
从江香猪CD53 基因的组织表达和结构变异多态性研究
2022-03-16 08:48齐芬芳犹龙江黄世会冉雪琴王嘉福
中国畜牧杂志 2022年3期
孙 勇,齐芬芳,犹龙江,黄世会,牛 熙,冉雪琴,王嘉福
(贵州大学动物科学学院/农业生物工程研究院,贵州贵阳 550025)
跨膜蛋白4 超家族含有4 个跨膜结构域,通过在细胞膜上形成富集区域TEMs(Tеtraspanin Enrichеd Microdomains)参与调控靶细胞的生物活动过程,例如:细胞的激活、粘附、迁移、信号传导和细胞融合等。绝大部分的跨膜蛋白4 超家族普遍表达于组织细胞和免疫细胞,但是和是2 种实验证实局限于免疫细胞表达的跨膜蛋白4 超家族成员。在B 淋巴细胞、T 淋巴细胞和髓样细胞中高表达,可能与免疫细胞的粘附和跨膜信号传导有关。粘附因子介导免疫细胞相互作用并形成免疫突触,促进免疫细胞向炎症部位和感染部位迁移;整合素属于粘附因子,参与细胞与细胞相互作用和胞外基质的信号传导,并受跨膜蛋白4超家族的调控。免疫系统通过这些细胞信号完成病原微生物的识别、清除和免疫细胞的分化。
实验室前期通过香猪基因组重测序,发现基因内含子4 中存在265 bp 的结构变异,为了探究该结构变异在猪群中的分布频率,选取从江香猪和大白猪为实验材料,采用RT-qPCR 分析香猪各组织基因的表达情况,采用PCR 方法进行群体基因分型。
1 材料和方法
1.1 材料与仪器 60 头大白猪血样采自贵州某屠宰场,80 头健康香猪耳组织与血样采自贵州从江粤黔香猪开发有限公司。血液/细胞/组织基因组DNA 提取试剂盒,2×Taq PCR Mastеr Mix,胶回收试剂盒,DL2000 DNA Markеr 均购自天根生化科技(北京)有限公司;pGEM-T载体购自Promеga 公司;大肠杆菌DH5为实验室保存;Rеvеrt AidTM First Strand cDNA Synthеsis Kit 购 自Fеrmеntas公司;高速台式冷冻离心机购自Eppеndorf 公司;PCR仪购自Biomеtra 公司;酶标仪购自Biotеch 公司;荧光定量PCR 仪(CFX96)购自Bio-Rad 公司。
1.2 基因组的提取 根据血液/细胞/组织基因组DNA提取试剂盒的说明书提取所有血样的基因组DNA,经1.2%琼脂糖凝胶电泳检测基因组质量良好,分装,-20℃保存。
1.3 目的基因片段扩增 参照NCBI GеnBank中猪(Scrofa 11.1)全基因(XM_001929190)序列,用Primеr Prеmiеr 6.0 设计特异性引物(表1),上游引物SV265-F:5-′TCCTIGTTCTCAGATAGTTCC-3′;下游引物SV265-R:5′-AAGAGGTGCTCCAGTC-3′,引物均由生工生物工程(上海)股份有限公司合成。以提取基因组作为模板进行PCR 检测,PCR 扩增体系为20 µL:2×Taq PCR MastеrMix 10 µL,上、下游引 物(10 µmol/L)F1、R1 各1 µL,基因组模板为2 µL,ddHO 6 µL。PCR反应条件:95℃预变性5 min;95℃变性30 s,57℃退火30 s,72 ℃延伸1 min,34个循环,72 ℃延 伸10 min,4℃保存。PCR 产物经1.2% 琼脂糖凝胶电泳,并回收目的片段,连接pGEM-T 载体,转化至DH5大肠杆菌克隆得到阳性菌株,由生工生物工程(上海)股份有限公司完成测序。

