
解钾菌及其释钾微观机制的研究进展*
2022-06-09 01:16朱晓艳陈国炜万小铭
土壤学报 2022年2期
韩 苗,朱晓艳,陈国炜,万小铭,王 钢†
(1. 中国农业大学土地科学与技术学院,北京 100083;2. 合肥工业大学土木与水利工程学院,合肥 230009;3. 中国科学院地理科学与资源研究所,北京 100101)
解钾菌(potassium-solubilizing bacteria,KSB)通常是指从土壤中分离出来的一类能够分解含钾矿物的细菌,它能将矿物钾转化为速效钾而供植物直接利用,对作物产量的提高和土壤肥力的改善具有重要作用。解钾菌也被称为硅酸盐细菌或钾细菌,其分离及释钾能力的评定通常是在以钾长石、白云母或黑云母作为唯一钾源的改良Aleksandrov培养基上进行的。解钾菌的植物促生、土壤培肥以及环境友好等特点,促使对其的相关研究受到国内外学者的广泛关注。
土壤中解钾菌种类不一、功能多样。解钾菌不仅能够通过分解含钾矿物来增加土壤速效钾含量,而且还可通过活化土壤中的生物酶类(如土壤脲酶、过氧化氢酶和蔗糖酶等)并使其活性处于适宜范围,从而促进动植物残体及凋落物的分解与转化,进而提高土壤中其他营养元素的有效性。此外,解钾菌还能够通过分泌吲哚乙酸(indole-3-acetic acid,IAA)、1-氨基环丙烷-1-羧酸(1-aminocyclopropane-1-carboxylate,ACC)、赤霉素(gibberellin,GA)、铁载体(siderophore)和脱氨酶(deaminase)等物质来促进作物生长,同时对植株抗逆性的提高也有关键作用。需注意的是,蒋先军等的研究表明促进作物生长的解钾菌代谢产物存在适宜浓度范围。
近十年来,学者们采用室内盆栽试验和大田试验对接种解钾菌的效果进行了大量研究,发现接种确实能显著提高土壤肥力,提升作物产量和品质。Prajapati 和 Modi的研究结果表明在水培条件下能显著增加黄瓜的根长、开花结果率、果实成熟度、钾含量和叶绿素含量。Sugumaran 和Janarthanam通过微缩试验发现使用从印度土壤中分离得到的接种花生,可提高土壤钾含量、植株产量和种子含油量。此外,一些田间试验也证明接种解钾菌可以提高茄子产量,烟草生物量、养分含量、钾吸收量和叶片品质以及水稻株高、茎粗、根长、叶面积和生物量干重。此外,学者们对解钾菌作为生物钾肥的施用效果也进行了多次试验,结果表明生物钾肥能提高土壤有效氮、磷、钾含量以及作物块茎产量和品质。因此解钾菌作为生物肥料用于农业改良确实可以有效减少化肥使用,促进生态友好型作物的生产。
近年来,大量学者聚焦于研究解钾菌与含钾矿物或其他植物促生根菌(plant growth-promoting rhizobacteria,PGPR)结合施用对作物生长的影响。许多研究表明将含钾矿物与解钾菌结合作为生物肥料施用,可能是一种替代传统化肥、降低生产成本的有效手段。Sarhan 和Abd El-Gayed研究了将长石作为钾源并结合一些生物刺激物(解钾菌、腐殖酸和农家肥)的施用对棉花产量和收获后土壤性状的影响,结果表明解钾菌分别与长石、腐殖酸或农家肥结合施用对棉花产量的影响更显著。同时,大量试验证明解钾菌与解磷菌、丛枝菌根真菌或固氮菌等PGPR 共同接种更能够促进作物生长发育,增加土壤养分供给。
