
全新世西风模态下中亚干旱区孢粉类型多样性变化特征
——以阿尔泰山为例
2022-06-08 07:12张东良
干旱区研究 2022年3期
张东良
(1.中国科学院新疆生态与地理研究所,新疆乌鲁木齐 830011;2.中国科学院中亚生态与环境研究中心,新疆乌鲁木齐 830011;3.中国科学院大学,北京 100049)
全球平均气温正接近过去120 万年的上限[1],它们的影响在地球上每个生物群落都有记录[2]。随着未来几十年气候变化加剧,保护生物多样性以及生态系统为自然和人类提供的服务必须放在国际政策议程的重要位置[1-4]。有效的干预措施需要根据可靠的预测和基于证据的解决方案采取保护行动[2]。然而,因较短的时间观测导致物种到生态系统层面对气候变化响应的可靠预测仍很困难[4]。因此,从地质历史时期找寻生物多样性演化的证据备受关注[5],其中,地质记录中孢粉是最为丰富的古生态学记录,可以提供过去植物存在和分布的证据,是开展古植物多样性研究非常有效的材料之一[6]。
位于中亚干旱区的天山-阿尔泰山,是世界多样性热点地区之一。近年来,该区域增温幅度远超全球平均水平,使得极端气候事件出现的频次增加。该结果对干旱区脆弱生态系统造成了严重的威胁,导致树木生长下降和死亡[7-9]。最新预测表明,干旱和半干旱地区面临显著增温和干旱的气候特点,使得该区植被栖息地破碎,且导致未来全球变化对该区植被生态系统的演化产生更大的不确定性[10]。可见,我们迫切需要借助孢粉学手段来研究该区域孢粉类型多样性与气候之间的关系。已有学者对天山孢粉类型多样性和山地森林-草原交错带在全新世“西风模态”(即全新世早期降水少,全新世中晚期降水多)[11-12]变化下的动态规律进行了研究[13-14],但对阿尔泰山孢粉类型多样性和植被带的迁移过程仍不清楚。许多学者已在阿尔泰山不同海拔高度开展了一系列的孢粉组合研究[15-20]。这些研究可为我们开展过去气候变化是否引起阿尔泰山孢粉类型多样性增加/减少、是否影响植被带的变化提供数据支撑。
本研究共选取阿尔泰山不同海拔3个高分辨率孢粉序列(哈拉萨孜泥炭、娜仁夏泥炭和喀纳斯湖,图1)来探讨孢粉类型多样性对全新世“西风模态”的响应模式。本研究关注2 个科学问题:(1)不同海拔全新世时段孢粉类型多样性是如何演化的?(2)全新世孢粉类型多样性如何响应全新世气温和降水变化?
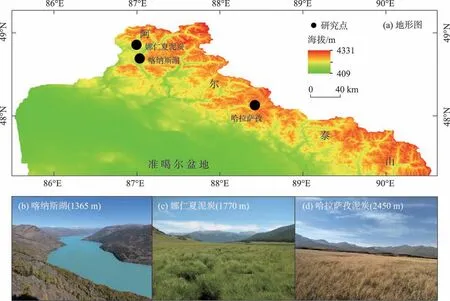
图1 研究区地形图(a)和研究点照片(b~d)Fig.1 Terrain of the Altai Mountains and photos of the studied sites
1 材料与方法
1.1 研究区概况
