
环境因子和火干扰因子对喀纳斯泰加林柳兰分布的影响
2022-03-11 09:51刘小菊褚江涛张越单奇
生态环境学报 2022年1期
刘小菊,褚江涛,张越,单奇
新疆农业职业技术学院,新疆 昌吉 831100
植物与环境关系一直是众多生态学研究的重点之一(Guisan et al.,2000),生态学家从未停止过对决定物种分布及多样性的因子的探索(Benjamin et al.,2005),这一问题的研究有助于了解物种分布的空间特征,从而为人们制定物种多样性保护对策,进一步管理、利用植物资源提供理论依据(任学敏等,2012)。众多研究表明,海拔、坡度、pH等因子对物种分布和多样性具有重要影响(Schuster et al.,2003;Medinski et al.,2010;马旭东等,2010)。
柳兰(Chamerion angustifolium)是桃金娘目柳叶菜科柳兰属的多年生草本植物,分布于我国东北,西北,华北,西南,长白山区(杨义波,2008),黑龙江省小兴安岭,老爷岭及张广才岭等林区(权曼曼等,2010),以及阿尔泰山林区。柳兰易在坡地草地上形成片植景观,花期7—8月(廖成松等,2006),开花时十分壮观,是喀纳斯自然保护区观鱼台景点主要观赏植物之一。近年来,有关柳兰的研究主要集中在引种适用性(刘旭华,2009),播种繁殖(纪福利,2015),扦插繁殖(康健,2011;钟双林等,2014),组织培养(雷颖,2003),传粉生物学特性(权曼曼等,2010)、蜜腺解剖结构(王虹等,1999),以及化学成分(田静等,2017)和矿质成分方面(姜守刚等,2018)。柳兰的生态方面的研究也有报道,廖成松等(2018)研究了野生柳兰和主要气候因子关联性。而关于野生柳兰的分布与火干扰因子关系的研究少见报道。
喀纳斯泰加林属于火成演替群落,林下也有柳兰分布,火干扰对喀纳斯森林景观(刘翠玲等,2010;师瑞峰,2012)、草本层分布、物种丰富度和地上生物量(刘小菊等,2019),土壤有机碳库(王卫霞等,2020)都有着十分重要的影响。火干扰因子和环境因子是否对柳兰的分布产生影响,哪些因子对柳兰的分布起主导作用。本文以喀纳斯泰加林林下的柳兰为研究对象,开展生态因子调查,并将生态因子分解为环境因子和火干扰因子,分析了柳兰的分布与环境因子和火干扰因子的关系,旨在为喀纳斯林下野生柳兰种群的保护提出科学的建议。
1 材料与方法
1.1 研究区概况
研究区位于新疆维吾尔自治区阿勒泰地区布尔津县境内的喀纳斯国家自然保护区实验区(87°01′45″—87°33′50″E,48°36′18″—48°38′56″N,面积875 km2)。年均气温-0.2 ℃,年均降水量1065 mm,年均蒸发量1097 mm,无霜期80—108 d,相对湿度一般为59%—90%,8月初开始降雪,一直到翌年5月下旬或6月初,常年盛行西南风,最大风力可达8级(刘小菊等,2020a;刘小菊等,2020b)。喀纳斯的山地森林是北方针叶林地带的西伯利亚山地南泰加林在南端的延伸和楔入草原地带的我国北方森林的典型代表(杨玉萍等,2019)。
1.2 取样及数据收集方法
1.2.1 火干扰因子调查
火干扰因子包括火后时间(post-fire time,PT)和林火烈度(fire severity,Se),依据林木火疤年龄分析法确定样地火干扰的发生时间(刘小菊等,2020c),调查年份与火疤木成疤时间的差值称为火后时间。根据火疤木的外在属性因子—火疤高度(Lentile et al.,2006;Halofsky et al.,2009;Keeley,2009;Santis et al.,2009)、火疤宽度和火疤深度确定林火烈度(Liu et al.,2019;刘小菊等,2020c),分为低烈度、中烈度和高烈度,并将其量化为1,2和3。
1.2.2 植物调查
