
次生天然林不同恢复阶段土壤硝化、反硝化功能微生物特征及影响因素分析
2022-04-27 12:56张宇杨军钱
福建农业科技 2022年2期
张宇 杨军钱





摘 要:為了解次生林自然恢复过程中硝化、反硝化微生物功能基因丰度变化规律及影响因素。采用空间代替时间的方法,选取环境条件基本一致的5、8、21、27、40年自然恢复的次生林和大于100年原始林(对照CK)为对象,研究了表层(0~10 cm)土壤中参与硝化过程的功能基因AOA、AOB和反硝化功能基因NarG、NirK、NirS和NosZ丰度变化,以及与土壤理化性质的关系。结果表明:AOA基因主导了恢复阶段土壤的硝化过程,pH和铵态氮含量是影响AOA基因丰度变化的因子,进一步支持了AOA适合低pH和较低浓度NH4+的观点。反硝化功能基因丰度主要受AOB基因和土壤C/N的影响,且(NirK+ NirS)/NosZ丰度比小于1,表明自然恢复的次生林能够更好地将氮素保持在土壤中,减少土壤中N2O的排放。随着次生林自然恢复时间推移,氮碳养分的逐渐积累,为微生物活动提供了丰富的底物和能量来源,微生物活动也逐步恢复。以上结果表明,自然恢复的次生林能够更好保持土壤肥力,并且具备更高的生态系统功能。
关键词:次生天然林;硝化过程;反硝化过程;功能基因丰度;自然恢复
中图分类号:S 792.155 文献标志码:A 文章编号:0253-2301(2022)02-0063-08
DOI: 10.13651/j.cnki.fjnykj.2022.02.012
Microbial Characteristics and Influencing Factors of Soil Nitrification and Denitrification Microorganismsin the Secondary Natural Forests at Different Restoration Stages
ZHANG Yu1,2, YANG Jun-qian1,2*
(1. Fujian Normal University, Fuzhou, Fujian 350007, China; 2. State Key Laboratory Breeding Baseof Humid Subtropical Mountain Ecology, Fuzhou, Fujian 350007, China)
Abstract: In order to understand the changes and influencing factors of the functional gene abundance of nitrification and denitrification microorganisms during the natural recovery process of secondary forests, by using the method of space instead of time, 5, 8, 21, 27, 40 years of natural recovering secondary forest and more than 100 years of primeval forest ( the control group CK) were selected as the research objects, and then the abundance changes of the functional genes such as AOA, AOB and the denitrification functional genes such as NarG, NirK, NirS and NosZ genes which were involved in the nitrification process in the surface soil (0-10 cm) and their relationships with the soil physical and chemical properties were studied. The results showed that AOA gene dominated the nitrification process in the soil at the recovery stage, and the pH and ammonium nitrogen content were the factors affecting the abundance change of AOA gene, which further supported the view that AOA was suitable for low pH and low concentration of NH4+. The abundance of the denitrification functional genes was mainly affected by AOB gene and the soil C/N, and the abundance ratio of (NirK+NirS)/NosZ was less than 1, indicating that the natural recovering secondary forest could better maintain the nitrogen in the soil and reduce the emission of N2O in the soil. With the natural recovering time of secondary forest, the gradual accumulation of nitrogen and carbon nutrients provided abundant substrates and energy sources for the microbial activities, and the microbial activities were also gradually recovered. The above results showed that the natural recovering secondary forest could better maintain the soil fertility and had higher ecosystem functions.