可见,前人主要围绕解钾菌宏观方面开展了大量研究,然而对影响解钾菌释钾能力的关键——解钾菌和矿物间微观互作机制——了解不多。本文主要从单个浮游解钾菌细胞入手,阐述其在含钾矿物固-液界面的聚集、吸附及生物膜形成和循环,直至最终解钾功能的发挥,并总结了影响该微观过程的关键因素,且详细介绍了与该过程相关的前沿科学技术,旨在为解钾菌微观释钾机制研究提供理论和技术参考。
1 解钾菌宏观解钾机制
在过去的几十年里,研究者们从宏观角度详细阐述了解钾菌解钾机制,主要的观点有酸解、酶解和多糖络合溶解等,该类结论均是学者们基于解钾菌能够分泌胞外聚合物(extracellular polymeric substances,EPS)而得出的。然而事实上,除细菌“EPS 分泌量和组分”影响其解钾能力外(不同细菌EPS 分泌种类和含量差异较大,表1),细菌运动性也是影响解钾能力的核心因子,运动性通过调控细菌在矿物表面的黏附量和生物膜循环影响矿物释钾能力。无机酸和有机酸的产生是含钾矿物风化的主要机制。解钾菌产生的有机酸和无机酸可以通过降低其周围环境的pH,直接促进含钾矿物的风化,导致可交换性钾的缓慢释放,提高环境速效钾含量。另一方面,含有羟基和羧基的有机酸易与矿物中的金属离子发生反应形成金属-有机络合物,从而破坏矿物的结构,促进矿物中钾的释放。Groudev于1987 年首次发现胞外多糖也能够提高解钾菌解钾能力。胞外多糖中含有具有络合作用的官能团,它们能与矿物中的Si、Al和K络合从而促进含钾矿物的分解。同时,Liu 等的研究表明,菌株产生的多糖能够吸附有机酸并附着在矿物表面,从而导致矿物表面微区域有机酸浓度的增高。除此之外,荚膜多糖还能够通过吸附SiO来影响固相和液相的平衡,从而加速SiO和K的溶解。除了有机酸和多糖外,解钾菌分泌的蛋白质(或酶)也参与含钾矿物的分解。一方面,蛋白质中含有具有络合作用的官能团(氨基、羧基等);另一方面,在矿物存在的条件下,细菌能够通过相关功能基因组的表达来实现蛋白质的调节,从而控制微生物对矿物的风化。例如,Chen 等在使用双向凝胶电泳(two dimensional electrophoresis,2-DE)研究胶质芽孢杆菌分解磷矿石机理时,发现处理组(含矿物)菌体细胞的蛋白点表达量与对照组(不含矿物)相比,共有差异蛋白点496 个,其中,有207 个蛋白质点为新合成,214 个蛋白质点表达上调。这表明细菌在风化矿石时能够激活或增加某些蛋白的表达,这些蛋白点很可能与细菌的代谢调控、增生和信号转导通路等有关。

表1 土壤中解钾菌性能特征(胞外分泌物、革兰氏染色反应和运动性)Table 1 Characteristics of soil potassium-solubilizing bacteria(EPS secreted,gram reaction,and motility)
2 解钾菌微观解钾机制
解钾菌与含钾矿物的直接接触对于解钾过程至关重要。解钾菌可通过分泌多糖黏附于含钾矿物表面从而形成特殊的微环境来促进其解钾功能的发挥,同时,其生长过程中所带来的物理破坏作用也对含钾矿物的风化非常关键。但直接接触并不是必需过程,孙德四等研究了环状芽孢杆菌在细菌-矿物直接接触与间接接触(微孔滤膜将细菌与矿物隔离)两种模式下对铝土矿风化分解行为的影响,发现细菌及其代谢产物能够通过直接和间接作用共同促进铝土矿的分解,但直接作用的强度明显大于间接作用。此外,满李阳等的研究结果也进一步证实了这一观点,并提出含钾矿物中钾的释放在接触前期主要受间接作用的影响,而在接触后期主要受直接作用的影响。
