
H2S对CuO NPs胁迫下番茄幼苗Cu分布及细胞壁组分的影响
2022-05-27 03:40贾红磊马佩云黄蓝青
陕西科技大学学报 2022年3期
贾红磊, 陈 翠, 马佩云, 黄蓝青
(陕西科技大学 环境科学与工程学院, 陕西 西安 710021)
0 引言
纳米氧化铜(CuO NPs)是一种重要的纳米金属材料,被作为涂覆材料、催化剂、润滑剂、杀菌剂等广泛应用于各领域[1].随着 CuO NPs 的广泛应用,越来越多的CuO NPs 逐步渗透到人类的生存环境和生态系统中,并发生复杂的迁移转化、环境行为和生态效应[2-4].植物是生态系统的重要组成部分,进入环境的纳米氧化铜可被其吸收、累积,并造成严重的毒效应.其致毒机制主要通过以下三方面:(1)释放Cu2+产生离子毒害作用;(2)诱导生成活性氧(ROS),产生氧化胁迫造成细胞膜、细胞器和DNA的损伤;(3)纳米颗粒本身通过物理吸附作用附着于受害体表面或堵塞细胞膜上的各种离子通道,导致细胞损伤.且有研究表明CuO NPs表现出的纳米颗粒生物毒性远大于其释放的Cu2+的毒性,所以CuO NPs对生物毒害作用受到广泛关注[5].
植物细胞壁具有一定弹性和硬度,存在于细胞质外,可为细胞提供结构支持和保护作用.植物细胞壁分为中层、初生壁和次生壁.中层主要由果胶和蛋白质组成;初生壁具有纤维素微纤丝的刚性骨架,其中镶嵌有果胶、半纤维素和糖基化蛋白;次生壁质地坚硬,有增强细胞壁机械强度的作用,由纤维素、半纤维素和木质素组成[6].细胞壁组分的果胶和半纤维素具有大量的羧基、醛基等基团,这些基团可以吸附并区隔化固定重金属离子,减少重金属离子向原生质体迁移,缓解对细胞的毒害作用[7].因此细胞壁在维持植物生长和抵抗非生物胁迫中有着重要的作用[8].
硫化氢(H2S)是存在于细菌、无脊椎动物、植物和哺乳动物中的一种气体信号分子[9],对植物的研究中外源H2S的供体通常使用硫氢化钠(NaHS)[10].H2S不仅在植物生长发育中起着重要作用,而且还影响着植物在非生物胁迫下的反应[11],在植物体内广泛参与各种生理反应,比如调节植物气孔闭合[12],减缓植物硼毒对根伸长的抑制[13],降低Cd在植物体内的积累和氧化损伤[14],增加Cu胁迫下植物种子的萌发率[15].目前尚不清楚H2S是否会缓解CuO NPs对番茄幼苗的胁迫.通过研究CuO NPs胁迫下外源H2S对番茄幼苗体内Cu分布,以及细胞壁组分的影响.为外源H2S调控植物抵御纳米重金属胁迫提供一定理论基础.
1 实验部分
1.1 试验材料与设计
试供植物为番茄(SolanumlycopersicumL.),品种为石红三号,发芽率为90%,购自新疆石河子蔬菜研究所番茄研究开发中心.
挑选大小均一,饱满的种子用3%的 NaClO溶液浸泡消毒15 min,然后用去离子水冲洗多次,均匀放置在装有蛭石的穴盘中,避光放置,待种子发芽后用棉花包裹幼苗茎部转移至含有1/4 Hoagland的水培箱培养两周,之后1/2浓度的Hoagland 营养液水培2周.培养条件为:光照16 h黑暗8 h,相对湿度40%~60%,温度为26±2 ℃(昼),20±2 ℃(夜),培养至有四片真叶长出时选取长势良好且一致的幼苗,移至不同的黑色处理罐中培养,根据实验需求收样测定相关指标.
