
季节放牧下青藏高原高寒草甸牧草生物量空间分布特征
2022-03-21 04:41张芊妤曾奕丰李文洁王硕华王树林阿的鲁骥侯扶江
草业科学 2022年2期
张芊妤,曾奕丰,李文洁,申 洁,王硕华,王树林,阿的鲁骥,李 岚,侯扶江
(农业农村部草牧业创新重点实验室 / 兰州大学草地农业生态系统国家重点实验室 / 兰州大学草地农业科技学院, 甘肃 兰州 730020)
草畜互作是草原生态系统的主要驱动力之一。放牧草原的生产力提供了美国草原70%的畜产品、新西兰家畜95%和澳大利亚家畜90%的营养需求[1]。草原生产力是生物因素、非生物因素和社会因素共同作用的产物[1]。高寒草甸作为气候变化的生态脆弱带,对环境变化较为敏感,研究其生产力具有重要意义。
一般情况下,草原群落地下生物量占总生物量80%以上,是植物体中最为稳定的碳库,植物地下部分对草原生态系统的生产力和生态功能(缓解气候变暖、助力碳达峰和碳中和等)具有重要意义[2]。在放牧过程中,放牧家畜采食植株地上部分,通过践踏和排泄改变植被和土壤理化性质,间接作用于植株地下部分,影响草地群落[1]。植株地上生物量与地下生物量均对放牧具有显著的响应,二者之间存在显著相关性[3]。同时,二者共同影响根冠比,反映植被生物量的分配格局,是植被对环境适应性的综合表现。季节性放牧影响高寒草甸植物群落组成,以及不同功能群植被的相对生物量[4],适当的放牧强度和季节性放牧能够缓解放牧压力,促进植物再生[5]。近年来,国内对高寒草甸牧草生物量空间分布开展了大量研究,多集中于环境因子[6-7]、气候变化[8]和放牧活动[9],而对高寒草甸不同功能群与地下生物量分配关系的研究较少[10]。此外,现有地下生物量的直接测定方法存在弊端,包括土柱法、钻土芯法和内生长土芯法[11],挖掘对长期和小面积样地破坏性较大,钻土芯法均费时费工。因此,通过植物群落地上生物量与地下生物量的关系预测较为可行。
青藏高原被誉为“中华水塔”和“世界第三极”,高寒草甸约0.7 亿hm2,接近青藏高原总面积的一半[12]。高寒草甸的放牧是当地社会发展的重要途径[13-14]。本研究通过测定不同季节放牧的高寒草甸地上与地下生物量,分析生物量的空间和种间分布及其相互关系,以期为可持续的放牧管理提供科学依据和方法。
1 材料与方法
1.1 研究区概况
