研究的材料来自于17 个不同国家。Lou 等[28]使用90 对苇状羊茅SSR 标记对115 份苇状羊茅进行遗传分析,共得到1 010 个等位基因,平均基因多样性指数(H)为2.55,平均引物多态性(PIC)为0.211,本研究中H、PIC较上述研究略高,说明供试材料遗传背景丰富。UPGMA聚类结果有一定的地域性趋势,但是不明显。比如欧洲群体的T3、T4、T5、T20 被聚为了一类,亚洲群体的T15、T16、T17、T18 被聚为一类。但位于同一地域,比如新疆的苇状羊茅居群并没有完全聚在一类,这与李倩等[18]用EST-SSR 标记对新疆黄花苜蓿野生资源的评价结论一致。一些居群的独立聚类表明它们具有遗传特异性,不同地区的苇状羊茅有些聚为一类,表明其亲缘关系较近,可能是不同地区种质材料进化来源相同[29],也有可能是不同种质在长期的自然选择过程中存在基因交流的现象。其中T23 草种的遗传距离为0.24 差异最大,而它的种子形态却没有大差异。苇状羊茅和黑麦草曾经划分为羊茅族[30],它与属内草种乃至黑麦草属(Lolium)草种高度亲和,比如目前国审的“Johnstone” 苇状羊茅就是由“G1-316”和“G1-307”两个无性系培育,这些无性系均是通过一年生黑麦(Lolium multiflorum)与多年生黑麦草(Lolium perenne)的杂交后代与苇状羊茅再杂交而来[31]。因此这份草种可能在授粉过程中出现杂交的情况,该材料可能具有更大的研究价值。
本研究中,使用K 均值聚类法将36 份材料的种子特征分为4 大类。T6、T22、T33 被聚为一类,它们均具有短、窄、薄、千粒重小的特点,这与遗传距离在0.56 时的聚类结果相似,进一步佐证了SSR 数据的置信度。但是,K均值结果与UPGMA 结果并不完全相同。比如T15 和T29、T13 和T27 等遗传距离较远的草种却被聚为一类,可能是种子特征一般受多基因控制[32]。所以在今后研究中,有必要在构建高密度饱和遗传图谱的基础上,对控制种子大小性状的相关基因或主效数量性状基因座(QTL)进行精细定位。此外,本研究表明,欧洲群体的千粒重显著低于亚洲以及北美洲群体,其原因可能是,欧洲的典型气候是温带海洋气候,绝大部分地区气候具有温和湿润的特征[33],有利于种子的萌发,在长期的进化过程中,其养分储存需求逐降低。相反,种子的长、宽、高3 个指标在地理上并不存在显著差异,一方面可能是异花授粉作物存在广泛的基因交流,另一方面可能是本研究样本数较少,比如北美洲群体只有3 个样本,不具有代表性。
4 结论
从地理区划、EST-SSR 标记、种子特征3 个方面对36 份苇状羊茅进行遗传多样性分析发现,种质高度异质,是良好的育种材料。3 种分类结果各有异同,植物遗传背景需要从多角度分析。另外,以种子特征探究苇状羊茅的遗传差异是可行且必要的。
猜你喜欢
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
新疆农业科学(2022年6期)2022-07-13
计算机应用与软件(2022年6期)2022-07-12
中国农学通报(2022年12期)2022-06-01
自然灾害学报(2022年2期)2022-05-10
作物学报(2022年4期)2022-02-10
中国种业(2022年1期)2022-01-27
中国种业(2022年1期)2022-01-27
中国学校体育(2021年10期)2021-04-26