间接接触:解钾菌分泌的无机酸、有机酸、多糖以及蛋白质等物质能够通过降低环境pH、质子交换、络合或螯合等作用促进含钾矿物的分解。而从微观角度解释这些物质分泌机理的研究还处于空白阶段,目前作者设想该过程可能是由细菌的竞争(bacterial competition)或群体感应(quorum sensing)导致。在养分缺乏的环境中,细菌的竞争机制能够促使其向外分泌特殊的代谢产物、酶或胞外囊泡等,或通过占用其他菌株的EPS,来与竞争者争夺养分或抑制竞争者的生长。例如,细菌可以通过向外分泌铁载体(特殊的代谢物)的形式占据更多的铁素资源,从而减少其竞争者的铁供应,铁载体不仅可与溶液中的铁离子形成络合物,而且还可以将一些深埋于矿物或其他螯合物中的铁离子拽出来,从而促进菌体对铁的吸收;细菌的运动性及其分泌的EPS(如黏附素、鼠李糖脂和脂肪酸顺-2-癸二酸等)有助于其占据生长空间,通过促使生物膜的形成来提高竞争力。此外,细菌能通过向外分泌自我诱导物(autoinducers,即信号分子)来进行群体感应,当信号分子的浓度达到一定阈值时,菌体相关基因的表达就会被启动以适应或改变周围环境。
直接接触:解钾菌单个细胞(single-cell)可通过主动扩散(自主运动)或被动运输(水体剪切流、重力沉降等)的方式到达矿物界面,形成生物膜。上述过程对解钾菌解钾作用的发挥和解钾效率至关重要。细菌往往生活在低雷诺系数(Reynolds numbers,约10)的水环境(acqueous environment)中,其扩散能力可以由扩散系数(diffusion coefficient,D)来表征,能动性细菌(如)在水中的扩散系数大约为4×10m·s,而不具有运动能力的细菌(如缺失鞭毛的)的扩散系数大约为3×10m·s,远小于运动细菌。不具有运动能力的单个细菌在水中的运动主要依靠布朗运动,其扩散系数的值相当于与其等大小的惰性粒子的值,计算公式如下:

式中, <>为均方位移( the mean-square displacement,MSD),为时间(time)。
而同等大小的惰性粒子的布朗扩散系数可由斯托克斯-爱因斯坦方程(Stokes-Einstein equation)得出:

式中,为波尔兹曼常数(Boltzmann constant),为绝对温度(Kelvin temperature),为黏度(viscosity),a 为粒子半径(particle radius)。目前实验已证明用以上两个公式计算不运动细菌的扩散系数,其值相差不大,约为10m·s。
此外,Lovely 和Dahlquist于1975 年首次给出了基于单细菌细胞运动速度计算扩散系数的方法:

式中,为运动速率(speed),为细胞单个运动轨迹的平均持续时间(the mean duration of a single run trajectory),为旋转角度余弦的平均值(the mean cosine of the angle between successive trajectories),其测量需要细菌的三维运动轨迹。同时该公式的成立需要很多假设条件(如前进方向为直线、所有轨迹速度相同等),与细菌实际运动状况存在差异,还需要我们进行更多的深入研究。
上述公式是基于单个细菌细胞对细菌扩散的计算。此外,仍可利用菲克方程(Fick’s equations)计算基于细菌种群水平的扩散系数,
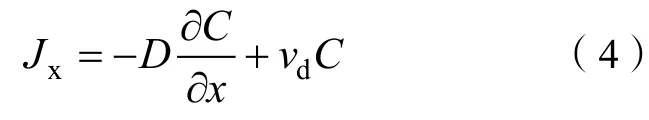