Key words: Secondary natural forest; Nitrification process; Denitrification process; Abundance of functional genes; Natural recovery
氮循环是地球生态系统元素循环的核心之一,在不同形式氮的转化中起着重要作用[1-2],氮素不仅是土壤的生产力和可持续发展的重要指标,氮素流失还会影响生态系统平衡[3]。微生物作为氮循环的主要驱动因素,在不同的氮素转化过程中发挥着重要作用,如固氮、硝化、反硝化、厌氧氨氧化等。不同的微生物群落及其功能基因(例如用于硝化的amoA,以及用于反硝化的NarG、NirK、NirS和NosZ)参与不同的氮转化过程[4]。硝化、反硝化功能基因丰度常作为预测和评估潜在的硝化、反硝化速率的重要指标,研究表明反硝化功能基因的动态变化可以为森林土壤反硝化潜势提供生物学指标[5]。因此了解参与氮循环的功能基因丰度的差异及影响因素,有助于进一步探索和解释氮循环的微生物机制。
硝化过程作为氮循环的中心和限速环节,其主要是将土壤中的铵态氮氧化成硝态氮,硝化过程主要的功能基因是氨氧化细菌AOB和氨氧化古菌AOA[4];反硝化过程是指反硝化微生物在氧气缺乏的状态下以NO3-为电子受体,以有机碳为电子供体,将硝态氮逐步还原为N2O或N2[2]。参与反硝化功能基因包括编码硝酸盐还原酶的NarG,编码亚硝酸盐还原酶的NirK和NirS,编码一氧化氮还原酶的NorB和编码氧化亚氮还原酶的nosZ[4]。
我国日益增长的生产需要导致森林破坏和面积减少的同时,也带来了一系列的环境问题。如何保证森林恢复的同时减少环境问题成为国家和生态学者的研究重点[6]。人工造林和自然恢复两种模式是目前世界上森林恢复主流方式,现有研究发现,自然恢复的天然林可以恢复退化的土壤特性并提高生态系统功能[7]。氮循环作为森林重要生态系统功能之一是否会随森林破坏恢复发生变化,参与硝化、反硝化特定功能基因丰度是否随之改变?目前对于这方面特别是次生天然林的研究较少,对其认识不够清楚。鉴于此,本研究通过空间替代时间的方法,选取了不同恢复阶段(5、8、21、27、40年)的次生林作为研究对象,并选取当地林龄大于100年、历史上未曾遭受人类活动影响原始林作为对照,研究天然次生林恢复过程中硝化、反硝化功能基因的丰度并探索硝化、反硝化功能基因的丰度与土壤理化性质的关系,明确次生林恢复过程中硝化、反硝化的影响因素。借此评估次生林的恢复重建,参与氮循环的特定功能微生物是否也能够随之恢复到先前水平。为该地区森林生态系统功能恢复和重建,温室气体的排放等研究提供方法和思路。
1 研究区和研究方法
1.1 研究区概况
试验样地依托于福建省上杭縣白砂国有林场(25°04′~25°15′N,116°30′~116°38′E),属于武夷山脉南段的低山丘陵地带,中亚热带气候,海拔为400~800 m,坡度10°~40°,年平均温度为20.1℃,年平均降雨量1600 mm,无霜期270 d左右。该地区土壤主要由花岗岩发育而成的红壤组成,土层深厚,立地条件中等。根据林业历史资料选取了土壤结构一致,并且都是皆伐地上发育的林地,皆伐之前,林地的树种组成一致,都是成熟的天然常绿阔叶林[8-9]。
1.2 样地设置与样品采集