试验共设置7组处理,分别为对照组(Control)、200μM NaHS(NaHS)、20μM CuSO4·5H2O (Cu2+)、20 mg/L CuO NPs (CuO NPs1)、20 mg/L CuO NPs+NaHS (CuO NPs1+NaHS)、 100 mg/L CuO NPs (CuO NPs2)、100 mg/L CuO NPs+NaHS (CuO NPs2+NaHS).
1.2 实验方法
1.2.1 番茄幼苗体内Cu亚细胞分布
Cu的亚细胞分布采用差速离心法,参照Hao等[16]的方法进行,将所得的三部分样品消解定容后采用原子吸收分光光度计(ZEEnit700p,耶拿,德国)测定Cu含量.
1.2.2 番茄幼苗细胞壁各组分中Cu分布
植物细胞壁主要成分有:果胶、纤维素、半纤维素[17]番茄幼苗细胞壁提取方法参照Ren等[18]的方法.准确称取10 mg提取的细胞壁干样,分步进行果胶、半纤维素、纤维素组分的提取.果胶根据提取剂不同可分为水溶性果胶(WSP)、螯合型果胶(CSP)、碱溶性果胶(NSP).半纤维素(HC)又可分为半纤维素1(HC1)和半纤维素2(HC2),具体参照Wu等[19]的方法.将各部分提取液以及最后剩余残渣经电加热消解仪消解后于火焰原子吸收分光光度计下测定各部分Cu含量.
1.2.3 番茄幼苗细胞壁多糖组分含量测定
果胶染色根据Molnar等[20]的方法并稍作修改,用0.002 5%(w/v)的钌红染液(溶于10 mM Tris-HCl,pH=7.4)孵育番茄根系15 min,之后用去离子水将根系表面附着的染料冲洗干净,压片置于光学显微镜下观察染色情况.根据Ren等[18]所述的方法测定果胶以及半纤维素含量.纤维素含量测定采用蒽酮比色法,具体操作参照Viles等[21]的方法.
1.2.4 番茄幼苗根系木质素和胼胝质定性分析
木质素以及胼胝质染色方法均参照 Molnar等[20]的方法.采用间苯三酚-HCl溶液观察根中木质素的含量.用苯胺蓝溶液染色检测胼胝质.
2 结果与讨论
2.1 H2S对CuO NPs胁迫下番茄幼苗Cu亚细胞分布的影响
有研究表明重金属的亚细胞分布与植物中的重金属解毒有关[22].为了确定H2S对番茄幼苗中Cu毒性的调控作用,测定了Cu在实验番茄幼苗根和叶亚细胞的分布情况,如图1和2所示.实验结果表明,总量85.25%的Cu主要分布在细胞壁和细胞质中.由图1可知,在根中不同浓度CuO NPs处理比Cu2+处理显著增加Cu在亚细胞中的分布,表明CuO NPs比Cu2+具有更强的生物毒性.与单独CuO NPs处理相比,添加NaHS降低了根中三种组分Cu的含量,细胞器中CuO NPs1和CuO NPs2处理下分别降低20.73%和30.66%.由图2可知,叶中所有处理组细胞质Cu含量>细胞壁>细胞器, CuO NPs1和CuO NPs2处理组细胞器中Cu含量分别比对照组升高了38.76%和98.45%,添加NaHS后抑制了其上升的趋势.CuO NPs 胁迫下NaHS对细胞质中Cu含量的影响不显著,但显著降低了细胞壁和细胞器中Cu含量.H2S能够抑制植物体对重金属的吸收,因此显著降低细胞器中重金属的含量[23,24].本研究发现,NaHS能够通过调节生物体内的CuO NPs的亚细胞分布,降低细胞壁和细胞器中Cu含量,减轻CuO NPs对番茄幼苗的毒性效应.