式中,为轴的通量(flux at point),为细菌种群浓度(bacterial population concentration),为轴的距离,为细菌在轴的速度(the velocity in the +direction)。式(4)为菲克第一定律,适用于浓度恒定的稳态扩散(steady-state diffusion),与均不随时间变化,即每一时刻从前边扩散来多少细胞,就向后边扩散走多少细胞,浓度不随时间变化。实际上,自然条件下大多数扩散过程均是在非稳态条件下进行的,
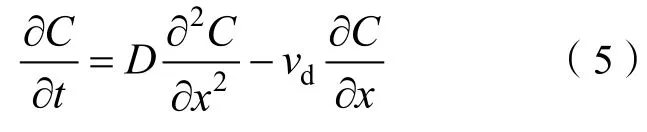
式中,为细菌种群浓度,其值随扩散时间变化。式(5)为菲克第二定律,适用于浓度随时间变化的非稳态扩散(none steady-state diffusion)。
同时,由式(2)的计算结果可知养分分子在水中的扩散系数大约为10m·s,明显大于细菌随机运动的扩散系数。同时,在具有一定化学浓度梯度的溶液中,细菌的扩散系数会显著增加。这说明溶液中的养分分子更容易到达固-液界面,从而诱导细菌朝固-液界面进行趋化运动并形成生物膜。这很可能是土壤中99%的细菌都吸附于生物或非生物表面(如土壤颗粒、根系等),以生物膜的形式存在并发挥作用的主要原因。单个浮游解钾菌微观释钾过程可以大致认为是单个浮游细菌趋向矿物表面运动聚集、吸附及生物膜形成和循环的过程,这个过程可大致分为四个阶段(图1e):
(1)聚集阶段:大量单个浮游细菌细胞迁移至矿物-水界面并被其捕获;
(2)可逆附着阶段:细菌细胞感知固-液界面,并在“物理-化学综合作用力”下可逆附着到固-液界面上;
(3)不可逆附着阶段:细菌细胞由对固-液界面的可逆附着向不可逆附着转移;
(4)生物膜形成与循环:不可逆黏附在固-液界面上的细胞大量分裂增殖,直至形成成熟的生物膜,最后生物膜最外层的活细胞脱落并开启新循环。
2.1 聚集阶段
在聚集阶段,细菌的运动性(趋化运动或随机运动)及水体的剪切流发挥着主导作用。在适宜的环境条件下,具有游动(swimming)能力的细菌能够借助自身鞭毛马达的旋转在溶液中进行运动,细菌根据自身鞭毛类型(生长位置和数量)进化了多种运动模式。如周生鞭毛菌、和等执行的前进-翻滚(run-tumble)运动(如图1a)。从细菌后面来看,鞭毛逆时针旋转拧成一束从而推动细胞向前运动,而细胞体顺时针旋转以保持扭矩平衡,当某些鞭毛顺时针旋转或散开时,细菌前进方向就会改变,所选择的方向取决于束相对于细胞体的变化。而单根极性鞭毛菌由于不能通过鞭毛束的散开随机选择方向,其运动性更为复杂。早期,学者们认为只有布朗运动可以帮助单根极性鞭毛菌完成再定位,这是因为布朗运动能够对小型单鞭毛细菌的运动产生影响,同时改变它们的前进轨迹和后退时的细胞取向,如执行的前进-停止(run-stop)运动(如图 1c)以及和(PAO1)执行的前进-后退(run-reverse)运动(如图1d)。目前,关于PAO1 的运动模式还存在争议,Qian 等的研究表明其运动遵循run-reverse-turn 模式,但Stocker则认为 PAO1 的运动很可能按照 runreverse-flick 模式。近年来,研究者们发现单根极性鞭毛菌也可以通过鞭毛离轴变形(旋转轴心偏离细胞体轴心)来改变其前进方向,如执行的前进-后退-轻弹(forward- reverse-flick )运动(如图1b)。然而,在实际情形中,细菌进行随机运动的机会并不多。