2019年5月,采用空间代替时间的方法,选取5年、8年、21年、27年、40年、大于100年生的6个林龄段天然林作为研究对象,并在每个林龄段设置3~4个试验小区,大小为20 m×30 m,相同林龄段样地之间的间隔大于5 km,共计21个试验小区。
土壤样品于2019年5月,在不同恢复阶段的次生天然林样地内,使用3.5 cm土钻在各小区随机选取8~10个点取0~10 cm土层的土壤样品,自封袋密闭保存后及时带回实验室行土壤样品前处理。将土壤样品中的碎石以及植物残体用镊子挑出后,过2 mm筛网。试验所需土壤样品分为3部分:(1)储存于4℃冰箱内的用于测定土壤铵态氮、硝态氮、可溶性有机碳氮等理化性质;(2)部分放置室内等待自然风干后过0.149 mm的筛网,测定土壤pH、土壤总碳总氮含量等;(3)用于土壤微生物DNA的提取和土壤微生物功能基因丰度的测定保存在无菌管内,储存于-80℃冰箱中备用。
1.3 土壤理化性质测定
土壤pH采用电位法(土水比为1∶2.5),用pH计测定;土壤含水率测定采用烘干法,称取10 g鲜土在105℃的烘箱中连续烘干24 h直至恒重后称重计算;土壤全碳、全氮称取约1 g研磨后风干土样,使用碳氮元素分析仪(Elemental EL MAX CNS analyzer,德国)测定并计算土壤碳氮比;NH4+-N和 NO3--N含量为称取相当于 10 g干土的新鲜土样,加入 100 mL 2 mol·L-1的 KCl溶液浸提后,在连续流动分析仪上测定。可溶性有机碳(DOC)采用冷水浸提和总有机碳分析仪测定;连续流动分析仪测定土壤无机氮和总氮含量,通过公式计算土壤有机氮(DON)含量。土壤微生物生物碳氮采用氯仿熏蒸法,连续流动分析仪测定。
1.4 土壤DNA提取与荧光定量PCR
称取0.25 g土壤样品,使用MOBIO的PowerSoil DNA Isolation Kit试剂盒提取后,分光光度计检测DNA纯度和浓度后,在-20℃冰箱储存备用。
荧光定量PCR(qPCR)在伯乐CFX96定量PCR仪上进行。定量PCR所用的反应体系均为20 μL,包含:10 μL SYBR Premix Taq DNA聚合酶(TaKaRa,日本)、各0.4 μL的前后引物、8.2 μL ddH2O和1 μL DNA模板。通过连续稀释质粒构建每个功能基因的标准曲线,根据标准曲线计算样品的基因拷贝数。结果扩增效率在88%~100%,相关系数r2>0.99。
1.5 数据分析与处理方法
数据分析与处理均基于SPSS 21.0软件分析。通过单因素方差分析和多重比较法(LSD)分析不同恢复阶段之间土壤硝化、反硝化功能基因丰度的差异;使用皮尔逊相关系数检验功能基因丰度与土壤理化性质之间的相关性;土壤理化性质对硝化、反硝化功能基因丰度的影响采用冗余分析(RDA)实现;逐步回归分析利用vegan包中的lm和step函数在R软件( Version 3.4.1)上进行。
2 结果与分析
2.1 次生天然林不同恢复阶段土壤理化性质分析
由表1可知,次生天然林在恢复过程中,土壤理化性质pH、碳氮养分含量等也随着时间增长发生改变。土壤 pH随着森林恢复年限的增长呈下降趋势,大于100年生的原始林pH为 4.2,显著低于其他恢复阶段;土壤总碳、总氮、有机碳氮在森林恢复期间出现下降趋势,但是大于100年原始林土壤养分含量又出现回升,达到最大值。这说明天然次生林恢复早期,森林植被处于生长阶段,土壤养分被植物吸收利用较多,随着恢复至后期,植物吸收养分减少,而凋落物数量的增多,养分逐渐归还至土壤中,土壤有机碳氮等养分含量回升。
