
草地利用方式对呼伦贝尔草地微生物群落结构的影响
2022-03-26 02:15王德平刘忠宽刘玉玲戎郁萍
草业科学 2022年3期
王德平,宋 倩,刘忠宽,赵 坤,刘玉玲,肖 红,曹 婧,戎郁萍
(1.中国农业大学草业科学与技术学院, 北京 100193;2.河北省农林科学院农业资源环境研究所, 河北 石家庄 050051)
呼伦贝尔草地是我国北方重要的生态屏障与草牧业生产基地,随着全球气候变化及人为不合理利用,该地区80%的草地出现不同程度的退化,生物多样性与稳定性降低,草地生产力下降,生态系统结构和功能受损[1]。土壤微生物是维持草地生态系统物质循环和能量流动的关键因子[2],对环境的变化较为敏感,对土壤健康和草地退化具有较好的指示作用[3]。土壤微生物通过分解有机质,为土壤养分循环提供动力;并且通过菌丝缠绕、分泌有机酸及黏结作用促进大团聚体的形成,在改良土壤质量方面发挥关键作用[3];同时土壤微生物通过与植被进行碳交换,在调节植物生长、发育中也发挥重要作用[4-5]。一些研究认为土壤微生物的多样性及复杂性在预测草地群落的多样性及稳定性方面具有关键指导作用[6];同时土壤微生物群落结构的多样性有利于提高土壤对环境变化的缓冲能力,进而保持草地生态系统功能的稳定[7]。
放牧是天然草地的普遍利用方式,呼伦贝尔草地产草量高,为满足畜牧业需要,一部分天然草地将会被刈割,并加工处理为饲草料,因此,刈割是除了放牧之外的草地主要利用方式;而开垦为农田有利于获得更大的经济效益[8]。因此,草地刈割、放牧、开垦成农田等是呼伦贝尔地区主要草地利用方式,草地利用方式的转变可以通过影响土壤理化性状,进一步改变土壤微生物生物量和群落结构[9]。方圆等[10]对北方温带草原研究发现,草甸草原土壤微生物群落主要受土壤全碳(total carbon,TC)、全氮(total nitrogen,TN)调节,宋倩[11]在呼伦贝尔草原的研究也发现TC、TN 与细菌、真菌磷脂脂肪酸(phospholipid fatty acid, PLFA)量呈正向关系。放牧通过采食践踏及粪尿归还,影响土壤微生物生物量及组成。Zhao 等[12]发现放牧显著降低土壤微生物总生物量,但不同微生物群落对放牧的响应存在差异;Xu 等[13]在亚热带放牧场的研究发现,放牧显著提高土壤微生物生物量,但细菌丰度增加而真菌丰度降低,这主要是由于真菌比细菌对复杂底物具有更高的利用效率,而细菌更利于分解简单底物[14];同时,放牧作用促进高质量凋落物(低碳氮比)输入土壤中,提高土壤养分状况从而降低真菌丰度。基于内蒙古草地的meta 分析发现[15],重度放牧显著降低土壤有机碳,轻度放牧显著增加了土壤有机碳,而土壤有机碳与微生物生物量呈显著正相关关系,土壤微生物群落对放牧的响应方式及其与土壤理化性状的关系仍需进一步研究。围封是退化草地常用的恢复措施,围封通过提高地上生物量和根系分泌物,有利于土壤微生物繁殖[16]。但是,长期围封导致立枯物和凋落物分解缓慢,植被绿色活体生物量降低,进而影响养分循环,从而抑制土壤微生物对养分的获取[17],因此,围封可能在短期内提高土壤微生物生物量,但长期围封可能对微生物群落起抑制作用。刈割通过地上植被的移除,改变地上植被的丰富度和多样性,也影响微生物群落结构[18]。有学者研究发现刈割导致植物光合器官损伤和消耗,使土壤微生物可获得的养分及凋落物减少,导致微生物生物量降低[19]。在高草草原的研究中发现,适度刈割通过减少遮阴增加光合作用、补偿生长和改变分配模式[20];有利于地上、地下生物量的积累,促进微生物群落的繁殖。草地开垦为农田使地上生物多样性显著降低,同时人为的田间管理(施肥、灌溉、翻耕等)对土壤的扰动很大[21],土壤微生物生物量及群落结构对草地转变为农田的响应极为复杂。