阿尔泰山位于西伯利亚泰加林和中亚沙漠边界,其独特的地形造就了特殊的植被景观。该区域平均年气温约-8~4 ℃,平均年降水量约300~500 mm[21]。由于冷季西伯利亚高压的发展,冷干的气候主导这一地区;在暖季盛行西风将北大西洋水汽输送至这一地区,导致阿尔泰山呈现降水向东减少、大陆度向东增加[22];现代植被也很好的反映了这一特征。具体说来,阿尔泰山北部、西部和西南部主要被暗针叶林所占据(包括冷杉、云杉和西伯利亚松)。进一步向东,西伯利亚松和西伯利亚落叶松盛行,在阿尔泰山的东南端高山草甸成为主导植被[22]。
阿尔泰山南坡地处其中端,全长约750 km,平均宽度约60~140 km。该区域形成了明显的垂直植被带[23],从低海拔到高海拔依次为:(1)荒漠草原带(海拔小于800 m),植被主要为由藜科、蒿属组成的荒漠类型,优势种为角果藜、豆科红车轴草、菊科刺头菊、蒿属、蓼科和禾本科等草本植物;(2)山地草原带(海拔800~1400 m),主要有蔷薇科绣线菊、麻黄属、禾本科、莎草科、豆科和唇形科等;(3)山地森林带(海拔1400~2300 m),该带植被类型多由西伯利亚落叶松与西伯利亚云杉构成混交林,或伴生有杨属和桦木属等;(4)高山草甸带(海拔2300~2700 m),主要有莎草科、禾本科、蔷薇科及石竹科等草本植物;(5)高山苔原带(海拔2700~3200 m),主要有苔藓和地衣[23]。
1.2 数据来源
本研究共选取阿尔泰山不同海拔3个高分辨率的典型孢粉序列来探讨孢粉类型多样性的演化。这3个研究点分别是哈拉萨孜泥炭、娜仁夏泥炭和喀纳斯湖(图2),基本情况如下:哈拉萨孜泥炭(48°07′N,88°22′E,海拔高度2450 m)位于高山草甸带。该钻孔有14个14C测年,分辨率约37 a,共分析了297个样品,每个样品鉴定数大于500个[16-17]。娜仁夏泥炭(48°48′N,86°54′E,海拔高度1760 m)位于山地森林带中。该钻孔有11 个14C 测年,分辨率约64 a,共分析了189 个样品,每个样品鉴定数大于300个[18]。喀纳斯湖(48°43′N,87°01′E,海拔高度1365 m)临近森林带下限。该钻孔有10 个14C 测年,分辨率约57 a,共分析了124个样品,每个样品鉴定数大于300个[19]。该3个研究点均覆盖全新世时段。
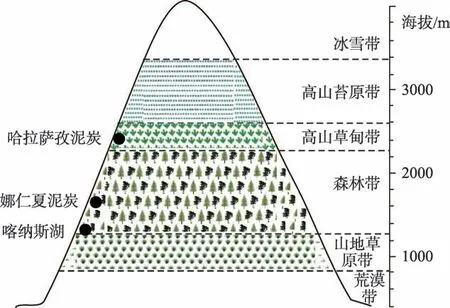
图2 阿尔泰山南坡植被带Fig.2 Vegetation belts on the southern slope of the Altai Mountains
1.3 孢粉多样性指数计算方法
本研究选择Shannon-Wiener 指数(H)来开展对孢粉类型多样性的信息度量。该指数是基于孢粉组合中随机抽样个体归属的种类的不定性[24]。当孢粉种类数量越大,则个体种类归属不定性就越大,表明孢粉组合的多样性就越高[24]。该指数的计算公式如下:
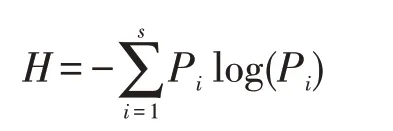
式中:log 可以选用以10 为底;S为孢粉种类数;Pi=ni/N表示第i个种类物种丰富度;ni表示每个孢粉分类单元的个体数;N代表群落中所有孢粉类群的个体总数。