于2019年7—8月和2020年8—9月,在喀纳斯泰加林内沿着海拔梯度,在有柳兰分布且存在火疤木的林分内设置乔木层样地,样地面积为 30 m×30 m,共设置35个样地,在样地的中心和4个角设置5个小样方(1 m×1 m)进行草本植物调查。记录了样地的海拔(altitude,AL)、坡度(slope gradient,SG)、坡向(slope aspect,SA)和坡位(slope position,SP)及各个物种的种类、多度、频度、数量和高度,估测草本层盖度(herbaceous layer coverage,HC)和灌木层盖度(shrub layer coverage,SC),计算乔木层样地的郁闭度(canopy density,CD)。依据坡向值分级,将坡向值划分为8类(权曼曼等,2010),北坡(0°—22.5°,337.5°—360°)1,东北坡(22.5°—67.5°)2,西北坡(292.5°—337.5°)3,东坡(67.5°—112.5°)4,西坡(247.5°—292.5°)5,东南坡(112.5°—157.5°)6,西南坡(202.5°—247.5°)7,南坡(157.5°—202.5°)8,数字越大,表示坡向越朝阳。
1.2.3 土壤调查及分析
在林冠层样方内沿坡体方向分别在上、中、下挖土壤剖面各1个,记录土壤剖面特征,分0—10、10—25和25—70 cm用100 cm3的土壤环刀进行取样,烘干后测定土壤容重(BD),用木铲分3层取样后按层混合约1 kg左右风干后去除杂物,过筛后进行实验分析。在北京时间14∶00左右用Hydra便携式土壤水分/温度测定仪(美国 EUTECH)测定与土壤采样深度一致的土壤体积水分含量(W)。
用常规方法测定土壤的理化性质(鲍士旦,2000)。电导率(HJ 802—2016,C)和pH值-电位法(GB 7859-87,H);土壤容重(GB 7833—1987,BD)-烘干法;土壤有机质-重铬酸钾氧化-外加热法(GB 7857-87,SOM);碱解氮—碱解扩散法(GB 7849-87,AN);全氮-半微量开氏法(GB 7173-87,TN);全磷-钼锑抗比色法(GB 7852-87,TP);全钾-火焰光度法(GB 7854-87,TK);有效磷-钼蓝法(GB 7853-87,AP);速效钾—1 mol·L-1中性醋酸铵浸提-火焰光度法(GB 7856-87,AK)。
1.3 数据处理与分析
1.3.1 重要值
草本重要值=(相对盖度+相对高度+相对多度)/3。
1.3.2 相对频度
物种相对频度=(该种频度/所有植物频度)×100%
1.3.3 物种多样性指数测度
草本层Patrick丰富度指数、Simpson优势度指数(D)、Shannon-Wiener多样性指数(H)和Pielou均匀度指数(J)利用马克平(1994)所列公式进行计算。
1.3.4 数据分析
所有数据的基本分析与处理均利用Office 2010办公软件完成。采用相关分析解析各生态因子间的关系,在SPSS 24.0程序中进行;运用Canoco 5.0,以物种重要值、草本层物种多样性和生态因子建立物种—生态因子矩阵,采用CCA(典范对应分析)排序解析物种重要值和草本层物种多样性指数与环境因子(18个)和火干扰因子(2个)之间的关联关系。运用蒙特卡罗置换检验物种重要值和草本层物种多样性和生态因子的相关性,检验结果显示显著(P=0.003),表明排序效果良好,可以接受生态因子对物种分布的解释。分别计算两类生态因子对物种分布的解释量。
2 结果与分析
2.1 主要草本物种组成及环境因子和火干扰因子的相关性分析
在调查的35个样地中,选择相对频度>51.43%的10个物种进行分析,柳兰样地物种组成见表1。

表1 柳兰样地的主要草本物种Table1 Main herb species in Chamerion angustifolium plots