施肥是农田管理的重要措施,主要包括矿物肥料和有机肥料。矿物肥料可能通过影响土壤的非生物特性(如酸碱度)[22],强烈影响土壤微生物生物量和微生物群落的组成和功能。与矿物肥料相比,有机肥料的应用导致了土壤微生物群落结构和功能的根本不同,并导致土壤微生物生物多样性的减少[23]。农田翻耕降低了土壤团聚体的稳定性,加速了微生物对团聚体中碳的利用[24];对81 个草地开垦的案例进行meta 分析时发现,草地向农业用地的转变普遍造成了土壤有机碳的流失,但不同地区抵御土壤有机碳流失存在显著差异[25],具体的作用机制及调控策略尚不明确。
本研究以围封草地为对照,通过调查不同草地利用方式(刈割、放牧、开垦)下土壤理化性状和磷脂脂肪酸含量,研究草地利用方式对土壤微生物群落结构的影响,以期为呼伦贝尔地区草地合理利用和退化草地修复提供理论依据。
1 材料与方法
1.1 试验区概况
试验地位于内蒙古呼伦贝尔市陈巴尔虎旗特尼河九队(49°20′~49°26′ N,119°55′~120°09′ E,海拔629~653 m),温带大陆性季风气候,年均温-1.1 ℃,7 月为最热月,月均温在20~22 ℃,1 月为最冷月,月均温为-28~-25 ℃;无霜期100~115 d。年降水量约345 mm,其中6 月 - 9 月降水量约占全年降水量的80%。土壤类型为黑钙土,土壤表层有机碳含量约36.7 g·kg-1。
试验区草地为温性草甸草原,建群种为羊草(Leymus chinensis),伴生植物有贝加尔针茅(Stipa baicalensis)、无芒雀麦(Bromzus inermis)、裂叶蒿(Artemisia tanacetifolia)、 星 毛 委 陵 菜 (Potentilla acaulis)、寸草苔(Carex duriuscula)、日荫菅(Carex pediformis)、 黄 花 蒿(Artemisia annua)、 斜 茎 黄 芪(Astragalus adsurgens)等。
1.2 野外调查方法
2019 年8 月中旬进行野外调查取样,选取空间上相近的3 种草地(刈割、放牧、开垦)和围封草地;其中,围封样地自2016 年开始围封;刈割样地自2000 年起,每年8 月中、下旬刈割,留茬8 cm;放牧样地自2013 年起,每年6 月 - 10 月初放牧,其余时间休牧,载畜率为每年每公顷0.43 标准牛单位;草地开垦转化为农田样地已耕种60 年,种植作物为小麦(Triticum aestivum)。在上述样地中,每类样地选取具有代表性的地段,设置3 块100 m × 300 m 的子样地,共计12 块子样地。在每块子样地中采用“S”型取样法,随机选取10 个样点,在每个样点处随机选取一个50 cm × 50 cm 的样方,使用直径5 cm 的土钻采集0 - 10 cm 土壤,将每个子样地所取的10 个样方土壤样品混合为3 个土壤样品;所有样地共计获得36 个土壤样品。将所得土壤样品去除石砾、根系及杂物,过2 mm 筛;将剩余土壤样品分为3 份,一份置于-20 ℃冰箱用于测定土壤铵、硝态氮,一份置于-80 ℃冰箱,用于测定土壤微生物PLFA 含量;一份自然阴干后用于土壤基础养分的测定。
1.3 土壤样品测定

表1 特定微生物磷脂脂肪酸标记Table 1 Labeling of specific microbial phospholipid fatty acids
1.4 数据分析
对土壤理化性状、磷脂脂肪酸类型、微生物各类群PLFA 含量进行单因素方差分析及Duncan 差异显著性检验(α = 0.05)。使用Vegan 包中的decorana、rda 函数对土壤理化性质与微生物群落结构进行冗余分析,用Ezekiel 公式进行R2校正,采用蒙特卡洛置换检验进行约束轴的置换检验,置换次数为999 次。数据在R3.6.0 中分析,Sigmaplot 14.0 作图。