为讨论孢粉序列中不同孢粉类型与多样性的关系,将孢粉进行了归类,共分为4类:泰加林组分、苔原组分、草原组分和荒漠组分[25-26]。前1 个称为木本组分,后3个统称为非木本组分。此外,为讨论全新世不同时段的孢粉类型多样性的变化,全新世(约11750 年前以来)可分为3 个阶段:全新世早期(约11750—8200年前)、全新世中期(约8200—4200年前)和全新世晚期(约4200年前—0)[27]。
2 结果与分析
2.1 哈拉萨孜泥炭
通过分析哈拉萨孜泥炭苔原、泰加林、草原和荒漠组分与Shannon-Wiener 指数的相关性可以看出,泰加林组分与Shannon-Wiener 指数的相关性最高(R2=0.21,P<0.01),其次是荒漠和苔原组分(R2=0.18,P<0.01;R2=0.10,P<0.01),草原组分与Shannon-Wiener指数相关性不显著(R2=0.00,P>0.05)(图3a)。研究结果表明,全新世期间泰加林和荒漠植被对哈拉萨孜泥炭孢粉类型多样性有贡献且泰加林的贡献最大,而草原植被对该泥炭孢粉类型多样性的贡献较小。
全新世期间,哈拉萨孜泥炭Shannon-Wiener 指数平均值为1.44,未表现出明显的变化趋势(图4a)。高于平均值的时段有约10600—8900年前、约8200—5000 年前、约4100—2300 年前和约1700—460 年前,其余时段Shannon-Wiener 指数均低于平均值。全新世时段最高值出现在约4100—2300 年前,最低值出现在约11600—10800年前。进一步对木本和非木本Shannon-Wiener 指数追踪后发现(图4a),非木本Shannon-Wiener 指数表现出与整个Shannon-Wiener指数一致的变化趋势,但木本Shannon-Wiener 指数可划分为3 个阶段:全新世早期Shannon-Wiener 指数较低,平均值为0.21;全新世中期Shannon-Wiener指数较高,平均值为0.33;全新世晚期Shannon-Wiener指数逐步下降,平均值为0.21。
2.2 娜仁夏泥炭
从图3b可以看出,全新世期间苔原、泰加林、草原和荒漠组分对娜仁夏泥炭孢粉类型多样性均有贡献,其中苔原组分与Shannon-Wiener 指数为负相关(R2=0.92,P<0.01)外,其余均为正相关。根据相关性分析结果(图3b),草原组分对孢粉类型多样性的贡献最大(R2=0.57,P<0.01),其次是泰加林(R2=0.53,P<0.01)和荒漠组分(R2=0.20,P<0.01)。
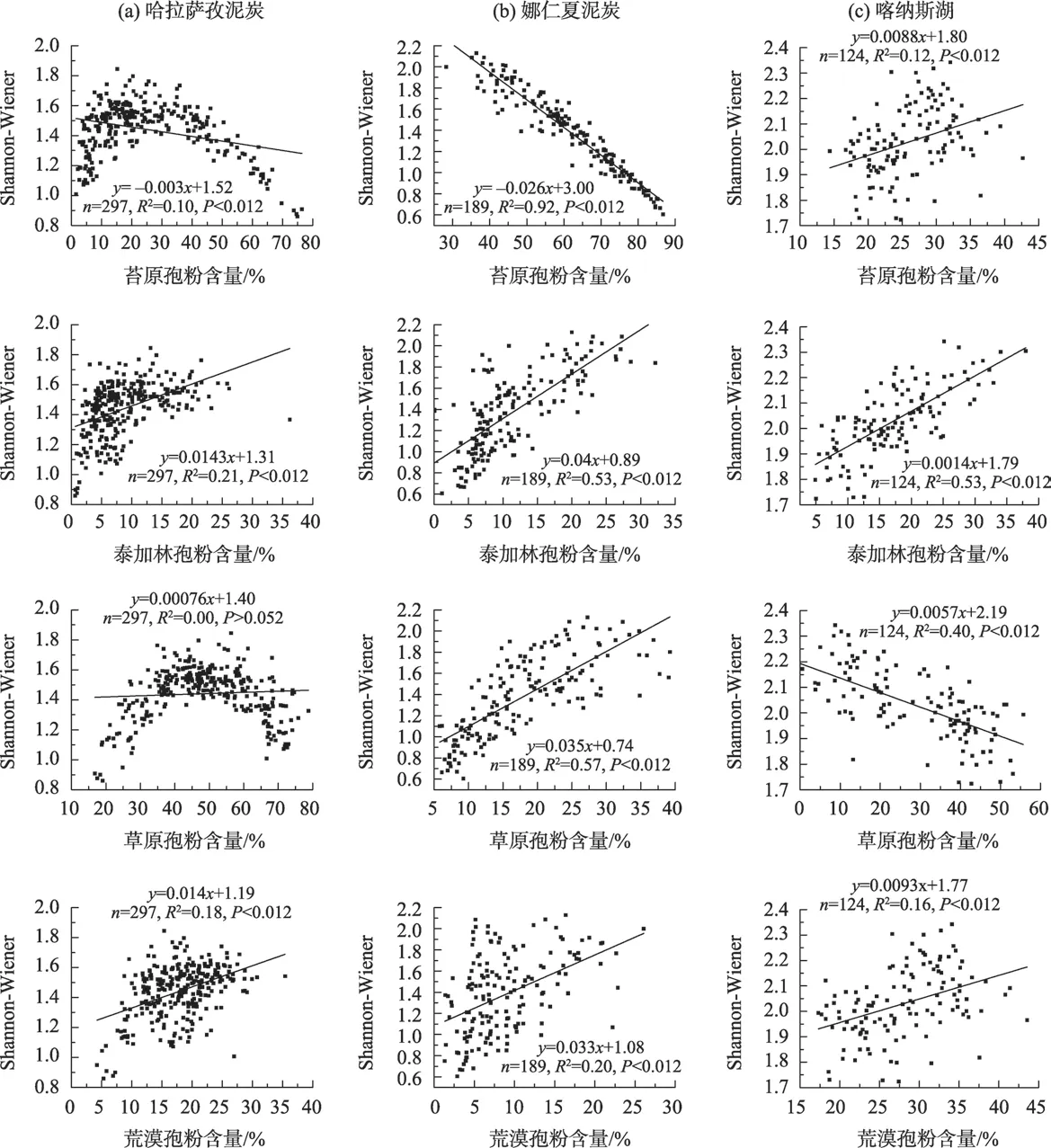
图3 不同研究点孢粉组分与Shannon-Wiener指数的线性关系Fig.3 Linear relationships between the pollen percentages and the Shannon-Wiener index from different sites