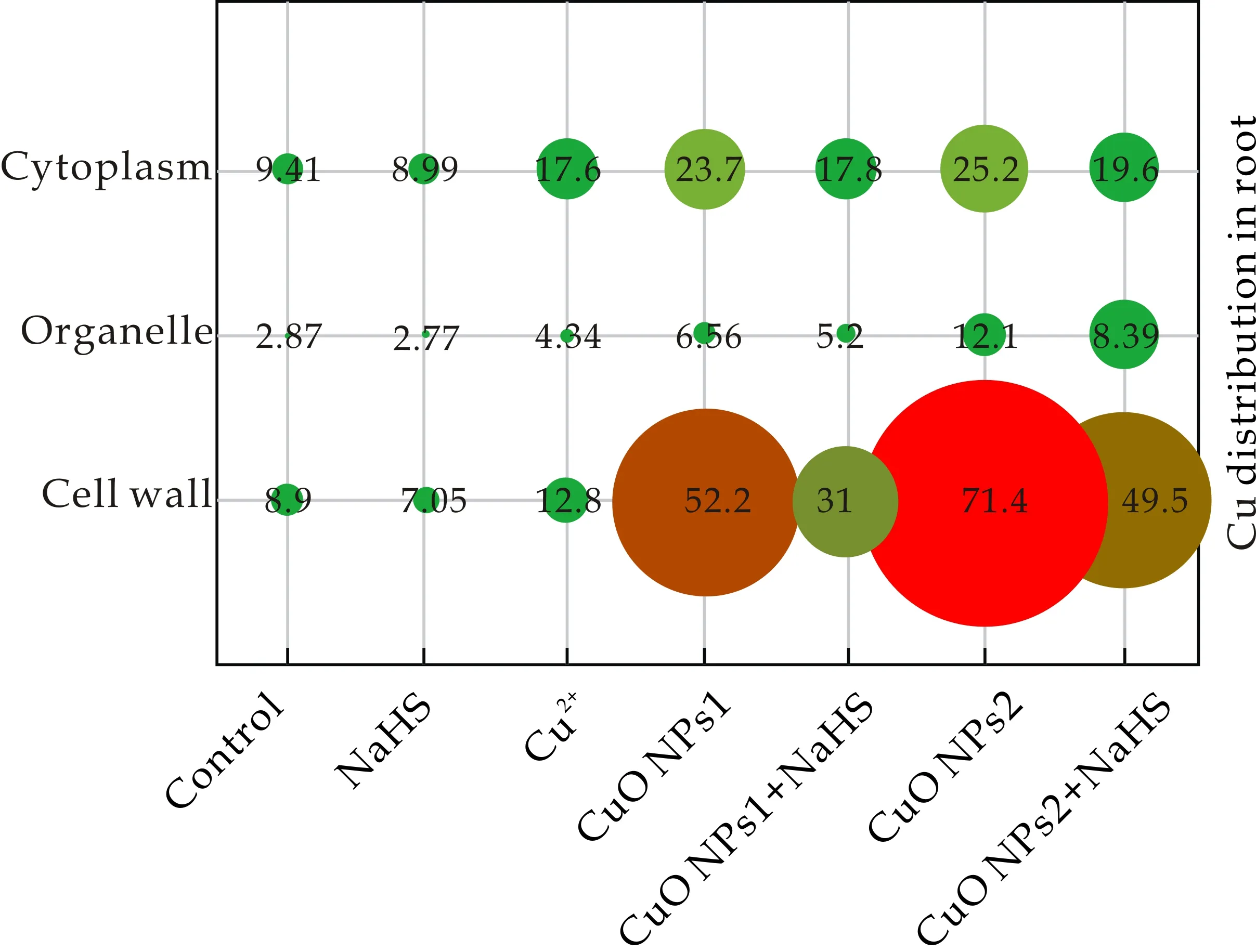
图1 不同处理下番茄幼苗根亚细胞中Cu的分布
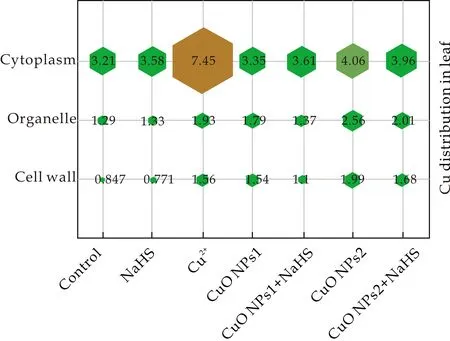
图2 不同处理下番茄幼苗叶亚细胞中Cu的分布
2.2 H2S对Cu在番茄幼苗细胞壁多糖组分中分布的影响
细胞壁中各种化学组分(果胶、纤维素、半纤维素、蛋白质等)中的负电基团通过沉淀、吸附、络合等作用将金属离子固定在细胞壁中,从而减少金属离子通过跨膜运输进入原生质体,在一定程度上降低了金属胁迫对植物正常生理活动的干扰[25,26].