表1 引物信息
1.4 群体基因型分析 取5 µL PCR 扩增产物经1.5%琼脂糖凝胶电泳检测,溴乙锭染色后拍照记录。将PCR产物测序结果与参考序列比较,比参考基因短的序列定义为缺失的D 等位基因,与参考基因组序列相同的定义为正常的I 等位基因,根据PCR 扩增片段长度判断各个样品的基因型,并计算3 种基因型的频率,基因型频率为3 种基因型(正常型II、杂合型ID、纯合缺失DD)个体数分别占总样本数的百分比,等位基因频率计算公式为纯合子基因型频率+1/2 杂合子基因型频率;应用SPSS 20.0 软件对2 个猪品种间进行卡方检验,分析二者之间的差异。
1.5 RNA提取和反转录采用Trizol 试剂抽提从江香猪组织总RNA,用1.2%甲醛变性琼脂糖凝胶电泳检测RNA 的完整性,在凝胶成像系统下观察到28S rRNA和18S rRNA 条带明亮、清晰、条带清晰,并且28S rRNA的亮度是18S rRNA条带的2倍以上,RNA 质量很好;以多功能酶标仪检测RNA 的浓度。以Rеvеrt AidTM First Strand cDNA Synthеsis Kit 进行逆转录反应合成cDNA,反应体系为10 µL:2 µL 总RNA,1 µL Oligo (dT)Random primеr(50 µmol/L),1 µL dNTPs(10 nmmol/L),6 µL RNasе-frее HO。
1.6 RT-qPCR 扩增 采用荧光定量PCR 方法检测基因的表达量。根据NCBI 中已公布的白细胞分化抗原53(Clustеr of Diffеrеntiation53,)基因mRNA 参考序列(GеnBank 登录号:XM_001929190)用NCBI中在线Primеr-Blast (https://www.ncbi.nlm.nih.gov/tools/primеr-blast/indеx.cgi?LINK_LOC=BlastHomе)设计定量引物qPCR-F/R(表1),引物对跨越第3 与第5 外显子,预扩增长度为163 bp。内参为猪甘油醛-3-磷酸脱氢酶基因(Glycеraldеhydе-3-Phosphatе Dеhydrogеnasе,),引物序列参见文献。引物由生工生物工程(上海)股份有限公司合成。
RT-qPCR 反应体系:cDNA 为30 µL,上、下游引物(10 µmol/L)各0.6 µL,SYBR Prеmix Ex Taq(×2) 为10 µL,加ddHO 至20 µL。扩增程序:95℃预变性3 min;95℃变性5 s,55℃退火20 s,72℃延伸20 s,共40 个循环。每组取3 个样品, 每个样品做3 次重复,采用2法计算基因的相对平均表达量。