研究区位于兰州大学玛曲草地农业试验站,地处甘、青、川三省交汇处(地理位置为101°53' E, 33°58' N,海拔3 500 m)。研究区年平均气温2.0 °C,年均降水量611.9 mm,土壤类型主要为高寒草甸土[15],主要植物包括禾叶嵩草(Kobresia graminifolia)、垂穗披碱草(Elymus nutans)、草地早熟禾(Poa poophagorum)、小花草玉梅(Anemone rioularis) 、甘肃马先蒿(Pedicularis kansuensis)、多枝黄芪(Astragalus polycladus)等。牧草4 月下旬返青,10 月初期开始枯黄。
1.2 研究方法
1.2.1 样地设置
2019 年3 月,选取地势平坦、植被相对一致的区域开展春季(3 月 - 5 月)、夏季(6 月 - 8 月)、秋季(9 月 - 11 月)、冬季(12 月至翌年2 月)和全年连续放牧、封育6 种放牧处理,每个处理3 个重复小区。牦牛放牧,留茬高度均为5~8 cm。生长季为轮牧,植被高度20 cm 时放牧。
1.2.2 植被取样与测定
2020 年8 月上旬,在每个小区随机选取10 个50 cm × 50 cm 样方,共120 个。齐地面刈割,分种统计植物地上生物量,按功能群分为禾本科、莎草科、豆科、毛茛科、菊科和其他科。在每个样方内用根钻(直径10 cm)取1 份0 - 40 cm 土样,过筛收集根样并洗净。植被地上部分和根样放入烘箱65 ℃恒温烘48 h 后,测定地上和地下生物量,计算根冠比。
1.2.3 统计分析
试验所有数据均经过Shapiro-Wilk 检验,并满足正态分布。一般线性混合模型分析放牧对植物群落和植物功能群地上生物量、地下生物量、根冠比的影响,y= G +Γ+ε,固定效应为放牧,G是放牧,Γ是重复小区的随机效应,而ε是模型误差,显著水平P< 0.05。分析植物群落和功能群地上生物量、群落地下生物量和根冠比在不同季节牧场的变异系数,研究季节性放牧对高寒草甸牧草生物量的空间异质性。Logistics回归模型对地上生物量、地下生物量、根冠比与其分布累计频率进行拟合,并对其进行一阶、二阶导数运算,由此得出 Logistic 速度函数的3 个关键时期渐增期、快增期和缓增期[16],探求高寒草甸牧草生物量的空间分布特征。利用主成分分析(principal component analysis,PCA)研究植物功能群地上生物量与地下生物量和根冠比的关系,由多元线性回归的标准化系数得出植物功能群地上生物量对地下生物量和根冠比的贡献率。利用一般线性模型建立单个功能群地上生物量对地下生物量和根冠比的一元预测方程。采用多元线性回归模型,并逐步回归,根据VIF (variance inflation factor) < 5剔除共线性因子,建立植物功能群地上生物量对地下生物量和根冠比的多元预测方程。数据分析采用SPSS 26.0 和R 4.1.0 完成。