一方面,环境中的水流会影响细菌的运动方向,当剪切速率不大于10 s时,鞭毛的推动力仍然在细菌运动中发挥主导作用,而当剪切速率大于10 s时,剪切流就会通过影响细菌细胞体-流体界面的作用力来主导细菌的运动方向。另一方面,环境中常常存在化学浓度梯度,而细菌细胞膜上的跨膜蛋白能感知化学浓度变化,并将信号传递给鞭毛,促使其通过调节自身旋转方向进行具有偏向性的趋化运动。矿物-水界面由于表面张力,常常吸附大量养分分子(钾素及其他养分),细菌的趋化性会驱动细菌主动向矿物-水界面靠近,以便从中获得代谢过程所需要的电子受体、钾素及其他营养物质,最终使细菌大量聚集到矿物-水的近界面区域内。研究发现,相对于随机运动而言,细菌趋化运动会使细菌在沿着高养分浓度梯度上的运动时间增加。相对于游动细菌而言,不具有运动能力的细菌到达矿物-水近界面区域的主要方式有布朗运动、重力沉降和剪切流。除剪切流和养分浓度梯度外,环境溶液的离子强度、pH、温度及矿物表面物理化学特性(粗糙度、所带电荷性质及数量等)均会影响细菌在矿物-水界面的聚集。因此,我们推测上述物理-化学因素均是影响解钾菌在含钾矿物表面聚集的关键,且聚集过程又是影响解钾菌与含钾矿物接触并在其表面形成生物膜的关键过程。
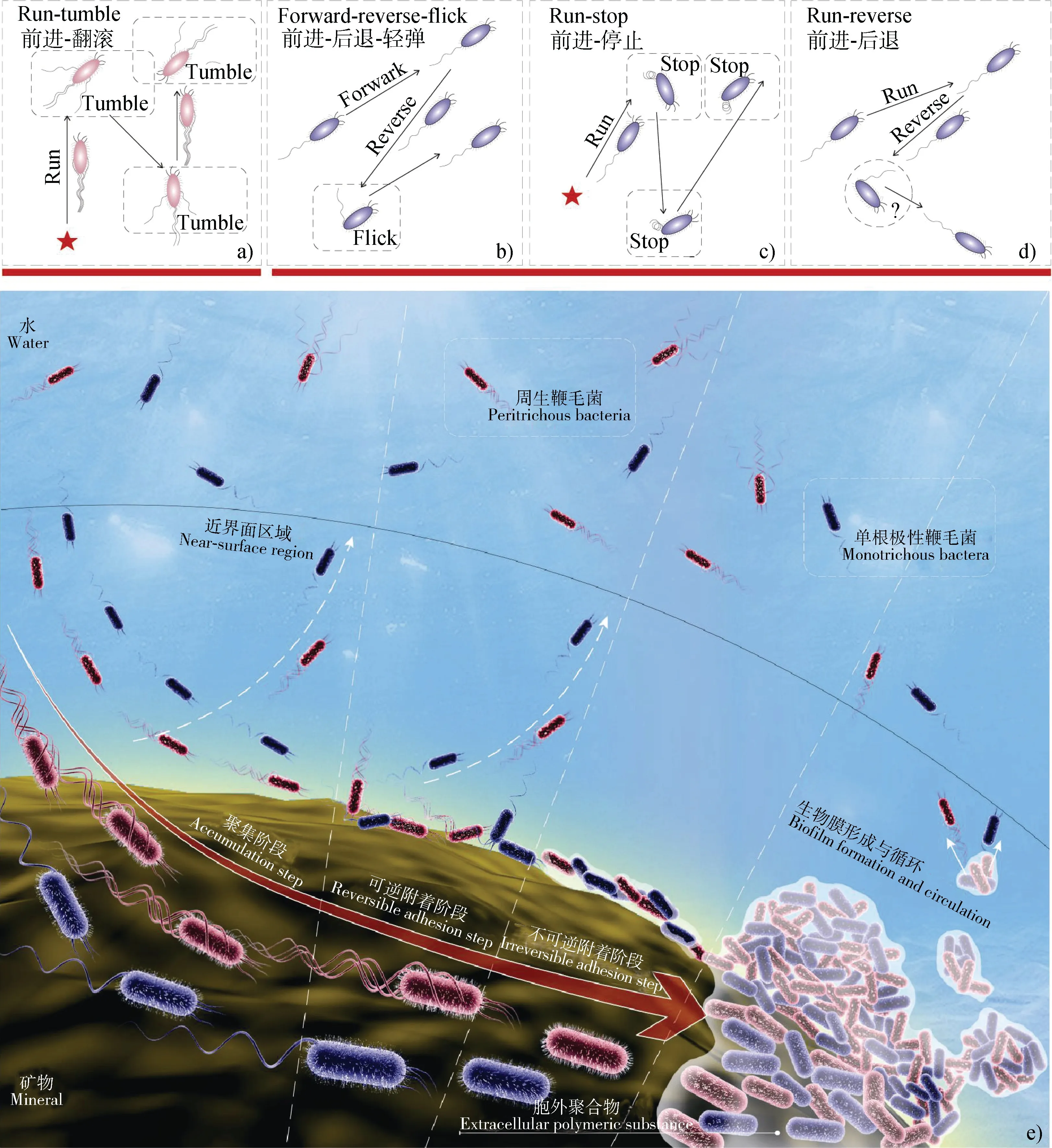
图1 细菌在矿物表面生物膜形成过程Fig. 1 Biofilm forming process on mineral surfaces
2.2 可逆附着阶段
在可逆附着阶段,细菌到达矿物-水近界面区域(距离界面20 μm 以内的区域,不固定,随细菌和界面特征而变化)内,虽然自身趋化性整体效果减弱,但却被细菌与界面间的相互作用力(水动力学相互作用力和DLVO 力等)捕获而难以逃离。Molaei等指出在近界面约20 μm 区域内,的tumble行为降低了50%,且再定位的方向主要局限于与界面平行的方向上,这表明这时tumble 并不能使细菌挣脱界面的束缚。当细菌游动时,鞭毛的运动会干扰周围液体从而形成流动,而固液界面(尤其是距离细菌几个身体长度范围内的界面)的存在又会影响这种流动,从而反过来影响细菌本身的运动,这种机制被称为水动力学相互作用,可以大大促进细菌在矿物-水界面的定殖。当一个非圆形细胞从自由溶液向固液界面靠近时,由于作用于细胞体和鞭毛束下部的黏性曳力大于作用于两者上部的曳力,细菌在界面上方产生高曲率曲线运动,同时,由于其非球形而产生的曳力会使细胞保持一个相对于界面的特定倾角,因此细菌常在近界面区域以一定的角度向下倾斜着做曲线运动并逐渐向界面旋转靠近。由于细菌的形状、大小、鞭毛马达的扭矩特征、鞭毛丝的变形及其与界面间的作用力在细菌的不同种类间存在差异,其旋转方向和速度也会因此存在不同。
当细菌与界面相距大约9 a(a 为细胞体半径)时,界面与细胞体及胞外运动附属物(鞭毛和Ⅳ型菌毛)间的摩擦力变得不可忽视,摩擦力会通过影响细菌的运动速度来增加细菌在界面的停留时间,从而提高细菌黏附到界面的可能性。随着距离的进一步缩短,细菌鞭毛和胞外Ⅳ型菌毛(type Ⅳpili,TFP)开始感知到界面的存在并对界面产生吸附作用,其中TFP 通过自身的收缩或拉伸介导的细菌在矿物表面的运动(walking 和crawling)对细菌充分利用矿物表面空间非常关键。另一方面,与界面的接触会抑制鞭毛马达的旋转和TFP 的收缩,从而促使细菌在界面的附着。