2.2.1 果胶Cu分布
果胶作为植物细胞壁主要成分之一,它在植物的生长发育、维持植物的形态结构、调节细胞的渗透性及pH、信号转导、结合重金属离子等方面发挥着重要的作用[26].图3所示为经不同处理后番茄幼苗果胶中Cu含量.由图3(a)、(c)、(e)可知,不同处理下番茄幼苗根WSP中Cu含量均低于CSP和NSP中Cu含量,表明CSP和NSP结合Cu能力较强,而WSP结合Cu能力较弱.单独添加CuO NPs1和CuO NPs2后各形态果胶中Cu含量均显著高于对照组和Cu2+处理组,再添加NaHS后CSP中分别下降7.77%和27.51%;NSP中分别下降33.32%和24.28%;而WSP则呈上升趋势.
由图3(b)、(d)、(f)可知,叶与根不同的是,叶中三种形态果胶中Cu含量均较低,且在添加NaHS后与单独CuO NPs1处理相比,CSP和NSP中Cu含量无显著差异.与根中变化趋势一致的是,CuO NPs2处理组加入NaHS后WSP显著升高,而CSP和NSP显著降低.由此表明,NaHS可以通过显著降低CSP、NSP中Cu的含量,以降低CuO NPs胁迫下番茄幼苗细胞壁中Cu的积累.有研究表明提高果胶的含量可以使果胶结合更多的重金属,导致细胞壁中重金属含量上升[27].CuO NPs胁迫下,加入NaHS后果胶含量的改变可能是影响果胶结合Cu含量变化的重要因素.

(a)根WSP中Cu含量
2.2.2 半纤维素Cu分布
除果胶外,组成细胞壁的另一主要多糖物质为半纤维素,因含有大量的-OH,在细胞壁的重金属富集中也发挥着一定的作用[28].在本研究中,对番茄幼苗经不同处理后半纤维素中Cu含量的测定结果如图4所示.由图4(a)、(c)可知,根HC1和HC2中Cu含量在CuO NPs1+NaHS和CuO NPs2+NaHS处理下与对照、Cu2+和NaHS处理相比显著增加,与CuO NPs1、CuO NPs2处理相比HC1中分别显著降低了27.51%和19.66%,HC2中分别显著降低了2.45%和6.16%.所以NaHS的添加可以降低CuO NPs胁迫下HC1和HC2中Cu含量,且对HC1中Cu含量调节作用强于HC2.
由图4(b)、(d)可知,CuO NPs1胁迫下添加NaHS对HC1、HC2中Cu含量影响不显著; 同样NaHS对CuO NPs2胁迫下HC2中Cu含量影响也不显著,仅在HC1中显著下降了10.20%.所以NaHS对细胞壁HC1中Cu含量有更强的调节作用.Jie等[29]研究发现随着半纤维素含量的提高,结合Cd的基团数量增多,半纤维素中Cd含量上升.本研究中CuO NPs胁迫导致HC1中Cu含量显著上升,而H2S降低HC1对Cu的结合能力.因此推测H2S通过降低HC1的含量,以降低半纤维素中重金属的含量,最终降低植物对重金属的累积.