2 结果
2.1 高寒草甸地上和地下生物量的分配特征
2.1.1 放牧对地上生物量、地下生物量和根冠比及异质性的影响
放牧显著影响植物群落地上生物量和根冠比(P<0.001) (图1)。夏季牧场和全年连续放牧区的群落地上生物量显著低于不放牧样地、春季牧场、秋季牧场和冬季牧场。其中,不放牧和全年连续放牧区的群落地上生物量的变异系数高于其他季节牧场。根冠比在夏季牧场和全年连续放牧区高于其他样地,在冬季牧场最低。放牧降低夏季牧场、冬季牧场和全年连续放牧区群落地下生物量,但影响不显著(P>0.05),全年连续放牧区最低。不放牧样地的群落地下生物量和根冠比的变异系数最高,春季牧场最低。

图1 放牧季节对地上生物量、地下生物量及根冠比的影响Figure 1 Impacts of grazing season on aboveground biomass,belowground biomass, and root to shoot ratio
2.1.2 放牧对高寒草甸功能群地上生物量及其异质性的影响
禾本科地上生物量在秋季牧场最高,全年连续放牧区最低,前者是后者的519.62% (P< 0.05) (图2a)。春季牧场禾本科地上生物量的变异系数高于其他牧场(图2b)。放牧降低莎草科和其他科地上生物量,夏季牧场最低,分别为40.40、21.44 g·m-2,与不放牧样地差异显著(P< 0.05)。秋季牧场莎草科地上生物量的变异系数最高。毛茛科地上生物量在冬季牧场最高,在全年连续放牧区最低,后者为前者的192.69% (P< 0.05)。放牧对菊科和豆科地上生物量影响不显著(P> 0.05),但其变异系数高于其他功能区。

图2 放牧对高寒草甸功能群地上生物量的影响Figure 2 Impact of grazing on aboveground biomass on the functional groups in an alpine meadow
2.1.3 高寒草甸生物量分布特征