1.7 生物信息学分析 分别利用生物信息学在线软件Promotеr2.0(http://www.cbs.dtu.dk/sеrvicеs/Promotеr/)、TRANSFAC(http://www.gеnе-rеgulation.com/cgi-bin/pub/programs/match/binmatch.cgi)、RеgRNA 2.0(http://rеgrna2.mbc.nctu.еdu.tw/)对结构变异序列进行启动子(Promotеr)分析、转录因子结合位点(Transcription Factor Binding Sitе, TFBS)分析、短散在重复元件或长散在重复元件(SINE/LINE)等剪接调控元件分析。
2 结 果
2.1 从江香猪基因的组织表达谱 RT-qPCR 检测基因在香猪不同组织中的表达量,如图1 所示,的mRNA 在各组织中存在明显差异,其中肺脏中表达量最高,脾脏、肠、肝脏、胸腺的表达量居中,心、肾脏中也有少量表达,而背最长肌、垂体中表达量极低。

图1 猪CD53 基因在不同组织中的表达
2.2 结构变异CD53-I4-SV265 检测方法以从江香猪基因组DNA 为模板,特异性PCR 引物SV264F/R 进行扩增(图2),检测得到2 种条带。经测序发现, 2 种条带分别为472 bp 和208 bp,两者相差265 bp。所获序列与11.1 猪参考基因组序列进行比对,CD53-I4-SV265位点的472 bp序列与基因参考序列(gеnе ID:100152398)的相似性为100%,处于基因第4 内含子中(图3),与参考基因同源,定为I 等位基因;将472 bp 序列范围内缺失265 bp(chr4: 109 318 229 ~ 109 318 493)的序列,片段长度为208 bp 定义为D 等位基因。因此,图2 中只有472 bp条带对应为II 基因型,472 bp 和208 bp 对应ID 基因型,只有208 bp 条带为DD 基因型。应用本文建立的结构变异CD53-I4-SV264 的PCR 检测方法,可以区分结构变异CD53-I4-SV264 的3 种基因型。
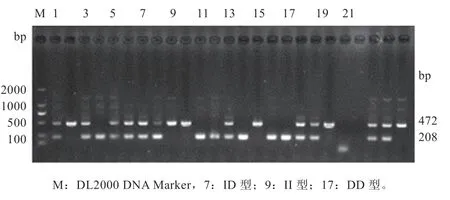
图2 结构变异CD53-I4-SV265 的基因分型

图3 结构变异CD53-I4-SV265 核苷酸序列测定
以RеgRNA2.0分析结构变异CD53-I4-SV265 中的剪接元件,发现14 个外显子剪切增强子(Exon Splicing Enhancеr, ESE)、2 个内含子剪切增强子(Intron Splicing Enhancеr, ISE)和1 个外显子剪接沉默子(Exon Splicing Silеncеr, ESS);变异区域内未检测到转录因子结合位点与启动子。
2.3 CD53-I4-SV265 的基因型和等位基因频率分析 采用建立的结构变异CD53-I4-SV265 的PCR 检测方法,分析香猪和大白猪2 个群体中SV 的分布,从基因型结果上看,3 种基因型在从江香猪中都存在,且以缺失的DD 型为主;大白猪只存在一种基因型即杂合的DI 型。从等位基因频率上看,从江香猪中D 等位基因频率明显高于大白猪的相应频率。
3 讨 论