娜仁夏泥炭全新世Shannon-Wiener指数平均值为1.36(图4b)。与哈拉萨孜泥炭相比较,娜仁夏泥炭指数趋势变化明显,可以划分为3个阶段:全新世早期和晚期Shannon-Wiener 指数较高,平均值均为1.58,而全新世中期Shannon-Wiener 指数较低,平均值为1.05。就木本和非木本Shannon-Wiener指数而言,二者均表现出与整个Shannon-Wiener 指数一致的变化趋势,但非木本Shannon-Wiener 指数变化幅度大于木本的。
2.3 喀纳斯湖
通过分析喀纳斯湖苔原、泰加林、草原和荒漠组分与Shannon-Wiener 指数的相关性可以看出,泰加林组分与Shannon-Wiener 指数的正相关性最高(R2=0.53,P<0.01),其次是荒漠和苔原组分(R2=0.18,P<0.01;R2=0.12,P<0.01),草原组分与Shannon-Wiener 指数呈现显著的负相关性(R2=0.40,P<0.01)(图3c)。与哈拉萨孜泥炭类似,全新世期间泰加林、苔原和荒漠组分对喀纳斯湖孢粉类型多样性有贡献且泰加林的贡献最大。
喀纳斯湖全新世Shannon-Wiener指数平均值为2.03(图4c),且趋势可划分为3个阶段:全新世中期指数较高,平均值为2.19,而全新世早期和晚期指数较低,平均值均为2.07 和2.00。就木本和非木本Shannon-Wiener 指数而言,木本Shannon-Wiener 指数变化趋势与整个Shannon-Wiener 一致,但非木本Shannon-Wiener指数呈现显著的下降趋势(R2=0.42,P<0.01)。
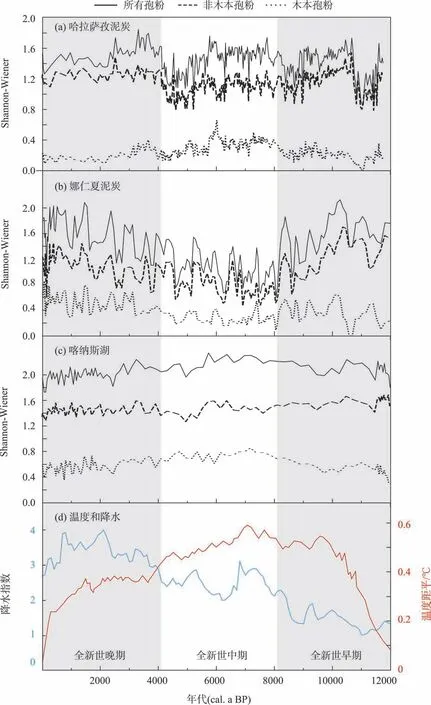
图4 阿尔泰山孢粉类型多样性的变化(a~c)与全新世气温[29]和降水[11](d)Fig.4 Changes in pollen taxa diversity in the Altai Mountains(a-c)and Holocene temperature[29]and precipitation changes[11](d)
3 讨论
通过分析阿尔泰山3 个研究点不同孢粉组分(苔原、泰加林、草原和荒漠)与Shannon-Wiener指数的关系可以看出,泰加林组分对不同海拔孢粉类型多样性均有显著的贡献,荒漠组分也表现出一致的正效应,而其他组分的表现略有不同(图3)。如草原组分对森林上限孢粉类型多样性的贡献很弱,对中海拔孢粉类型多样性是正贡献,而在森林下限孢粉类型多样性是负贡献。这些结果表明,利用Shannon-Wiener指数计算的孢粉类型多样性变化可以反映全新世时段阿尔泰山森林带的迁移变化特征。从图4a~图4c 可以看出,全新世时段3 个研究点的Shannon-Wiener 指数特征均未表现出一致的趋势,类似的不一致也出现在木本和非木本Shannon-Wiener 指数中。结合全新世气温[28]和降水[19]变化特征(图4d),进一步分析阿尔泰山不同海拔全新世孢粉类型多样性变化和植被带的迁移过程。