高寒草甸群落地上生物量109.32~725.38 g·m-2,平均322.64 g·m-2,渐增期为109.32~148.60 g·m-2,快增期为163.08~494.96 g·m-2,缓增期为558.16~725.48 g·m-2(图3)。群落地下生物量为876~3 456 g·m-2,平均1 986.72 g·m-2,渐增期为876~1 249 g·m-2,快增期为1 259~2 811 g·m-2,缓增期为2 842~3 455 g·m-2。高寒草甸根冠比为2.92~13.22,平均6.99,渐增期为2.92~3.27,快增期为3.95~10.61,缓增期为11.06~13.22。
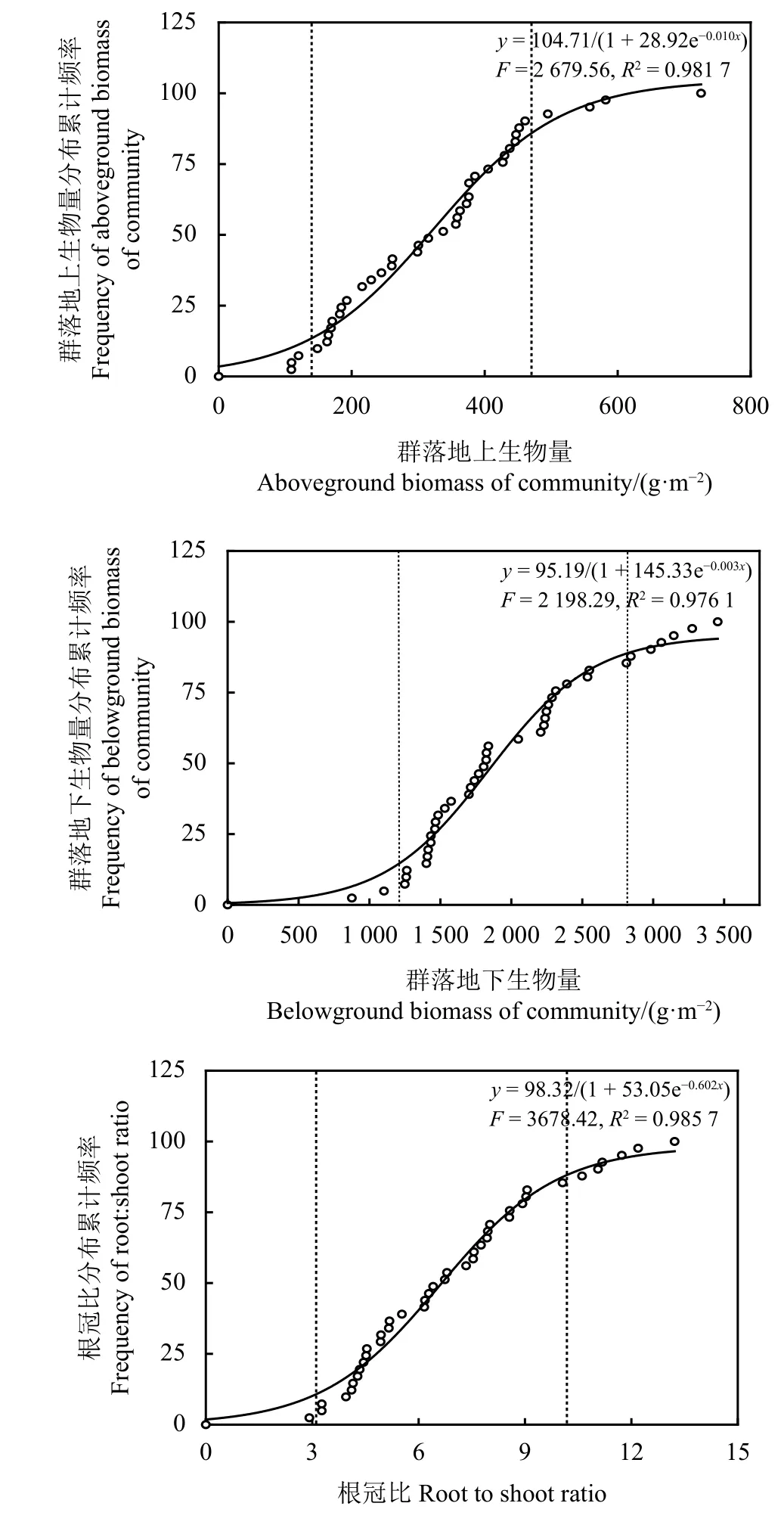
图3 高寒草甸地上生物量、地下生物量及根冠比分布Figure 3 Distributions of aboveground biomass,belowground biomass, and root/shoot ratio in an alpine meadow
2.2 高寒草甸功能组地上生物量与地下生物量和根冠比的关系及贡献
莎草科、禾本科、菊科、豆科、毛茛科和其他科地上生物量和群落地上生物量与群落地下生物量正相关,与根冠比负相关(图4)。各植被功能组与群落地上生物量的关系,毛茛科最密切,依次为毛茛科 > 菊科 > 其他科 > 禾本科 > 莎草科 > 豆科。群落地下生物量与植被功能组的关系密切程度,依次为莎草科 > 菊科 > 豆科 > 毛茛科 > 其他科 > 禾本科。群落根冠比与群落地上生物量关系最密切,关系依次为群落地上生物量 > 毛茛科 > 禾本科 > 其他科 >菊科 > 群落地下生物量 > 莎草科 > 豆科。
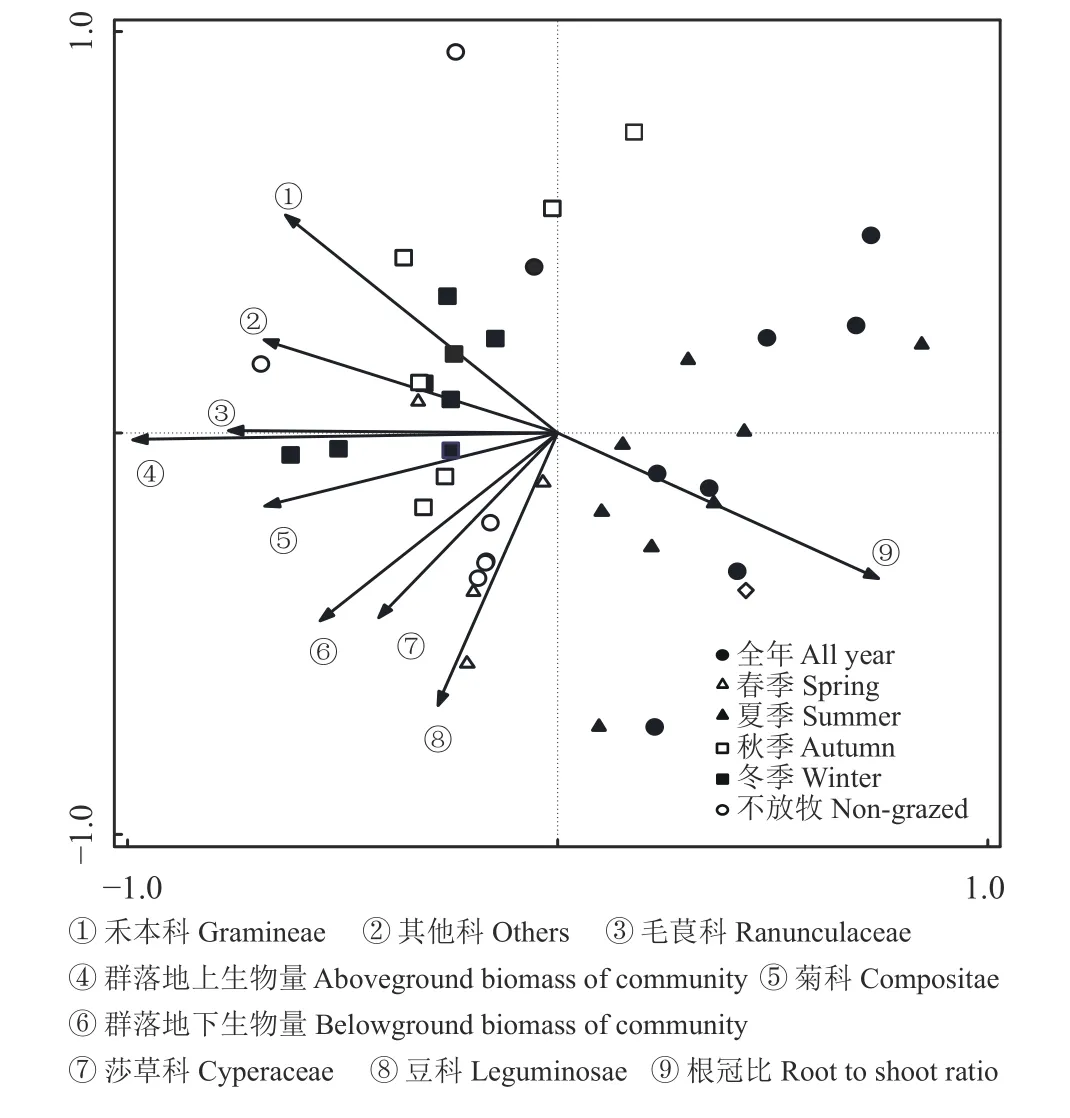
图4 高寒草甸不同功能群地上生物量与地下生物量和根冠比的主成分分析Figure 4 Principal component analysis of aboveground biomass of functional groups, belowground biomass, and root to shoot ratio in an alpine meadow
菊科地上生物量对群落地上生物量贡献率最大,豆科地上生物量贡献率最小(表1)。其他科、莎草科和菊科地上生物量对群落地下生物量贡献率较大,豆科地上生物量贡献率最小。毛茛科和禾本科地上生物量对根冠比的贡献率最大,其他科的贡献率最小。

表1 高寒草甸不同功能群地上生物量对地下生物量和根冠比的贡献Table 1 Contributions of aboveground biomass of functional groups to belowground biomass and root to shoot ratio in an alpine meadow