采用Pearson相关系数对火后时间、林火烈度、郁闭度(CD)、灌木层盖度(SC),草本层盖度(HC)、海拔、坡度、坡向、坡位、土壤容重、土壤体积含水量、电导率、pH值、土壤有机质、全氮、碱解氮、全钾、速效钾、全磷和速效钾共 20个环境因子和火干扰因子进行相关分析,剔除与其他2个以上因子呈显著和极显著相关的因子,以免在进一步分析中互相干扰。结果表明,TP与C、SOM和AP呈极显著相关,相关系数分别为0.588,0.785和-0.278(P<0.01);AN与HC,C,pH、SOM和TP呈极显著相关,相关系数分别为-0.311,0.604,0.402,0.616和0.599(P<0.01);SP与AL,AS,SL分别呈极显著和显著相关关系,相关系数为-0.573,-0.347和-0.334。因此剔除TP、AN和SP 3个因子,余下17个因子进行CCA排序分析。
2.2 柳兰样地的排序分析
从CCA分析结果可以看出(图1),草本物种在 CCA排序图中较好地区分开来并呈现出一定的规律。第一轴主要反映火烈度、海拔、坡向,灌木层盖度,pH值,有机质、全钾和速效钾的变化,其中与柳兰分布相关较大的因子是林火烈度、海拔、坡向、灌木层盖度、pH值、有机质、全钾和速效钾;第二轴主要反映林火烈度、火后时间、坡向、坡位、草本层盖度、土壤容重、电导率、有机质、有效磷和全氮的变化,其中对柳兰分布影响较大的因子有林火烈度、土壤容重和有机质。对柳兰影响较大的因子是林火烈度、海拔、坡向、灌木层盖度、土壤容重、有机质、全钾和速效钾,前两轴的累计贡献率为72.48%(表2)。总体来看,大部分物种在排序图上的分布格局和环境梯度较吻合,薄蒴草(Lepyrodiclis holosteoides)和圆叶鹿蹄草(Pyrola rotundifolia)对环境梯度和火干扰因子响应不敏感。柳兰位于排序图中心靠左的位置上,海拔区间是 1314.6—1908.4 m,海拔越高柳兰分布越少,低海拔有利于柳兰的生长;火后时间(16—90a)越短越有利于柳兰更新生长;在0—14°的坡度上柳兰都有分布,在此范围内坡度越小的下坡其生长更好;林火烈度越高越有利于柳兰的分布;阳坡、阴坡柳兰均有分布,但阳坡更适合柳兰的生长;土壤有机质(31.190—71.445 mg·kg-1)、速效钾(49.511—260.690 mg·kg-1)、全钾(3.791—16.678 g·kg-1)、pH值(4.6—6.2)越大越有利于柳兰的生长;土壤容重(0.626—1.614 g·cm-3)、电导率(40.599—415.527 μS·cm-1)、灌木层盖度(3.6%—65.7%)、草本层盖度(43.6%—98.4%)和郁闭度(42.9%—79.8%)越大越不适合柳兰生长。柳兰常与野火球(Trifolium lupinaster)、老芒麦(Elymus sibiricus)、白花砧草(Galium boreale)和直立老鹳草(Geraniumrectum)伴生。柳兰分布与草本层的物种丰富度指数正相关,与其他 3个物种多样性指数负相关,4个指数之间相关性较大。

图1 柳兰样地的CCA排序图Figure 1 CCA ordination diagrams of Chamerion angustifolium
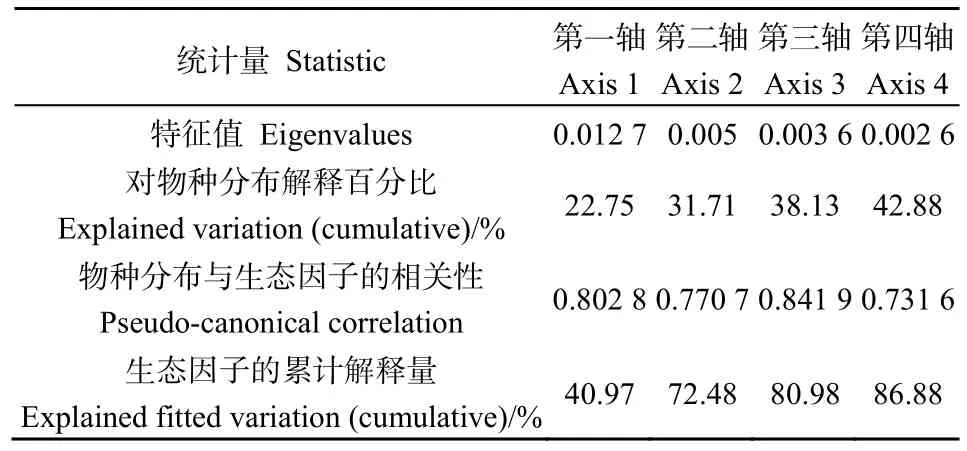
表2 CCA序轴的特征根及其与环境因子和火干扰因子之间的关系Table 2 Eigenvalues and correlation coefficients of environmental factors and fire disturbance factors for first four axes of CCA
从检验结果来看(表 3),柳兰与海拔呈极显著负相关,即海拔越高,柳兰分布的越少,达到一定的海拔高度,几乎无柳兰分布;与土壤有机质极显著正相关,表明柳兰在富含有机质的土壤中生长更好。柳兰分布与林火烈度呈显著正相关,林火烈度越高,形成的林窗越大,越容易为灌木和草本的定植创造条件,灌木的快速更新为柳兰的发芽和生长提供荫庇条件。柳兰分布与坡向、pH值、速效钾和全钾呈显著正相关,阳坡的光照条件优于阴坡,因此柳兰在阳坡生长更好,而火烧后的灰分进入土壤提高了土壤的pH值和速效钾和全钾,表明柳兰在火烧迹地可以很好地生长;与土壤容重和灌木层盖度表现为显著负相关,土壤容重越小,土壤疏松透气,有利于柳兰根系生长,灌木层盖度越大越不利于柳兰的生长。以上分析表明柳兰适合在高烈度火烧林分中更新,适合在海拔低、阳坡、灌木层盖度小、土壤呈弱酸性、土壤容重小、有机质、全钾和速效钾含量高的环境中生长。