全新世早期(约11750—8200 年前),区域气候经历了变暖和干旱。哈拉萨孜泥炭和娜仁夏泥炭Shannon-Wiener 指数均较高,而喀纳斯湖Shannon-Wiener 指数较低。对木本Shannon-Wiener 指数来说,哈拉萨孜泥炭和喀纳斯湖指数均较低,而娜仁夏泥炭指数较高。可见,这一时期森林带主要占据中海拔地区(即娜仁夏泥炭所占区域),未扩张至哈拉萨孜泥炭和喀纳斯湖所在区域。随着气候的逐渐变暖,森林上限逐渐上移,且随着湿度的逐步增加,森林下限开始下移。该森林带的迁移使得哈拉萨孜泥炭和喀纳斯湖木本孢粉类型多样性逐渐增加,而娜仁夏泥炭位于中海拔的森林带内,木本孢粉类型多样性持续保持在高值。
全新世中期(约8200—4200年前),区域气候最温暖且湿度增加。哈拉萨孜泥炭和喀纳斯湖Shannon-Wiener 指数处于高值,表明孢粉类型多样性较为丰富;而娜仁夏泥炭Shannon-Wiener指数下降,表明孢粉类型多样性随之下降。这一特征表明,在温暖变湿背景下,森林带持续扩张,上、下限逐步向上和向下移动,使得森林逐步占据原先不位于森林带的区域(即哈拉萨孜泥炭和喀纳斯湖所在区域),从而引起该区域木本孢粉类型多样性的增加,进而导致整个时段孢粉类型多样性较高。但是在此气候背景下,娜仁夏泥炭孢粉类型多样性记录了全新世时段的最低值,该低值在木本和非木本Shannon-Wiener 指数中均有体现。该时段明显下降的孢粉类型多样性可能归咎于两方面的原因:(1)温暖变湿的气候不仅促使森林带的扩大,而且引起森林带内的云杉和落叶松面积增加[15-18],进而导致其他树种面积和均匀度的下降[11],最终引起森林带木本多样性的下降;(2)森林带面积的增加势必会引起林间草地面积的下降,进而引起森林带非木本多样性的减少。
全新世晚期(约4200 年前—0),区域气候最湿润且气温下降。哈拉萨孜泥炭和娜仁夏泥炭Shannon-Wiener 指数处于高值,表明该时段孢粉类型多样性丰富;而喀纳斯湖Shannon-Wiener指数下降,表明该时段孢粉类型多样性有所下降。对于木本孢粉来说,哈拉萨孜泥炭Shannon-Wiener 指数缓慢下降,表明在变冷变湿的背景下森林带开始向下移动,自然会导致中低海拔孢粉类型多样性的增加。但喀纳斯湖Shannon-Wiener指数所记录的多样性有所下降,这一变化与笔者所理解的在变冷变湿背景下森林带持续向更低海拔移动所不同。造成这种差异的原因可能有:(1)气候的持续变冷使得向低海拔迁移的泰加林面积减少,进而引起木本多样性的下降;(2)全新世晚期低海拔人类活动的持续加强和火灾的频繁发生也会引起孢粉类型多样性的减少[29]。