2.3 预测模型
2.3.1 功能群地上生物量与地下生物量和根冠比一元预测模型
高寒草甸,禾本科、菊科、其他科和莎草科牧草地上生物量与群落地下生物量均显著正相关(P<0.05),而毛茛科和豆科地上生物量与群落地下生物量相关性不显著(P> 0.05) (图5)。

图5 禾本科、莎草科、豆科、毛茛科、菊科和其他科地上生物量与总地下生物量的回归关系Figure 5 Regression relationships between aboveground biomass of Gramineae, Cyperaceae, Leguminosae,Ranunculaceae, Compositae, others and total belowground biomass
除豆科外,禾本科、毛茛科、菊科和其他科地上生物量均与根冠比极显著负相关(P< 0.01) (图6)。
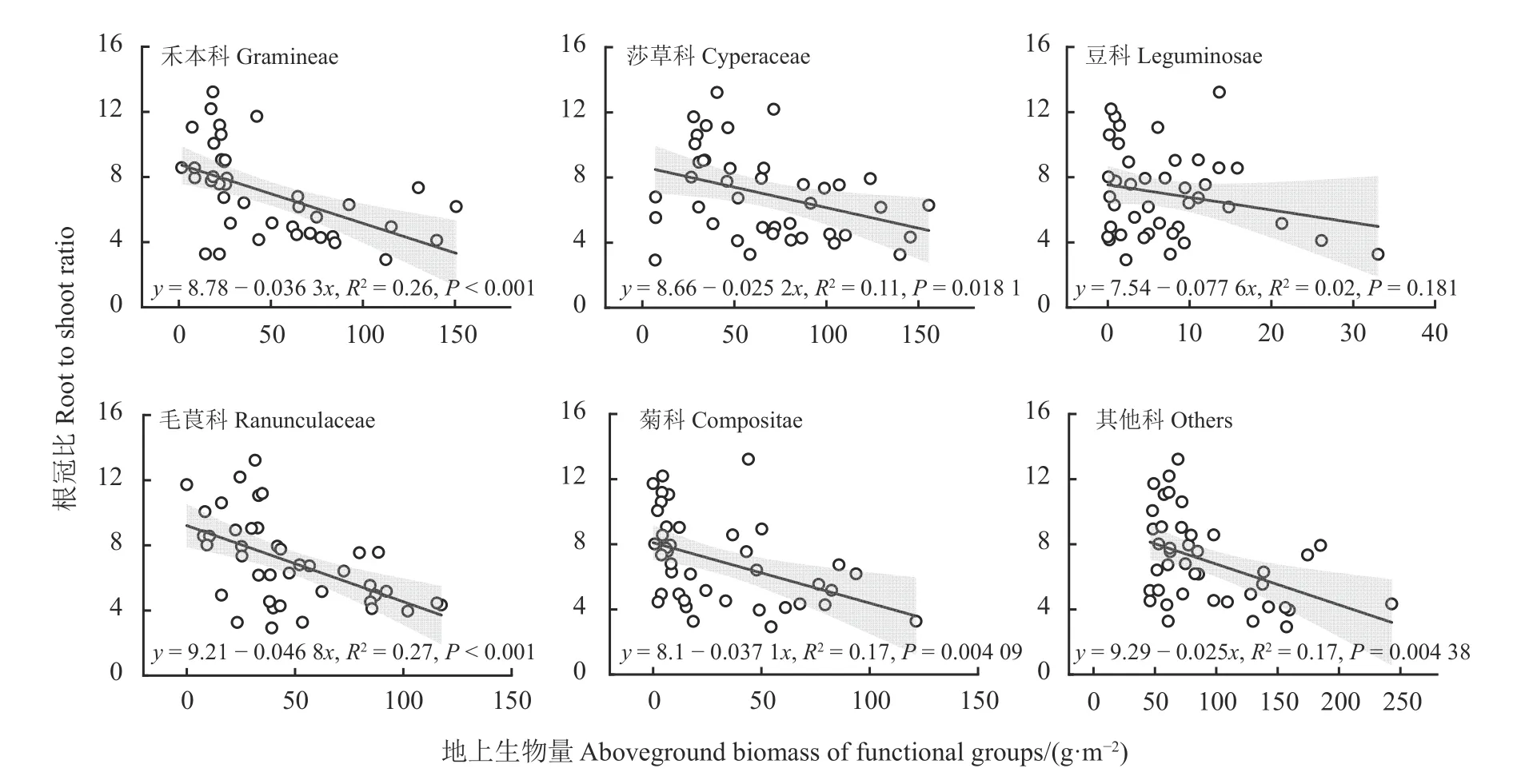
图6 禾本科、莎草科、豆科、毛茛科、菊科和其他科地上生物量与根冠比的回归关系Figure 6 Regression relationships between aboveground biomass of Gramineae, Cyperaceae, Leguminosae,Ranunculaceae, Compositae, others and root to shoot ratio
2.3.2 地上与地下生物量和根冠比多元预测模型
其他科和菊科地上生物量是群落地下生物量的重要预测因子,禾本科和毛茛科地上生物量是根冠比的重要预测因子(地下生物量 = 8.343 × 其他科 +6.104 × 菊科 + 354.194,R2= 0.275*,F= 6.94,P其他科=0.007,P菊科= 0.038;根冠比 = -0.111 × 禾本科 - 0.145 ×毛茛科 + 10.093,R2= 0.401**,F= 15.11,P其他科= 0.003,P菊科= 0.002),故分别选取群落地下生物量和根冠比进行预测模型的建立。
3 讨论