次生天然林恢复过程中土壤氮素形态主要以铵态氮为主,含量上为5年生天然林最低为7.49 mg·kg-1,显著低于其他阶段的次生天然林;硝态氮在恢复不同阶段无显著差异。随着次生林的恢复发展,土壤中铵态氮硝态氮含量均呈现上升趋势,并且逐渐达到与原始林相当的水平。此外森林恢复期过程中,8年生的土壤微生物生物量碳MBC显著高于其他林龄段,40年显著低于其他林龄段;微生物生物量氮 MBN在不同恢复阶段差异不显著。森林恢复期间土壤微生物生物量碳 MBC和微生物生物量氮 MBN均呈现下降的趋势,但与大于100年的原始林相比差异不显著。土壤含水率在不同恢复阶段差异不显著。
2.2 次生天然林不同恢复阶段硝化、反硝化功能基因丰度分析
由图1可以知,土壤AOA基因丰度的变化范围为7.82×105~2.33×107 copies·g-1(DW),其中大于100年的原始林基因丰度显著高于其他恢复阶段,总体上看,5~40年恢复期间AOA基因丰度变化趋势平稳;土壤AOB基因丰度不同恢复阶段之间差异不显著,其变化范围在 5.41×104~1.57×105 copies·g-1(DW);各恢复阶段的土壤样品中AOA基因丰度都要显著高于AOB,这表明数量上AOA基因在硝化功能微生物中占据多数地位。
由图2可知,NarG基因丰度8年的显著高于5年和40年的次生林,随着恢复时间的增长,土壤NarG基因丰度呈现上升趋势;编码亚硝酸盐还原酶的NirK和NirS基因具有相同的功能但属于不同的基因类型,NirK和NirS基因丰度的变化范围分别为1.71×107~3.74×107 copies·g-1(DW)、5.98×107~1.20×108 copies·g-1(DW),土壤NirK和NirS基因丰度在恢复过程中变化趋势较为一致,基因丰度上NirK始终高于NirS,不同恢复阶段均差异不显著;NosZ基因能够决定反硝化过程的产物,丰度的变化范围为1.09×108~3.24×108 copies·g-1(DW),结果表明次生天然林恢复早期的NosZ基因丰度显著低于恢复后期,基因丰度随时间推移而增高。
结合4种反硝化微生物功能基因来看,数量上呈现出8年和27年的反硝化功能基因丰度高于其他恢复阶段,整体上随恢复时间推移表现出相一致的趋势,反硝化基因丰度均随着恢复时间增长波动上升,逐渐恢复到与原始林相当的水平。此外研究发现,NosZ基因丰度大于 NirK和NirS的丰度和,其比值为1.52~2.04,这说明次生天然林生态系统中,反硝化过程的最终产物N2的产量大于N2O,这有助于缓解土壤在反硝化过程中产生的N2O向大气的排放。
2.3 土壤硝化、反硝化功能基因丰度与土壤理化性质的关系分析
由图3可知,土壤理化性质总共解释了土壤硝化、反硝化功能基因丰度74.9%的总变化,第一轴和第二轴分别解释土壤样品中61.5%的硝化和13.4%的反硝化功能基因丰度的变化。起主要作用的土壤理化性质的因子包括土壤 pH、土壤总氮含量以及土壤C/N。
由表2可知,硝化过程中丰度上占多数的AOA基因丰度与土壤TN、NO3--N、含量极显著正相关,与土壤NH4--N、DOC显著正相关,土壤C/N显著负相关;AOB基因丰度则与土壤理化性质并无显著相关性。参与反硝化过程的功能基因中,NarG基因丰度与土壤TN显著正相关;NirK基因丰度与土壤TN和DOC极显著正相关,与土壤TC和DOC显著正相关;氧化亚氮还原酶基因NosZ丰度与土壤C/N极显著负相关;同样是还原亚硝酸盐的功能基因NirS丰度则与土壤理化性质并无显著相关性。