而当细菌与矿物-水界面的距离进一步缩短至大约 50 nm 时,DLVO(Derjaguin-Landau-Verwey-Overbeek)力(范德华相互吸引力和静电相互排斥力)开始发挥作用,细菌细胞被DLVO 力捕获,但这个距离并不固定且取决于细胞和界面的物理化学特征。当细菌与界面距离较远时,起决定作用的是范德华相互吸引力,随着距离的逐渐缩短,细菌细胞的双电层与界面的电子层(均常带有负电荷)重叠产生了静电相互排斥力。当细菌细胞逐渐接近界面时会存在能量势垒(energy barrier),在能量势垒外存在一个浅层二次能量最小值(the secondary energy minimum),从界面到二次能量最小值的距离通常在几纳米或十几纳米(不固定,取决于溶液离子强度)内,这个距离是细菌与界面的最佳距离。在细菌黏附的第一步,细菌细胞通常通过自身的运动性或布朗运动到达这个位置,并可逆地附着在界面上。目前,关于DLVO 力在细菌可逆附着阶段的作用仍存在争议,Vigeant 等认为可逆吸附的细胞距离界面的距离太远,并不能用DLVO 力来解释,DLVO 力只能用于描述细菌从可逆附着到不可逆附着阶段。
Li 等发现当细菌在界面附近运动时,布朗运动的作用会被放大,这是由于布朗运动会显著改变细菌和固-液界面间的距离,进而影响两者间的水动力学作用和DLVO 力等,从而促使其对界面的附着。这些物理-化学作用力很可能也是非游动细菌(nonswimming bacteria)和失去趋化感知能力的游动细菌(non-chemotactic swimming bacteria)在矿物-水界面附着的主要原因。总而言之,在细菌细胞到达矿物-水近界面区域(大约<20 μm)后,水动力学相互作用力和布朗运动首先起作用;而随着细胞与固-液界面间距离的缩小(<9 a),水动力学相互作用力、细菌与界面间的摩擦力和布朗运动起作用;当细胞进一步靠近界面(<50 nm)时,由于空间有限细菌鞭毛无法旋转,且细菌很可能感知到固-液界面并停止鞭毛旋转从而黏附到界面上,细菌与固-液界面间的水动力学相互作用力消失,此时,DLVO 力、鞭毛和菌毛对于界面的黏附作用和布朗运动发挥作用。这些力都会增加细菌细胞可逆附着到矿物表面的可能性。而在此阶段中,环境温度、溶液离子强度、pH 和解钾菌胞外运动附属物的类型等均会影响解钾菌的可逆附着。
2.3 不可逆附着阶段
在不可逆附着阶段,细菌与矿物表面距离达到几十纳米,细菌由可逆附着向不可逆附着转移。此时,范德华相互吸引力和疏水相互作用(hydrophobic interactions)起主导作用。疏水相互作用力源于水分子间氢键聚合的键能,细菌与矿物间的疏水作用为引力,有利于两者的吸附。而在这个过程中,细菌开始向外分泌EPS,如DNA、蛋白质、脂类、脂多糖等,这些物质可以将细菌和界面连接起来,促进两者间的不可逆附着。此外,由于鞭毛和TFP 马达受到抑制,细菌会被促使着产生一些其他物质,如分泌的纳米黏合剂holdfast 和大多数细菌均能够产生的胞内环二鸟苷单磷酸(cyclic diguanylate monophosphate,c-di-GMP),从而使细菌在界面的附着更牢固。除此之外,细菌鞭毛和TFP 对于界面的吸附都会促进其不可逆附着。环境温度和离子强度、解钾菌和含钾矿物表面的离子性质和强度及解钾菌运动附属物的类型等都会对这个与之对应的解钾过程产生影响。
2.4 生物膜形成与循环阶段