青藏高原高寒草甸群落地上生物量、地下生物量及根冠比的分布符合Logistic 曲线分布。秋季、冬季牧场因试验当年未经过家畜采食,导致群落地上生 物 量 在109.32~148.60 和139.54~181.37 g·m-2,变异率低;而全年连续放牧区家畜的采食导致群落地上生物量在163.08~494.96 g·m-2,变异率较高。全年连续放牧区内放牧强度高于季节性放牧牧场,家畜的选择性采食导致群落呈现斑块化,植被异质性较高[17];不放牧样地群落地上生物量异质性较强,可能因为原样地为过度放牧区域,原有植被群落由于长期过度放牧而向以毒害草和裸地为主的“黑土滩”演替,植被群落呈典型斑块分布[18]。虽经短期围封草地演替方向改变[19],但禾本科与莎草科植物在毒害草斑块分布区域难以迅速占据优势地位,导致群落生物量较低;而在毒害草分布较少的区域则能迅速生长,导致群落生物量分布具有较高变异率。研究区豆科植物以毒害草披针叶黄华(Thermopsislanceolala)与甘肃棘豆(Oxytropis kansuensis)为主,此类植被在不放牧情况下受资源和空间限制,空间格局呈聚集分布,导致豆科植物在不放牧样地中具有较高的异质性[20]。夏季与全年连续放牧区群落地下生物量较低,导致群落地下生物量在1 259~2 811 g·m-2内数据的升高。冬季牧场根冠比较低,根冠比较多分布于3.95~10.61。不放牧样地的群落地下生物量和根冠比的变异系数最高,春季牧场最低。不放牧样地由于试验开始前的人为干扰导致群落演替方向变化,群落地下生物量与根冠比分布受原有植被条件影响而变化,变异系数高。春季放牧早于生长季,导致牧草再生长速率变慢呈现均一化趋势[21],群落地上生物量趋于一致,变异系数最低。
放牧季节对高寒草甸群落地上生物量影响显著,其中夏季和全年连续放牧区群落地上生物量显著低于其他牧场,这是由于牦牛的采食作用强于植物的补偿生长作用。植物地下生物量相对地上生物量对放牧季节响应不敏感,可能因为地下生物量对放牧的响应具有时滞效应[22],因而两年的放牧尚未对其造成显著影响。同时,地下生物量会受气候条件、放牧强度、放牧时间、土壤条件等诸多因素影响[23]。通常,不合理放牧抑制地上部分生长,也会减少地上部分向地下的物质分配,在一定程度上降低牧草地下生物量[24],适度放牧则促进地下部分生长[25]。由于本研究中季节性轮牧的放牧率较低,因此对地下生物量的影响不显著。较低的根冠比通常代表着植物光竞争能力强,而较高的根冠比则代表植物对土壤元素利用能力的增强[26]。放牧季节对根冠比的影响显著,夏季和全年连续放牧区根冠比显著高于其他牧场,这可能由于牦牛采食牧草光合器官削弱了冠层对资源的竞争能力,对地下生物量的影响较小,因而植物对土壤元素利用能力强[27];冬季牦牛放牧促进植物生长,降低了冬季牧场牧草的根冠比。