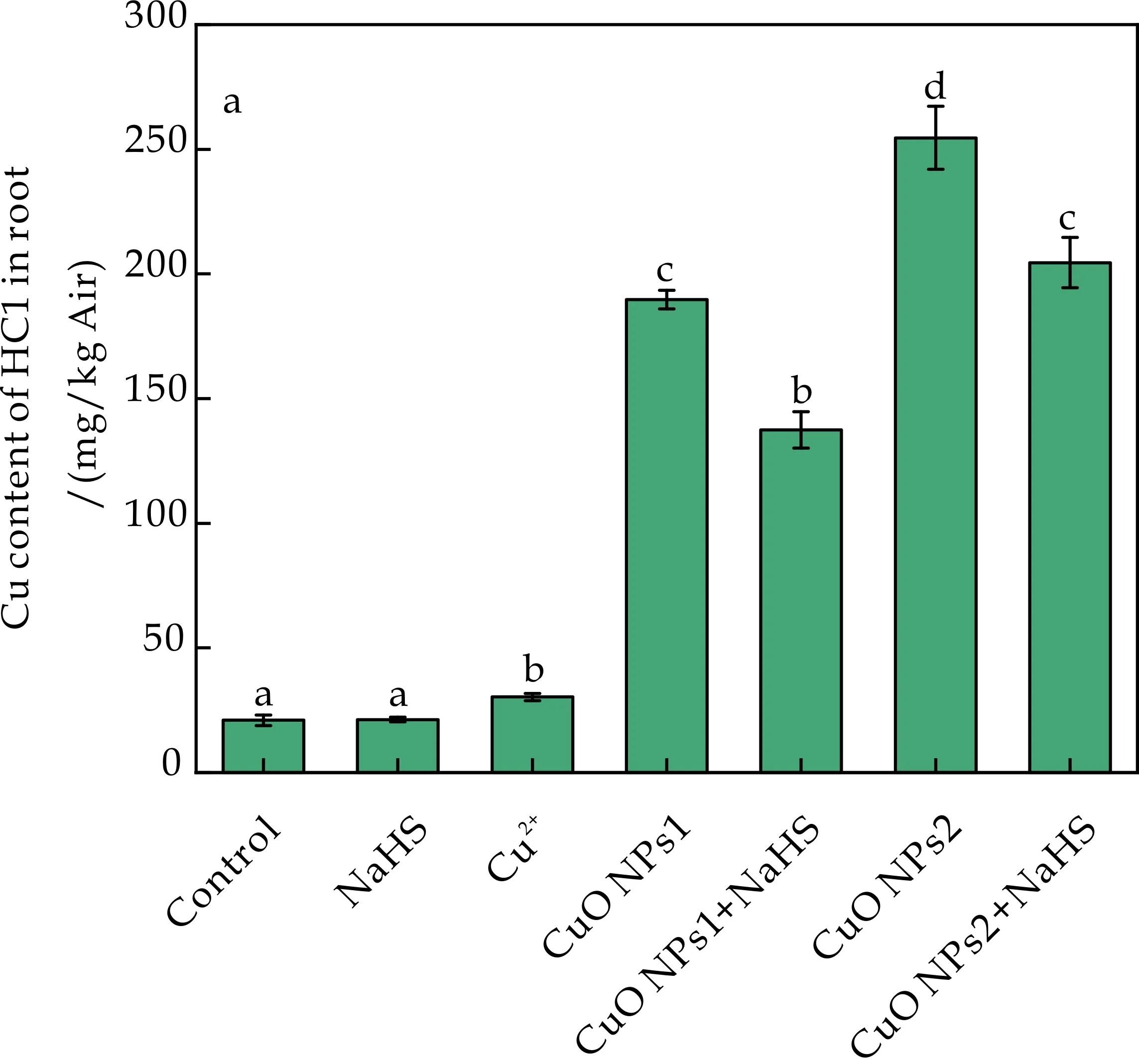
(a)根HC1中Cu含量
2.2.3 纤维素Cu分布
纤维素是细胞壁的骨架,为其提供了抗张强度,是植物体内负责机械强度的主要要素,其对于缓解重金属毒害效应有重要作用[30].如图5所示,测定了不同处理下番茄幼苗根和叶中纤维素Cu含量.结果表明,根和叶中纤维素Cu含量均较低为20~50 mg/kg,远远低于果胶、半纤维素中Cu含量.说明纤维素固定Cu能力较弱,但在根和叶中CuO NPs2处理下分别达到最高值46.28 mg/kg和36.58 mg/kg, NaHS参与调节后根中下降了5.08%(如图5(a)所示),叶中显著下降了17.30%(如图5(b)所示).表明NaHS参与调节番茄幼苗纤维素中Cu的累积.