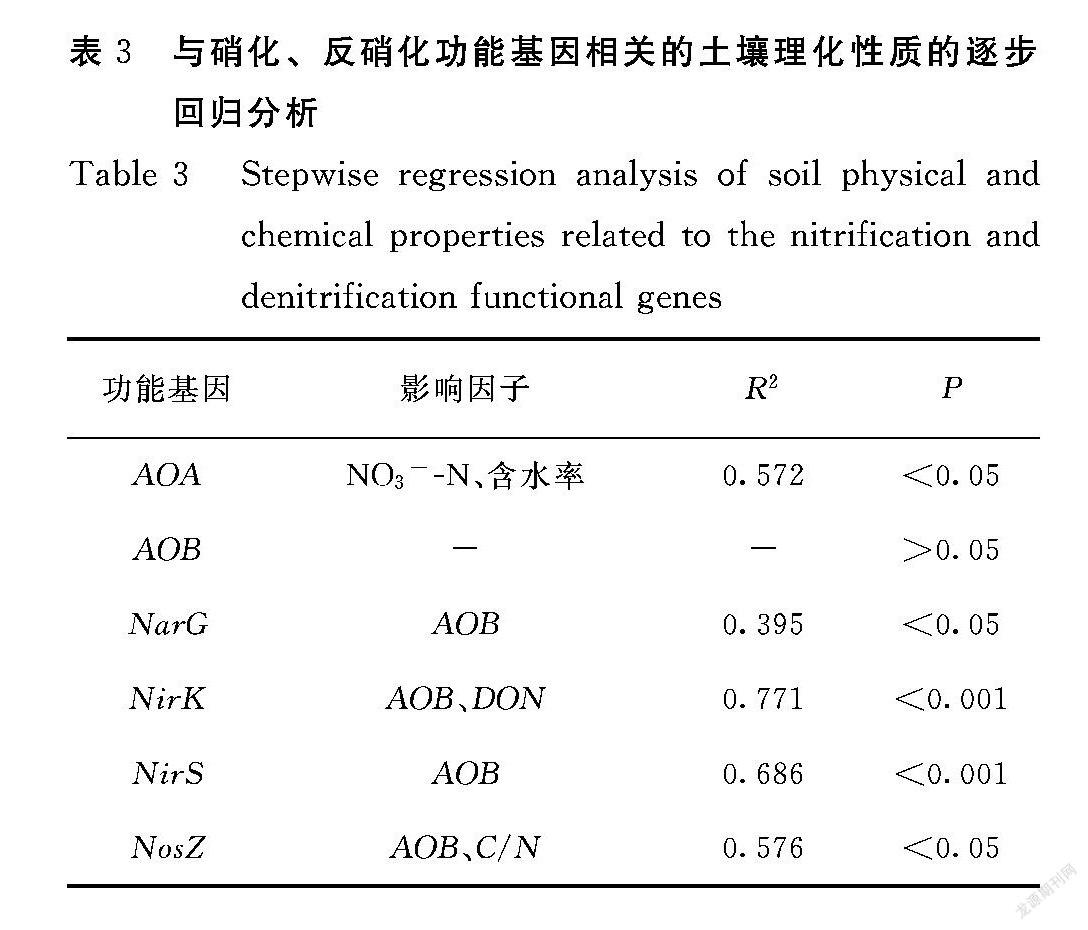
由表3可知,硝态氮和含水率是影响AOA基因丰度最主要的环境因子,可以解释57.2%的变量;反硝化基因NarG和NirS丰度主要受AOB的影响,可以解释 39.5%和68.6%的变量,反硝化基因NosZ丰度主要受AOB和有机氮的影响,可以解释77.1%的变量。反硝化基因NirK丰度主要受AOB和C/N的影响,可以解释57.6%的变量。
3 讨论
3.1 影响次生天然林土壤硝化微生物功能基因丰度的因素
土壤 pH是影响硝化功能微生物分布以及丰度的最主要因素[10]。已有研究表明,AOA基因丰度会随着土壤pH值下降而显著上升,而AOB基因丰度则呈现相反即下降的趋势[11-12]。He等
[13]在酸性土壤中的研究發现,AOA基因丰度与pH之间存在明显的负相关。AOB基因虽然有研究表明其参与硝化过程,但AOA基因在酸性土壤中对硝化过程的作用通常比AOB基因高得多,尤其是在天然林生态系统,AOB基因被认为是硝化过程的潜在贡献者[14]。本研究中发现AOA基因拷贝数与土壤pH并无显著相关性,可能是因为该地区的土壤pH变化范围为4.20~4.48,变化梯度较小导致作用不明显,但与对照原始林的结果分析,原始林土壤pH要显著低于其他恢复阶段的次生天然林,其土壤AOA基因丰度也是要明显高于其他阶段。此外不同恢复阶段样地中的土壤AOA基因丰度均要显著高于AOB基因丰度,其丰度比为6.6~160,这与此前大多数的研究结果一致[14]。其原因是AOA较AOB更能适应酸性土壤,AOA基因由于其独特的生物化学以及遗传学特性,使之更好地适应低pH环境[15]。在本研究中还通过相关分析发现土壤NO3--N含量与土壤AOA丰度极显著正相关,而NO3--N作为硝化过程的主要产物,表明森林恢复过程中功能基因AOA参与和主导土壤的硝化过程。
土壤中NH3的含量与可利用性同样是影响硝化微生物数量的重要影响因素,冗余分析结果表明土壤矿质氮含量与AOA基因丰度显著正相关,相关分析也进一步发现土壤AOA基因丰度与土壤NH4--N的含量呈显著正相关关系,这是因为NH4--N作为硝化过程的底物,其含量可能会影响到硝化微生物功能基因的丰度[16]。在本研究中也有体现,例如次生林恢复早期土壤由于铵态氮含量低,植物对土壤养分的竞争导致微生物功能基因丰度AOA低,但随着土壤养分的累积和植物获取能力的下降,铵态氮含量回升,其AOA基因丰度也达到最大值。
3.2 次生天然林土壤反硝化功能基因丰度的影响因素
本研究中,通过定量参与反硝化过程NarG、NirK、NirS和NosZ 4种功能基因分析不同恢复阶段次生林土壤反硝化功能基因的特征及影响因素。冗余分析结果发现土壤TN含量与反硝化功能基因丰度显著正相关,土壤C/N则与反硝化微生物基因丰度呈负相关关系,表明土壤中碳氮养分含量为反硝化微生物活动提供了底物和能源,这与Chen等的研究结果一致[17]。