综上所述,在全新世气温和降水的综合影响下,阿尔泰山植被带的变化呈现出独特的格局:全新世早期气候变暖和干燥,位于中海拔的森林带开始向上迁移,使得高海拔孢粉类型多样性增加。全新世中期气候温暖且湿润,森林带的持续向上和向下迁移使得哈拉萨孜和喀纳斯湖孢粉类型多样性较高,但因中海拔以云杉属孢粉和落叶松属孢粉占优势的林带扩张,导致泰加林物种均匀度下降,娜仁夏泥炭周边观察到的孢粉类型多样性较低。这一观察结果也得到了临近天山孢粉类型多样性变化和表土孢粉结果的支持[11-12]。全新世晚期气候变冷且更湿润,森林带上限向下移动,使得中海拔孢粉类型多样性显著提高。这些结果表明,当气候温暖干燥时,森林带沿海拔向上移动以获得更多的水分,反之亦然。同时,森林带通过调整林带宽度来响应温暖湿润的气候,进一步使森林带向下移动[11]。
从阿尔泰山不同海拔全新世孢粉类型多样性特征可以看出,娜仁夏泥炭所在的中海拔地区植被多样性的变化幅度最大(图4a~图4c),说明森林带所在区域是对气候响应最敏感区,也是最脆弱区。考虑到湿度敏感类群的气候生态位空间及其有限的生存空间,水分流失对相对狭窄的森林带的威胁最大。当气候干燥超过其阈值时,森林带中的一些类群可能会消失。例如,作为世界多样性热点之一的东安第斯山脉狭窄的植被带对气候变暖的脆弱性很大,因为预期的气候变化速度可能会在数百年内将狭窄分布的植物类群移出气候生态位空间[30]。值得关注的是,在当前变暖背景下,研究区面临干旱加剧的风险,干旱强度和持续时间表现出不断增加趋势[31]。可见,将来气候变暖背景下可能会对干旱区森林带植物类群造成难以估量的威胁,后续需要加强山地森林带的保护和维持,特别关注对湿度敏感的植物物种。此外,尽管因受孢粉鉴定精度、产量、传播能力、代表性与沉积环境不同等的影响,使得利用其恢复古植物多样性可能存在一定的偏差[24,32]。但孢粉分析仍然是一种具有巨大潜力来重建植物多样性的方法,可以为我们研究山地植被带对全新世气候变化的响应提供可参考的关键证据[33]。
4 结论
通过分析阿尔泰山基于孢粉序列记录的不同海拔孢粉类型多样性变化发现,在暖干的全新世早期,森林带上限开始上移,对高海拔孢粉类型多样性的贡献逐步增加。在暖湿的全新世中期,森林带变宽,向高、低海拔同时扩张,使得高、低海拔区木本孢粉类型多样性明显增加。但森林带内因物种均匀度的下降而孢粉类型多样性明显减少。在冷湿的全新世晚期,森林带上限开始下移,导致森林带孢粉类型多样性增加。可见,全新世气温和湿度(或降水)共同影响森林带的迁移,进而引起不同海拔孢粉类型多样性的变化。
猜你喜欢
军事文摘(2022年14期)2022-08-26
绿色中国(2022年2期)2022-02-20
散文诗(2021年22期)2022-01-12
下一代英才(2021年5期)2021-08-05
当代化工(2019年3期)2019-12-12
环球时报(2019-05-23)2019-05-23
森林与人类(2018年12期)2018-03-20
科技视界(2016年7期)2016-04-01
哈尔滨理工大学学报(2014年1期)2014-06-23
百科知识(2009年4期)2009-03-07