(a)根纤维素中Cu含量
2.3 H2S对CuO NPs胁迫下番茄幼苗细胞壁多糖含量的影响
植物细胞壁是由多种多糖构成的,多糖上存在大量羧基、醛基等能够固定重金属的基团[31],当植物受到重金属胁迫时,这些离子基团可以将其固定在细胞壁,增强植物对重金属的耐受性.此外有研究发现外源因素可以影响细胞壁各组分含量,而细胞壁成分的改变会改变其对重金属的吸附量,及重金属在各组分中的分布,进而改变植物对重金属的耐受和富集能力[32,33].
2.3.1 果胶含量
果胶经钌红染液染色结果如图6所示,NaHS处理组果胶含量高于对照组,对照经CuO NPs处理后根中果胶明显减少,再加入NaHS后明显增加.同形态果胶含量测定结果如图7所示.由图7(a)可知, CuO NPs处理使得根中WSP和CSP含量相对于对照均显著下降,其中WSP下降幅度大于CSP,表明CuO NPs对WSP影响更大.NaHS再处理后WSP和CSP含量与单独CuO NPs处理相比均出现不同程度的上升,CuO NPs1+NaHS分别上升69.31%和14.32%,CuO NPs2+NaHS分别上升62.92%和36.20%.而NSP与之相反,CuO NPs处理使得其含量显著高于对照,且NaHS的加入使其明显降低,CuO NPs1胁迫下降低了30.15%, CuO NPs2胁迫下降低了19.39%.总果胶(TP)含量在CuO NPs胁迫下也呈下降趋势,与WSP和CSP含量变化一致.CuO NPs胁迫抑制细胞壁中WSP和CSP的合成,增加NSP含量[34],而外源H2S的加入减小CuO NPs作用下的果胶含量变化,缓解CuO NPs的胁迫.

图6 番茄幼苗根果胶钌红染色
由图7(b)可知,研究NaHS、Cu2+、CuO NPs和NaHS+CuO NPs处理对番茄幼苗叶中各形态果胶含量的影响.结果表明,两种剂量的CuO NPs(CuO NPs1和CuO NPs2)与对照相比,分别提升TP含量20.47%和38.18%,增加NSP含量3.27倍和5.05倍,而显著降低WSP和CSP含量.相比之下,外源NaHS的添加明显改善了CuO NPs对各形态果胶含量的负面影响,特别是在CuO NPs1胁迫下,恢复了WSP和CSP的含量;对NSP而言,与单独的CuO NPs处理相比,加入NaHS后CuO NPs1降低64%,CuO NPs2降低19.98%,一定程度上减轻CuO NPs的负作用.外源NaHS调节细胞壁中果胶的合成[13],增加胁迫下WSP、CSP含量,降低NSP含量,保持细胞壁完整性[35].有效的阻隔外部的CuO NPs,降低CSP、NSP中Cu的含量.图3为图7所示的不同处理下番茄幼苗果胶所结合的Cu含量.由图3可知,NSP具有较强结合Cu的能力,因此显著降低NSP的含量可以减少细胞壁中Cu的累积,减轻CuO NPs对番茄幼苗的胁迫.
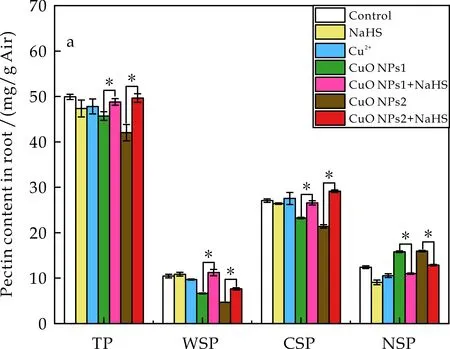
(a) 根果胶含量

(b) 叶果胶含量图7 不同处理下番茄幼苗细胞壁果胶各组分含量
2.3.2 半纤维素含量
图8表示实验番茄幼苗半纤维素含量.由图8(a)可知,根细胞壁中HC含量受CuO NPs影响,CuO NPs1胁迫下降低为77.74 mg/kg,加入NaHS后上升33.05%.添加两种剂量CuO NPs后HC1含量上升,再加入NaHS后降低18%( CuO NPs1处理)和17.41%(CuO NPs2处理).对于HC2,CuO NPs处理会降低其含量,相反加入NaHS后升高.可以看出,NaHS可以减轻CuO NPs所导致的HC含量变化,几乎恢复至胁迫前的状态.故施加NaHS可以缓解CuO NPs对细胞壁的损伤作用.