2 结果与分析
2.1 草地利用方式对土壤理化性状的影响
除铵态氮含量外,其他土壤指标4 种草地利用方式间均出现显著差异(P< 0.05) (表2)。草地开垦为农田土壤TC、TN 显著低于其他草地利用方式(放牧、围封、刈割);但开垦处理下AP、EC 及pH 均显著高于其他3 种草地利用方式(放牧、围封、刈割);与放牧、围封、刈割草地相比,农田土壤AP 分别提高了327.39%、371.93%和395.23%,EC 分别提高了149.49%、139.40%和102.30%。4 种草地利用方式中,仅农田土壤pH 表现为碱性,其余土壤pH均表现为酸性。与围封相比,放牧处理TP、硝态氮均显著降低,但AP、TC、TN 与围封并无显著差异。围封处理硝态氮显著高于开垦、刈割、放牧,硝态氮含量分别提高了26.25%、67.07%和20.08%。

表2 草地利用方式对土壤养分含量的影响Table 2 Effects of grassland utilization on soil nutrient content
2.2 草地利用方式对微生物群落结构的影响
土壤样品中检测出各处理共有的PLFA 标记磷脂脂肪酸26 种(图1)。其中,支链饱和脂肪酸(branched-chain saturated fatty acids, BFA) 10 种:13 ∶ 0 iso、14 ∶ 0 iso、15 ∶ 0 anteiso、15 ∶ 0 iso、16 ∶ 0 iso、16 ∶ 0 10-methyl、17 ∶ 0 iso、17 ∶ 0 anteiso、18 ∶ 0 iso、20 ∶ 0 10-methyl,BFA 相对含量占土壤磷脂脂肪酸总量的37.84%~42.71%。共检测出单烯脂肪酸(monoenoic fatty acids,MFA) 6 种:16 ∶ 1 w5c、16 ∶ 1 w9c、16 ∶ 1 w7c、18 ∶ 1 w7c、18 ∶ 1 w5c、18 ∶ 1 w9c,MFA 相对含量占PLFA 总量25.47%~29.95%。共检测出6 种直链饱和脂肪酸(straight chain saturated fatty acids, SFA):14 ∶ 00、15 ∶ 00、16 ∶ 00、17 ∶ 00、18 ∶ 00、20 ∶ 00,SFA 相对含量占PLFA总量19.96%~23.69%。多烯脂肪酸(polyenoic fatty acids, PFA) 2 种:18 ∶ 3 w6c、18 ∶ 2 w6c;相对含量均在5%以下。环丙烷脂肪酸(cyclopropane fatty acids,CFA) 2 种:17 ∶ 0 cyclo w7c、19 ∶ 0 cyclo w7c,相对含量在5.45%~6.82%。与放牧草地和围封草地相比,刈割草地和农田BFA 显著降低(P< 0.05),但MFA显著增高;农田SFA 显著高于刈割草地,但与放牧草地、围封草地无显著差异(P> 0.05);刈割草地PFA 显著高于其他草地利用方式。在各草地利用方式中BFA 占比均为最高;且土壤磷脂脂肪酸类型占比均为BFA > MFA > SFA > CFA > PFA (表3)。

表3 不同草地利用方式下土壤磷脂脂肪酸类型及其比例Table 3 PLFA and their proportions under different grassland utilization methods
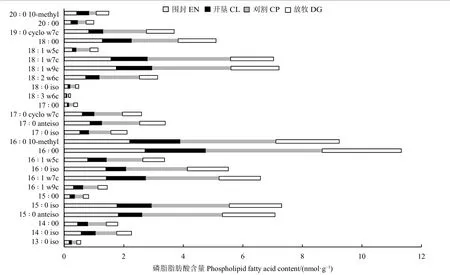