生物量反映植被对资源的竞争能力[28]。禾本科、毛茛科、莎草科植物对群落地上生物量贡献率较高,由于3 个功能群竞争力更强,具有较高的生物量占比,为高寒草甸优势功能群[29]。此外,禾本科植物主要为披碱草属(Elymus)、早熟禾属(Poa)和剪股颖属(Agrostis)植物,处于结实期,地上生物量较高[30],而且小花草玉梅(Anemone rivularis)等毛茛科植物地上部分较为发达。莎草科、菊科和其他科植物地上生物量对地下生物量作用较大,其他科植物主要为甘肃马先蒿(Pedicularis kansuensis)、鹅绒委陵菜(Potentilla anserina)、椭 圆 叶 花 锚(Halenia elliptica)。一方面,可能是因为甘肃马先蒿具有较高的地下生物量和根冠比[31];鹅绒委陵菜根系较为发达,且具膨大的地下贮存块根;椭圆叶花锚则为直根系植物,也具有较高的地下生物量。另一方面,可能由于家畜较少直接采食毒草或贴地生长的植物,这些植物有较多机会合成光合产物并向下运输[32],且毒草促使家畜避食其他植物,群落生长受放牧影响较小[33]。
莎草科植物为须根系,根系发达且分蘖结较多,地下生物量较高[34]。菊科主要为黄帚橐吾(Ligularia virgaurea),具有粗壮的须根根系和发达的横走根状茎[35]。草甸植被群落中,莎草科植物具有发达的根系和较多的分蘖结,地下生物量也较高[34],并可能通过发达的根系在浅层土壤中形成密集的草毡层[36],从而降低其他植物类群在浅层土壤中所能获得的养分与水分,并迫使其他植物根系生长至更深处土壤获取养分,促使牧草生物量向地下分配。此外,莎草科植物作为草甸建群种,其地上生物量与群落地下生物量相关性较高。禾本科和毛茛科植物由于具有较高的地上生物量[30],根冠比相对较低,且二者均为优势种,降低了高寒草甸群落根冠比,因而二者根冠比可能更接近高寒草甸群落根冠比,对根冠比贡献较高,故选用二者进行根冠比的预测。高寒草甸生态系统中,选用禾本科、毛茛科、其他科和莎草科牧草地上生物量预测群落地下生物量和根冠比,豆科生物量较少,故其地上生物量与群落地下生物量和根冠比无关联。莎草科和群落地下生物量较强的关联性可能是由于玛曲高寒草甸中莎草科在群落中优势度高,对群落地下生物量贡献率较高,根冠比也较高,从而能够提高群落根冠比[37],即根冠比更高的植物类群与较高的群落地下生物量之间有较强的相关性。