相关分析表明土壤TN含量与NarG基因丰度显著正相关;土壤养分TC、TN、DOC、DON含量与NirK基因丰度显著正相关,但NirS基因丰度与土壤理化性质无显著关系(P>0.05),这表明NirK基因丰度比NirS对环境因子的变化更为敏感;NosZ基因与土壤碳氮比显著负相关;此外有研究表明反应底物的NO3--N含量与反硝化微生物功能基因的丰度密切相关[4][18-19],在本研究中底物含量与功能基因丰度均没有显著的相关性,可能是因为土壤NO3--N的含量较低且在不同恢复间并无显著差异有关。本研究还发现,除了土壤理化性质会影响反硝化功能微生物基因丰度,参与硝化过程的功能基因AOB是NarG、NirK、NirS和 NosZ丰度变化的最主要影响因素,这强调了酸性土壤中AOB基因的作用和影响,作为反硝化过程的前一步骤的参与者,其丰度变化对后续反硝化过程起到深刻影响,后续应该加强两个过程之间关系的研究,有助于加深对氮循环潜在机制的理解。
(NirK+ NirS)/NosZ的比值常用来评价土壤N2O的排放潜势。本研究发现在次生天然林恢复过程中(NirK+ NirS)/NosZ比值小于1,说明在次生天然林恢复过程中,NosZ功能基因可以将反硝化过程产生的N2O转化为N2,NosZ基因丰度上的优势提高了对N2O的转化能力,减少土壤微生物活动产生的N2O向大气的排放。从微生物角度表明自然恢复次生林有着较为封闭的氮循环过程,次生天然林中的氮素不易流失,有利于土壤养分的恢复和积累,这与先前的研究结论相一致。
4 结论
通过对硝化、反硝化功能基因丰度的定量与分析,研究表明,随着次生林的自然恢复,土壤理化性质特别是碳氮养分会恢复到与原始林相当的水平,而受pH、碳氮养分影响的硝化、反硝化功能基因丰度也会随着恢复时间的推移而达到先前森林未破坏的水平,其中森林恢复期间硝化基因AOA始终主导着次生林天然土壤的硝化过程,但是反硝化功能基因NarG、NirK、NirS和NosZ会受到AOB基因丰度的影响,这强调了AOB基因在氮循环整体中的地位和作用,也明确了硝化过程在氮循环中心环节的地位。反硝化功能基因(NirK+ NirS)/NosZ丰度比小于1,表明自然恢复的次生林能够更好地将氮素保持在土壤中,减少温室气体N2O的排放,这为后续该地区森林恢复模式的选择提供了科学依据。
参考文献:
[1]THAMDRUPBO.New Pathways and Processes in the Global Nitrogen Cycle[J].Annual Review of Ecology Evolution & Systematics,2012,43(1):407-428.
[2]贺纪正,张丽梅.土壤氮素转化的关键微生物过程及机制[J].微生物学通报,2013,40(1):98-108.
[3]CANFIELD D E,GLAZER A N,FALKOWSKI P G, et al.The Evolution and Future of Earth's Nitrogen Cycle[J].Science,2010,330(8):192-196.
[4]LEVY-BOOTH D J, PRESCOTT C E,GRAYSTON S J.Microbial functional genes involved in nitrogen fixation, nitrification and denitrification in forest ecosystems[J].Soil Biol Biochem,2014,75(1):11-25.