图1 不同草地利用方式土壤微生物PLFA 图谱Figure 1 Soil microbial PLFA profiles map under different grassland utilization methods
草地利用方式对各微生物菌群均具有明显影响(图2)。除AMF 外,刈割草地各微生物菌群均显著高于农田、围封草地和放牧草地(P <0.05)。其中,细菌、放线菌、G+、总PLFA 菌群在农田、围封草地和放牧草地间无显著差异(P> 0.05),围封草地真菌显著高于农田。与刈割草地相比,农田G+显著降低,放牧草地G+与围封草地无显著差异。各微生物菌群均表现为刈割草地最高,且在4 种草地利用中细菌占总PLFA 比例均为最高。

图2 草地利用方式对微生物群落的影响Figure 2 Effects of grassland utilization on microbial community
不同草地利用方式细菌/真菌PLFA 浓度比值在各处理下均无显著差异(P> 0.05)。细菌/真菌PLFA浓度比值的变异系数在0.01~0.10,变异系数表现为围封 = 放牧 > 农田 > 刈割。草地利用方式对G+/G-影响明显,与围封草地相比,草地刈割和农田显著降低了G+/G-;放牧处理G+/G-低于围封,但二者无显著差异(P> 0.05)。G+/G-变异系数在0.03~0.09,具体表现为开垦 > 放牧 > 刈割 > 围封;开垦处理G+/G-最小,但变异系数最大(表4)。

表4 草地利用方式对土壤微生物PLFA 比值的影响Table 4 The influence of grassland utilization on the ratio of soil microbial PLFA
2.3 土壤理化性状与微生物群落的冗余分析
RDA1 轴和RDA2 轴共同解释微生物群落组成变异的51.04% (图3)。土壤AP、pH、EC 与G+、细菌、总PLFA、真菌呈显著负相关关系(P< 0.05);TC、TN 与G+、细菌、总PLFA、真菌呈显著正相关关系;AMF、G-、放线菌与AP、pH、EC、TC、TN 的相关性均较低,但总体仍与TC、TN 呈正相关关系,与AP、pH、EC 呈负相关关系。
3 讨 论
3.1 草地利用方式对土壤理化性状的影响
草地开垦为农田易导致土壤C、N 元素流失,本研究发现草地开垦为农田,土壤TC、TN 分别降低41.47%和45.08%。地上凋落物与植物根系是土壤C、N 的重要来源,与草地植被相比,农作物地下根系并不发达,且大多数农作物根系以浅层须根为主[27],由根系分解至土壤的C、N 相对较少。除此之外,研究区农田种植作物为小麦,地上生物量较大,农田收获导致大量的地上植被从农田生态系统中移除;因此,与其他3 种草地利用方式相比,农田生态系统由地上植被返还土壤的C、N 也相对较少。同时,草地开垦导致土壤透气性增加,提高了微生物对有机碳的矿化速率[28],且开垦导致的地表裸露,有利于风蚀对土壤团聚体的破坏,直接导致土壤团聚体中的微团聚体流失。草地转变为农田后,土壤有机碳由主要大团聚体储存,转变为主要由微团聚体储存[29],因此,微团聚体的损失会进一步加速土壤C、N 流失。除此之外,风蚀对土壤团聚体的物理破碎作用有利于提高微生物与有机质的接触面积[30],这可能是造成草地开垦后土壤C、N 损失的另一原因。土壤开垦后,地表水分蒸发较高,地下盐分随水分运移向土壤上层积累,进而导致草地开垦后pH增加[31]。同时,农田施肥导致速效养分及盐分归还至土壤,直接导致土壤速效磷及电导率的提高。