表3 CCA排序轴与环境因子和火干扰因子的相关系数Table 3 Correlation coefficients of the CCA ordination axes and the environmental and fire disturbance factors
2.3 环境因子对柳兰物种分布格局影响的定量分析
依据Borcard et al.(1992)的方法,环境因子和火干扰因子对物种分布影响的定量分离过程由 4个步骤完成定量分离(杨玉萍等,2019):(1)环境因子独立作用于物种分布部分;(2)火干扰因子独立作用于物种分布部分;(3)物种分布与环境和火干扰因子交互作用于物种分布部分;(4)环境因子和火干扰因子未能解释的部分。
定量分离结果显示(图2),20个生态因子分为环境因子和火干扰因子,环境因子解释的部分占41.9%,火干扰因子解释的部分占7.2%,二者交互作用解释部分占 39.1%,不能解释部分占11.8%。未解释部分可能是因为森林群落内部的自身作用和随机因素对物种分布的影响。

图2 生态因子的定量分解图Figure 2 Quantitative separation of ecological factors
3 讨论
林火烈度,海拔,坡向、灌木层盖度,pH值,有机质,土壤容重,速效钾和全钾对柳兰有着显著的影响。山地森林植被理论通常认为海拔变化是控制群落物种组成最主要的因素,海拔上升,导致温度、降水、地形有所不同,从而影响土壤湿度和光照条件,进而影响物种分布和个体生长(Zhao et al.,2005;Wimberly et al.,2008)。喀纳斯山地森林的柳兰在海拔梯度上分布比较明显,随着海拔升高,柳兰分布越来越少。许多研究证明,在环境因子中,海拔梯度为山区物种分布的主要控制因子(Poulos et al.,2010;Stella et al.,2013;Wang et al.,2014)。与本研究中海拔是柳兰分布的主要因子的结论一致。随着林火烈度增大,受干扰林分在物理基质及能量配置方面的重组程度加大,林分重新发育的可能性增大,从而给一些适生树种和草本及灌木提供了生存机会(Lentile et al.,2006)。坡向决定受光方向、光照强度和光照长度(马勇等,2021),阳坡的光照强度大于阴坡,柳兰在阳坡生长更好,表明柳兰是阳性植物。林火烈度越大形成的林窗越大,光照越多,有利于柳兰种子萌发,表明柳兰是火烧后的先锋植物,这与廖成松等(2018)发现光照强度是影响柳兰生长的主要因子之一的结论一致。本研究发现郁闭度低更适合柳兰的生长,灌木层的盖度小利于柳兰的分布,表明柳兰对光照的要求很高,光照强度大能促进柳兰的生长,再次证实柳兰属于喜光植物。
在本研究的pH值范围内,柳兰在微酸性的土壤环境中生长更好,火后时间越长的林分土壤 pH值越小;火后时间越短,土壤的pH值越大,呈微酸性,这是因为火烧后灰分进入土壤中,提高了土壤的pH值。在引种柳兰时注意调节土壤的酸碱性。此外,本研究中,全钾和速效钾是影响柳兰生长和分布的重要因子。钾元素不仅是植物生长发育所必须的营养元素,钾元素对植物具有抗寒冷的作用,海拔越高,植物枝、叶中的钾含量也会越高(Masood et al.,2016)。Certini(2005)研究发现森林火灾可以将固定在地表土壤有机质和凋落物中有机态的磷和钾元素以无机态的形式释放出来,这些被释放出来的无机态磷和钾元素随着灰分物质的沉降返回到森林地表,从而增加了土壤磷和钾元素的含量。相关研究发现植物生长每年要平均需消耗约112—224 kg·hm-2的 K+(Alcañiz et al.,2016)。