[5]LAMMEL D R, FEIGL B J, CERRI C C, et al.Specific microbial gene abundances and soilparameters contribute to C, N, and greenhouse gas process rates after land use changein southern Amazonian soils[J].Frontiers in Microbiology,2015,6(1057):1057.
[6]王蕓,欧阳志云,郑华,等.不同森林恢复方式对我国南方红壤区土壤质量的影[J].应用生态学报,2013,24(5):1335-1340.
[7]YUAN B C, YUE D X.Soil microbial and enzymatic activities across a chronosequenceof Chinese pine plantation development on the Loess Plateau of China[J].Pedosphere,2012, 22(1):1-12.
[8]王振宇,傅彦榕,邹秉章,等.亚热带森林不同恢复模式土壤与林下植被特征的动态研究[J].福建农业科技,2021,52(8):7-16.
[9]杨军钱.不同林龄天然次生林土壤氮素转化与相关功能微生物研究[D].福州:福建师范大学,2020.
[10]WU Y, XIANG Y, WANG J,et al.Heterogeneity of archaeal and bacterial ammonia-oxidizing communities in Lake Taihu, China[J].Environmental Microbiology Reports,2010,2(4):569-576.
[11]SHEN J P, ZHANG L M, DI H J, et al. A review of ammonia-oxidizing bacteria andarchaea in Chinese soils[J].Frontiers in Microbiology,2012,3(1):296-296.
[12]HE J Z, SHEN J P, ZHANG L M, et al.Quantitative analyses of the abundance andcomposition of ammonia-oxidizing bacteria and ammonia-oxidizing archaea of a Chinese upland red soil under long-term fertilization practices[J].Environmental Microbiology,2007,9(9):2364-2374.
[13]LI Y Y, CHAPMAN S J, NICOL G W, et al.Nitrification and nitrifiers in acidic soils[J].Soil Biology and Biochemistry,2018,116(1):290-301.
[14]HE J Z, HU H W, ZHANG L M, et al.Current insights into the autotrophicthaumarchaealammonia oxidation in acidic soils[J].Soil Biology and Biochemistry, 2012,55(6):146-154.
[15]刘卜榕.亚热带四种主要森林土壤氨氧化微生物群落结构及多样性研究[D].杭州:浙江农林大学,2012.
[16]CHEN J, LI J, SHEN W, et al.The structure and species co-occurrence networks of soildenitrifying bacterial communities differ between a coniferous and abroadleavedforests[J].Microorganisms,2019,7(9):361.
[17]XIE Z, ROUX X L, WANG C, et al. Identifying response groups of soil nitrifiers anddenitrifiers to grazing and associated soil environmental drivers in Tibetan alpine meadows[J].Soil Biology and Biochemistry,2014,77(3):89-99.
[18]PHILIPPOT L, CUHEL J, SABY N, et al. Mapping field-scale spatial patterns of size andactivity of the denitrifier community[J].Environmental Microbiology,2009,11(6):1518-1526.
[19]DESALEGN T, CRUZ F, KINDU M, et al. Land-use change and socioeconomic conditions of local community in the central highlands of Ethiopia[J].International Journal of Sustainable Development & World Ecology, 2014,21(5):406-413.
[20]王磊.亞热带不同树种对土壤氮保留及相关功能微生物的影响[D].福州:福建师范大学,2019.
[21]郑荧枫,李雪,万晓华,等.次生林不同演替时间土壤微生物群落结构的变化[J].亚热带资源与环境学报,2021,16(1):23-28,34.
[22]朱永官,沈仁芳,贺纪正,等.中国土壤微生物组:进展与展望[J].中国科学院院刊, 2017,32(6):554-565.
[23]罗蓉.黄土高原油松人工林参与土壤氮循环功能微生物群落结构研究[D].咸阳:西北农林科技大学,2018.
[24]张青山,欧阳运东,肖孔操,等.喀斯特峰丛洼地植被恢复对土壤硝化与反硝化潜势的影响[J].农业现代化研究,2018,39(3):520-526.
[25]VANG J W, HUYGENS D, BOECKXP, et al.The soil N cycle: new insights and key challenges[J].Gestión Y Análisis De Políticas Publicas,2014,1(1):235-256.
(责任编辑:柯文辉)