对高寒草甸各植物类群与群落地下生物量、根冠比的相关性分析表明,这些指标与多个植物类群的地上生物量相关。对于地下生物量的预测,由于菊科、其他科根系发达,根冠比较高,对群落地下生物量贡献率高,为群落地下生物量的重要预测参数;而在对群落根冠比的预测中,禾本科与毛茛科同样由于其较高的根冠比,对群落根冠比具有较高的贡献率,对群落根冠比的预测具有重要意义。综上,建立一个通过多种植物类群预测高寒草甸群落地下生物量与根冠比的多元预测模型,可以更加准确地通过地上群落推测地下生物量,并有更广泛的适用性,从而为地下生物量的测定提供便利。
4 结论
青藏高原高寒草甸地上生物量、地下生物量及根冠比的分布呈近似的正态分布。高寒草甸地上生物量为109.32~725.38 g·m-2,快增期为163.08~494.96 g·m-2。地下生物量为876~3 456 g·m-2,快增期为1 259~2 811 g·m-2。高寒草甸根冠比为2.92~13.22,快增期为3.95~10.61。快增期数据变化较快,对放牧管理措施具有重要意义,需通过放牧管理调控。季节性放牧对高寒草甸地上生物量、地下生物量及根冠比影响显著,夏季放牧和全年连续放牧区的地上生物量显著低于其余放牧季节,地下生物量在夏季牧场、全年连续放牧区和冬季牧场低于其他样地,根冠比在夏季牧场和全年连续放牧区高于其他样地,在冬季牧场最低。其中,菊科对地上生物量贡献率最大,豆科贡献率最小;其他科、莎草科和菊科对地下生物量贡献率较大,豆科贡献率最小;毛茛科和禾本科对根冠比的贡献率最大,其他科的贡献率最小。莎草科地上生物量可以在一定程度上反映群落总体地下生物量水平。高寒草甸地下生物量可以通过菊科和其他科地上生物量构建多元预测模型进行计算,根冠比则可以通过禾本科、毛茛科地上生物量构建多元预测模型计算。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
农业工程学报(2022年10期)2022-08-22
林业调查规划(2022年3期)2022-06-13
海外文摘·文学版(2021年3期)2021-05-19
安徽农学通报(2020年7期)2020-05-26
西部资源(2018年3期)2018-11-06
中国中药杂志(2017年17期)2017-09-23
延河·绿色文学(2016年8期)2016-05-14
农产品市场周刊(2015年39期)2015-10-31
现代农业科技(2009年9期)2009-02-21