阳离子交换量反映了土壤的保肥能力(Costa et al.,2004)。喀纳斯泰加林经过火烧后,土壤中的钾含量增加,但随着植被的更新,它们生长发育需要大量的速效钾,而且K+在土壤中流动性较强,容易被淋溶流失,导致土壤中速效钾含量降低。所以柳兰表现与全钾和速效钾呈显著正相关。本研究中柳兰的生长还受到土壤有机质和土壤容重的影响。土壤有机质是衡量土壤肥力的指标之一,相关研究表明,有机质含量越高越能促进植物的生长(Qu et al.,2014),与本研究中有机质高促进柳兰生长的结论一致。Li et al.(2006)研究发现土壤容重降低后土壤孔隙度、土壤含水量、团聚体稳定性以及饱和水力传导度均显著增加。相关研究表明减小土壤容重,增加土壤有机质含量,土壤肥力提高,利于植被的生长(刘效东等,2011),与柳兰分布与土壤容重显著负相关的结论一致。
在本研究生态因子范围内,草本层物种越丰富,柳兰分布的可能性就越大,而草本层优势度指数、均匀度指数和多样性指数越大,柳兰出现的可能性越小。黑穗薹草(Carex atrata)是喀纳斯泰加林林下最常见的优势草本,当其占绝对优势时,优势度指数增加,而柳兰出现的可能性则会降低,表明二者对环境因子的要求不同。
本研究影响因子定量分离分析结果显示,环境因子和火干扰因子解释了柳兰物种分布的88.2%,由于该研究区是自然保护区,人为干扰较少,物种分布和环境保持着良好的对应关系,这与黄治昊等(2017)对北京地区黄檗分布与环境因子的关系研究的结论保持一致。而火干扰因子对柳兰分布的影响只占7.2%,究其原因是此次调查的最近的火后时间是 16年,草本植物的寿命较木本短,因此林火对草本植物的中长期影响较难判断。环境因子和火干扰因子的交互作用占解释量的39.1%,表明火干扰受到地形的影响,同时又和地形一起共同对土壤物理、化学过程发生作用,而土壤物理、化学性质等环境因子的不同又直接关系到物种的分布和生长,因此,物种的分布是在历史过程中植物与环境、火干扰相互作用发展的结果(杨玉萍等,2019)。本研究中未能解释的部分占11.8%,而不能解释的部分主要包含随机因素和由生物间相互作用及引起的变化,随机因素包括如取样设计、测定误差等;物种之间的相互作用,如竞争排除引起的异域分布、互惠关系引起的种间联结对物种分布格局可能有一定影响(邵方丽等,2012)。总之,环境因子和火干扰因子对柳兰分布影响的定量分离结果显示了较好的排序结果,表明所选择的生态因子是影响物种分布的重要因子。
4 结论
环境因子和火干扰因子共同决定着喀纳斯泰加林林下柳兰的分布,但起决定性作用的生态因子是林火烈度、海拔、坡向、灌木层盖度、土壤容重、有机质、全钾和速效钾。在本研究生态因子范围内,柳兰适合在土壤呈弱酸性,海拔低,阳坡,灌木层盖度小、土壤容重小、土壤有机质、全钾和速效钾含量高的环境中生长。草本层物种越丰富,柳兰分布的可能性就越大。柳兰是火烧后的先锋植物,是阳性植物,但在适当遮光条件下可正常生长。柳兰与野火球、老芒麦、白花砧草和直立老鹳草对生态环境要求基本一致。环境因子和火干扰因子总共解释了喀纳斯泰加林森林植被分布变异的88.2%,环境因子占41.9%,火干扰因子占7.2%,不能解释部分占11.8%,表明环境因子和火干扰因子的解释能力较好,物种分布和环境保持着良好的对应关系。在引种驯化柳兰时要注意它对光照和土壤酸碱性的要求。
猜你喜欢
中国地震(2022年1期)2022-06-24
地震研究(2021年3期)2021-11-10
建材发展导向(2021年15期)2021-11-05
山西林业(2021年2期)2021-07-21
建材发展导向(2021年10期)2021-07-16
中国新闻周刊(2020年6期)2020-03-08
湖北农业科学(2019年10期)2019-07-08
安徽农业科学(2018年15期)2018-05-14
科技资讯(2017年3期)2017-03-25
草原与草坪(2013年3期)2013-10-22