与C、N 元素不同,P 元素易被植物吸收利用[32-33]。放牧通过家畜采食作用造成植被损失,进而影响植物将更多的磷吸收储藏,而未被分解归还至土壤[34],这可能是导致放牧草地土壤全磷显著低于围封草地的主要原因。研究区土壤速效氮主要以硝态氮形式存在,铵态氮含量较少;放牧对硝态氮的影响可能取决于放牧强度[35],Zhou 等[36]在全球尺度的meta分析表明,中、重度放牧强度导致土壤速效氮流失,而轻度放牧对土壤速效氮影响较小。首先,中、重度放牧下家畜的选择性采食导致草地群落组成改变[37],进而造成进入土壤的地上凋落物质量降低[38],从而降低土壤N 库。其次,家畜践踏导致土壤紧实,放牧草地氧分压相对较低,适宜反硝化细菌生存,且家畜粪便中C/N 比较低[39],粪便的返还为反硝化细菌提供N 源及易分解的有机质,进而造成反硝化作用增强,导致放牧草地硝态氮的损失。
3.2 草地利用方式对微生物群落的影响
在4 种草地利用方式中,土壤微生物均以细菌为主体,这与在青藏高原[40]的研究结果一致。谭红妍[41]在对呼伦贝尔草地的研究中发现,刈割提高了各菌群PLFA 量,这与本研究结果一致。冗余分析表明,TC、TN 与各微生物类群呈不同程度的正相关关系,土壤微生物碳、氮主要来源于土壤碳、氮[42],刈割处理中土壤TC、TN 在各个处理中均为最高,这在一定程度上解释了刈割导致各菌群PLFA 量升高。除此之外,适度的刈割促进地上植物生长,并进一步提升养分周转效率,这有利于促进地下共生微生物的繁殖,进而导致微生物PLFA 量增加[43]。同时AP、pH 和EC 与各微生物类群PLFA 量均呈负相关关系,Wen 等[44]研究认为,N 沉降背景下,过多的N 素进入土壤,可造成土壤速效磷增加,导致微生物底物酸化及木质素和纤维素降解能力的减弱等,进一步造成微生物生物量减少[45]。
与刈割不同,草地开垦后地上凋落物减少且土壤有机质流失严重[46],而真菌在分解凋落物及通过菌丝缠绕作用形成大团聚体中发挥关键作用[47],可能是开垦处理下,真菌PLFA 量显著降低的原因。与革兰氏阴性菌相比,革兰氏阳性菌分解复杂碳组分的能力更为突出[48],本研究中农田G+/G-显著降低(P< 0.05),表明农田土壤微生物群落向分解高活性碳底物方向转变。这可能是草地开垦后,由于人为输入碳底物,农田生态系统中易分解碳的比例增加,不利于主要分解复杂碳组分的革兰氏阳性菌生存。同时,草地开垦后,人为施肥、水分蒸发、土壤风蚀等原因导致农田生态系统较高的土壤EC、AP 和pH,EC、AP、pH 与土壤各微生物类群均成负相关关系(图3),这表明农田较高的EC、AP 和pH一定程度上抑制土壤各微生物类群的生长和繁殖。可能是由于人为施肥导致的P 添加,加剧了土壤微生物的C、N 限制;Xiao 等[49]在呼伦贝尔草地的N、P 添加试验也发现,P 添加增加了微生物的N 限制,N 添加增加了微生物的C 限制。因此土壤TC、TN 可被考虑作为草地开垦为农田后微生物群落的重要调控因素。
4 结 论
呼伦贝尔草地开垦为农田导致土壤C、N 流失,降低革兰氏阳性菌和真菌PLFA;且开垦处理下,土壤电导率、pH 及速效磷的提高不利于微生物生长、繁殖。刈割通过提高土壤TC、TN,降低AP 等方式为微生物各菌群生长提供有利土壤理化条件。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
当代水产(2022年8期)2022-09-20
农业工程学报(2022年10期)2022-08-22
中国音乐学(2022年2期)2022-08-10
浙江农林大学学报(2022年2期)2022-04-08
安徽农学通报(2020年7期)2020-05-26
学校教育研究(2020年7期)2020-04-09
大自然探索(2019年1期)2019-01-24
知识就是力量(2017年12期)2018-01-15
中学生理科应试(2017年6期)2017-09-27