在生物膜形成与循环阶段,细菌不断生长增殖并分泌大量EPS,在矿物表面形成有组织的生物膜。而当生物膜成熟之后,最外层的活细胞脱落并开始新生物膜形成的循环过程。在这个过程中,解钾菌分泌的EPS 通过上述宏观解钾机制促进含钾矿物的溶解以及细菌对营养元素的吸收。因此,解钾菌分泌EPS 的种类和数量对于解钾过程来说至关重要。
解钾菌的运动性在生物膜形成前期极其重要,EPS 在生物膜形成后期具有不可替代的地位,而解钾菌宏观解钾机理主要是其EPS 的作用,这说明解钾菌生物膜的形成过程(微观解钾机制)与解钾菌宏观解钾过程密不可分,息息相关。上述四个阶段清晰地介绍了单个浮游解钾菌细胞到达含钾矿物界面并在其表面形成生物膜,以及发挥解钾机制的具体过程及其影响因素,同时还强调了解钾菌自身运动性、水动力学相互作用、DLVO 理论和布朗运动在该过程中的重要性。从该视角出发能为解钾菌解钾机制的研究提供一个新方向。
3 解钾菌解钾机制研究的前沿技术和相关理论
越来越多的前言技术随着科学发展应运而生,显微成像技术(数字全息显微镜、全内反射荧光显微镜、激光共聚焦显微镜、原子力显微镜和电子显微镜等)和平面光极技术等的交叉、灵活应用对解钾菌解钾机制的深入研究具有重要推动作用(图2)。

图2 解钾菌微观机制研究的相关技术图Fig. 2 Technical drawings related to researches on microscopic mechanisms of bacteria solubilizing potassium
光学显微镜:(1)数字全息显微镜(digital holographic microscopy,DHM)主要是以再现技术和光学显微技术满足折射或者透视物体3D 轮廓,从而获取被测物体的振幅信息和相位信息,进而完成数字三维重构,其轴向分辨率为0.6 nm,横向分辨率为300 nm(取决于物镜的数值孔径),测量范围可达4 mm(1.25 倍)。该显微镜目前已应用于无荧光条件下生物样本和活体细胞的三维运动轨迹观测、三维形貌测量和变形测量。在解钾菌解钾机制的研究中,DHM 可以用于观测解钾菌由浮游状态运动至含钾矿物界面附近并在其界面形成微菌落的3D 运动轨迹,同时也能测定该过程中含钾矿物表面形态的变化。(2)全内反射荧光显微镜(total internal reflection fluorescence microscope,TIRFM)是利用全内反射产生的隐失波照明样品,使照明厚度限定在几百纳米厚(200 nm,甚至是100 nm)的光学薄层内的显微术。该显微镜可以用于活细胞单分子成像、生物大分子在界面吸附行为的研究和酶反应的观测等。TIRFM 能够用于测定解钾菌在含钾矿物界面附近几百个纳米范围内的运动特征,且能够观察到运动特征随着与界面距离的改变而产生的变化情况。(3)激光共聚焦扫描显微镜(confocal laser scanning microscope,CLSM)是一种集显微技术、高速激光扫描和图像处理于一体的新型光学显微术,放大倍数可达32 000 倍,分辨率可以达到普通显微镜的1.4 倍。该显微镜可用于测定矿物三维结构,表征细菌在界面形成的生物膜,获得高分辨率的细菌彩色荧光图像。结合细胞染色或荧光标记,CLSM 可以用于观测解钾菌的实时运动变化和在含钾矿物表面形成的生物膜形貌变化以及矿物表面粗糙度的实时变化。(4)原子力显微镜(atomic force microscopy,AFM)是用一端固定而另一端装有纳米级针尖的弹性微悬臂来检测样品表面结构三维形貌或原位定量测量界面和细菌细胞之间界面力的显微术。该显微镜可以用于测量含钾矿物表面粗糙度以及解钾菌与含钾矿物间的DLVO 力等,对于理解生物膜的形成以及解钾菌对于特定矿物表面的亲和力至关重要。