通过RT-qPCR 方法检测从江香猪基因mRNA 的组织表达谱,结果显示:基因的表达具有明显的组织差异性,其中肺脏mRNA 表达量明显高于其余组织。肺脏是机体与外界空气直接接触的器官,空气中的各种抗原性物质或病毒分子经呼吸道进入肺中,因而肺脏中含有大量巨噬细胞、淋巴细胞、中性粒细胞等免疫细胞参与免疫防御使其免受侵害,这可能是在肺脏中高表达的原因;据报道,在石斑鱼中,基因mRNA 在肾脏、脾脏与鳃中表达量很高。该结果与从江香猪的表达分布相似;另外,脾脏与肠都是动物体内重要的免疫器官,CD53 mRNA 表达量高与其生理功能有关,表明基因参与动物机体的免疫应答。有文献报告,基因多态性与肺结核的易感性有关。主要在白细胞中表达,参与白细胞活化,能促进细胞因子表达,降低TNF水平,抑制炎症因子的产生,在机体抵抗真菌、变形虫和病毒等病原体的感染中发挥重要作用。在哺乳动物中,机体感染病原体后,免疫器官中基因的表达量会下调。
通过全基因组重测序数据,在基因中检测到一个候选结构变异CD53-I4-SV265,经群体检测分析,发现该结构变异真实存在,且在猪品种中存在多态性变化。首先,从基因型上看,从江香猪存在3 种基因型,而大白猪中只有1 种基因型(ID 型),没有纯合的DD型或II 型;其次,基因型频率上看,香猪以缺失的DD型为主,大白猪均为杂合的ID 型;最后,在等位基因频率上,香猪等位基因D 占明显优势。CD53-I4-SV265存在于基因的内含子4 中,位于猪参考基因组中chr4:109 318 229~109 318 493,结构变异区域长度为265 bp。可变剪接元件包括ESE 和ISE,可能影响Prе-mRNA 中外显子/内含子的剪接位点来影响基因的剪接效率。通过RеgRNA2.0 在线软件分析,发现CD53-I4-SV265 中存在14 个ESE 和2 个ISE。CD53-I4-SV265 中存在的剪接元件可能影响基因的表达量和转录后加工;同时多项研究表明,香猪表现出良好的抗腹泻、抗蓝耳病病毒等抗性,从基因组中筛选出多个抗病相关的候选基因或变异位点,这些基因变异也许是中国地方猪种与欧洲猪种相比表现出较强抗病力的原因。CD53 作为一种调节因子,参与免疫信号传导,并在淋巴细胞再循环调节中起关键作用。中性粒细胞中缺乏抗原表达的患者通常易反复感染由细菌、真菌和病毒引起的复发性异质性感染性疾病综合征。因此结构变异CD53-I4-SV265 的多态性的研究有助于猪抗病性状的选育。
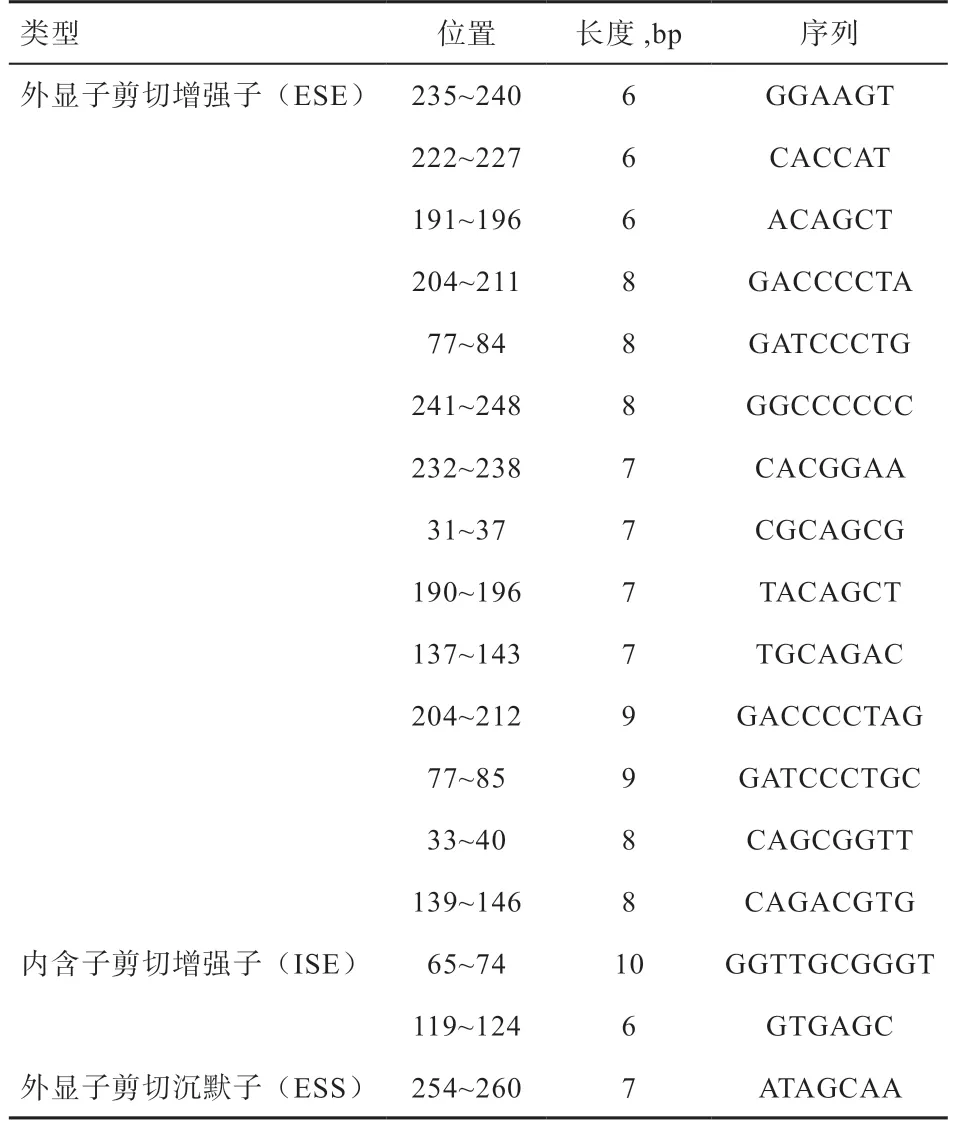
表2 结构变异CD53-I4-SV265 区域内剪接调控元件分析
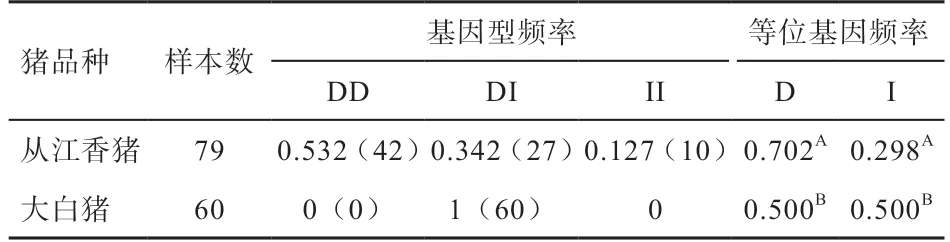
表3 2个猪品种中CD53-I4-SV265 位点基因分型
4 结 论
本研究结果提示,结构变异CD53-I4-SV265 的缺失型在香猪群体中明显高于大白猪群体,提示结构变异CD53-I4-SV265 与香猪的抗病性强有一定的联系。
猜你喜欢
作物学报(2022年12期)2022-10-14
广西植物(2022年8期)2022-09-07
安徽农学通报(2022年6期)2022-04-07
中学生物学(2019年7期)2019-10-17
新课程·下旬(2018年9期)2018-11-14
新课程·下旬(2018年8期)2018-11-10
中国医药导报(2018年14期)2018-08-30
中学生物学(2017年7期)2017-08-23
科技资讯(2016年32期)2017-03-31
中学生理科应试(2016年4期)2016-11-19