由图8(b)可知,在叶细胞壁中HC1与HC2含量变化趋势一致,添加CuO NPs后与对照相比均下降,再加入NaHS诱导其含量上升.与对照相比HC含量经CuO NPs1处理下降33.76%,CuO NPs2处理下降11.21%,得出CuO NPs1胁迫对HC含量影响较大,而添加NaHS后提升25.88%.综上可知,外源添加NaHS可以调节番茄幼苗中根、叶细胞壁中半纤维素含量,可能调整细胞壁结构的构建,提高细胞壁对重金属的屏蔽作用[36,37],从而降低CuO NPs对番茄植株的损害,改善细胞生长.

(a)根系半纤维素含量
2.3.3 纤维素含量
纤维素作为细胞壁的主要组成成分,植物受到Cd2+胁迫时,外源H2S增加细胞壁中纤维素的含量,保持细胞壁完整性[38].番茄幼苗经不同处理后纤维素含量测定结果如图9所示.其中 NaHS和CuO NPs1+NaHS处理后叶中纤维素含量与对照无明显差异,其余处理均显著高于对照.CuO NPs2处理后纤维素含量增加109.72%,继续添加NaHS后会有所下降.根细胞壁中CuO NPs1处理与对照相比显著降低,而CuO NPs1+NaHS比CuO NPs1处理增加64.16%.CuO NPs胁迫下,NaHS可能通过调节细胞壁中纤维素含量,影响细胞壁的组成结构[38],改变细胞壁中Cu含量.番茄幼苗根和叶应对CuO NPs胁迫的机制不同,加入NaHS可以调节细胞壁的组分,降低体内Cu含量,从而增强对CuO NPs的耐受性.
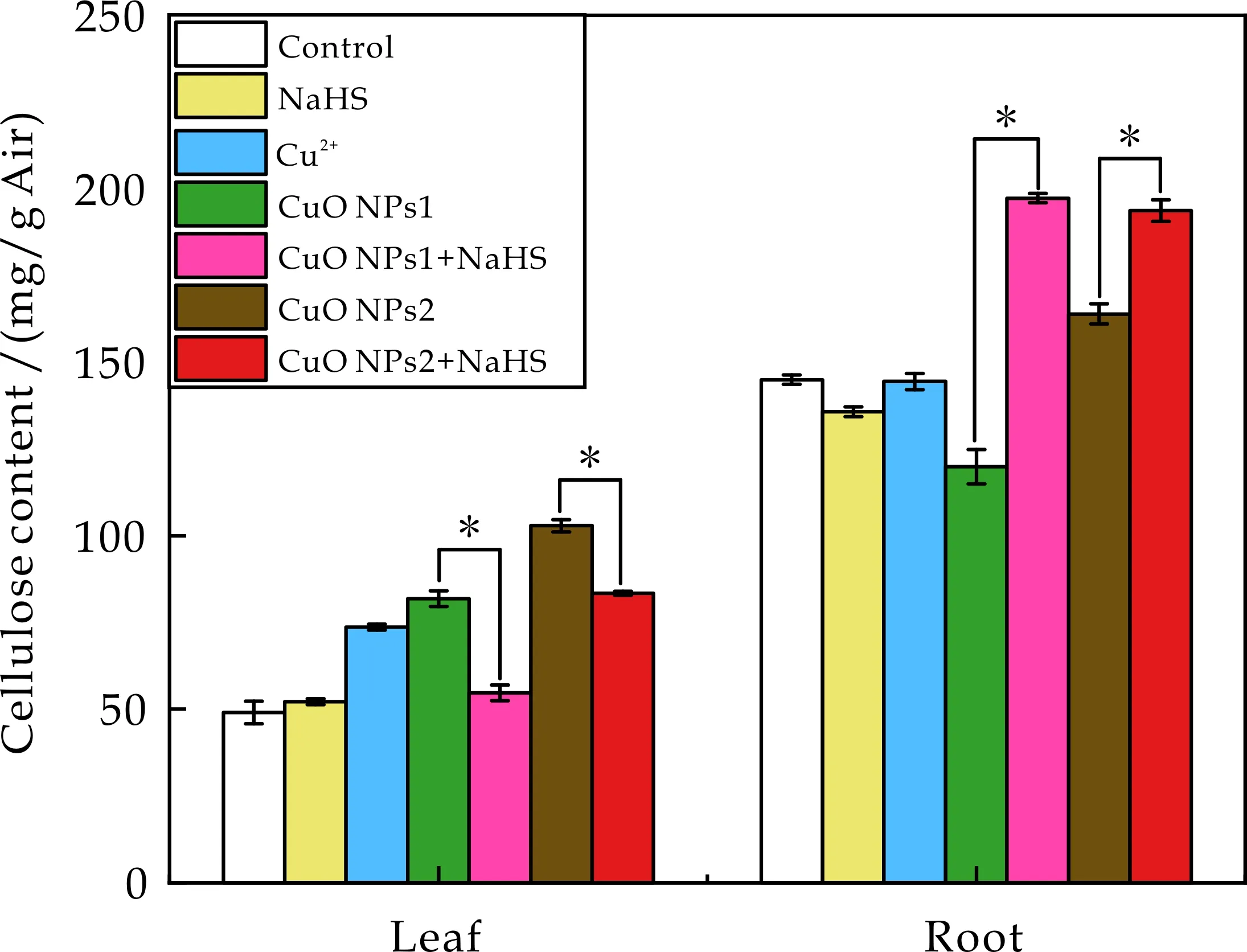
图9 不同处理下番茄幼苗细胞壁纤维素含量
2.4 H2S对CuO NPs胁迫下番茄幼苗根系木质素和胼胝质的影响