电子显微镜是使用波长极短的电子束做光源,电磁场做透镜,分辨率可达0.1 nm 的电子光学仪器。电子显微镜又分为透射电子显微镜(transmission electron microscope,TEM)和扫描电子显微镜(scanning electron microscope,SEM),前者是用电磁透镜对透射电子聚焦成像,而后者是利用末级透镜上面的扫描线圈(其作用是使电子束偏转,在样品表面做有规则扫描)来调节成像。由于胞外运动附属物一般极细,鞭毛直径约几十纳米,菌毛直径几纳米到几十纳米,只能通过电子显微镜才能观察到。同时,电子显微镜也可以用于矿物表面粗糙度和细菌在矿物界面生物膜形成情况的测定。
平面光极(planar optode,PO)是一种基于荧光分析原理,利用光学数字成像实时测定待测分析物(pH、O、CO、NH、温度等)二维浓度的技术。该仪器采用简单低成本的光电子器件,具有响应速度快、精度高、抗干扰能力强、稳定性好等优点。利用平面光极技术可以观测解钾菌-含钾矿物反应界面pH 分布及动态变化,从而获取有机酸等物质的分泌时间点。
4 展 望
结合光学显微术、电子显微术和荧光定量技术等各个前沿技术手段,可以跨尺度、多维度研究“功能性菌株对养分循环的影响机制”。例如:数字全息显微镜实时记录细菌在界面三维运动行为变化;原子力显微镜微尺度量化细菌和矿物界面间空间受力;激光扫描共聚焦显微镜定量生物膜三维形貌及其随时间变化情况;PO 实时微尺度表征环境物化指标变化情况。科学工作者们可结合自身目标,将上述技术巧妙结合。本文主要从土壤钾素出发综述了“解钾菌及其释钾微观机制的研究进展”,为同行们提出了新的研究思路和视角。事实上,对于常见功能性菌株而言(如:解钾菌、解磷菌、固氮菌等系列根际促生菌),其影响养分循环既存在共性又有细微差别,科学工作者们可结合本文提出的解钾菌微观机制适当的应用到其他课题中去,共同、逐步推进“功能性菌株对养分循环的影响机制”研究。
基于目前关于解钾菌的知识,我们提出以下几点未来的研究方向:
1)本文在探究解钾菌微观解钾机制过程中所应用的细菌与固-液界面微观相互作用知识同样适用于其他土壤植物促生根菌(可能会有细微差异),研究者们可结合本文思路进一步探究根际促生菌作用机理;
2)目前本文仅将细菌与固-液界面的相关研究以理论形式应用于解释解钾菌微观解钾机理之中,有关其具体的实际应用还需要进一步探索;
3)在研究解钾菌微观释钾过程中,探究水动力学相互作用、DLVO 理论、布朗运动以及细菌运动性在解钾的各个阶段的作用力大小,进一步预测解钾菌解钾能力强弱。
感谢朱堃老师对文章写作的建议,感谢博士生韩天富对文献框架和撰写思路方面的指导,感谢硕士生冉洪芋、肖鹏和雍路路对语言表述方面提供的帮助。
猜你喜欢
宝藏(2022年8期)2022-09-27
浙江农业学报(2022年7期)2022-07-28
航天器环境工程(2022年2期)2022-05-09
建材发展导向(2022年5期)2022-04-18
江苏科技报·E教中国(2021年22期)2021-12-23
中国药学药品知识仓库(2021年11期)2021-11-25
当代陕西(2020年13期)2020-08-24
科学之谜(2019年2期)2019-03-25
大陆桥视野·下(2017年12期)2017-11-29
声屏世界(2015年2期)2015-03-11