不同处理下番茄幼苗根系木质素染色结果如图10所示.经NaHS处理后木质素含量有所下降,而Cu2+处理没有对其造成明显影响.相反加入CuO NPs后,木质素大量累积,且与CuO NPs浓度呈正相关,再经NaHS处理后,木质素含量有下降趋势.

图10 番茄幼苗根木质素染色
此外研究发现,胼胝质作为应激反应的产物,在植物受伤、遭受病原菌感染或受到各种胁迫时快速而大量地合成[39].实验番茄幼苗根尖胼胝质染色结果如图11所示.对于CuO NPs2胁迫下胼胝质含量增加,添加NaHS后其含量显著下降,表明NaHS可以缓解CuO NPs的毒性作用.

图11 番茄幼苗根尖胼胝质染色
3 结论
通过上述研究,可以得出结论:CuO NPs比Cu2+具有更强的生物毒性,并逐渐渗入生态系统威胁着植物生长.结果表明,CuO NPs破坏番茄幼苗细胞壁结构引起体内Cu累积,表现为细胞壁多糖含量发生显著变化, 多糖和亚细胞中Cu含量显著升高.与此相反,施用NaHS可调节CuO NPs胁迫下番茄幼苗细胞壁组分,维持结构完整,降低细胞器中Cu累积和应激反应,最终达到增强番茄幼苗对CuO NPs的抵御.
猜你喜欢
造纸信息(2022年8期)2022-11-10
食品与发酵工业(2022年19期)2022-10-17
农产品加工(2022年9期)2022-06-17
大连工业大学学报(2022年1期)2022-03-14
广东工业大学学报(2022年1期)2022-02-11
蔬菜(2021年2期)2021-11-27
江苏农业科学(2021年8期)2021-05-26
科普童话·神秘大侦探(2021年12期)2021-01-19
三农资讯半月报(2020年18期)2020-10-14
中国造纸(2019年6期)2019-09-10
