
生物炭缓解乙草胺对草莓根系线粒体膜功能伤害的作用
2022-06-10 09:35鲁晓峰杜国栋孙海龙徐树广
中国南方果树 2022年3期
鲁晓峰,杜国栋,邵 静,孙海龙,徐树广
(1 中国农业科学院果树研究所/农业农村部园艺作物种质资源利用重点实验室,辽宁兴城,125100;2 沈阳农业大学园艺学院/沈阳市北方果树栽培与生理生态重点实验室,沈阳,110866;3 吉林省农业科学院果树研究所,吉林公主岭,136100)
乙草胺是一种作用于杂草幼芽和幼根的除草剂,具有选择性专一、持效期长及控制杂草效率高等诸多优点,在农业生产中被广泛应用[1]。然而,若在草莓繁育过程中不当使用乙草胺,则对幼苗植株的生长发育产生一定的伤害,对草莓苗木质量以及果实产量和品质会造成严重的负面影响。因此,探究乙草胺药害影响植株发育的内在生理代谢规律,对合理使用乙草胺及有效避免药害具有重要的指导意义。
除草剂可通过进入作物根围土壤来影响植物根系生理功能,从而表征除草剂胁迫对根系发育的伤害程度[2]。Ghazi-Khansari等[3]研究发现,利用草甘膦处理后,毛桃根系呈现出颜色变深、须根变短、变少等现象,说明除草剂胁迫对毛桃根系生长发育有较大伤害。土壤中的西草净含量达到0.8~8.0 mg/kg时,小麦根系中活性氧(ROS)大量产生,对根系细胞膜脂造成损伤[4]。外源施加丙酯草醚和丁草胺,均能不同程度破坏小麦、油菜和大麦体内的保护性酶活性,使植株体内的膜质过氧化产物丙二醛(MDA)含量提高[5-6]。线粒体是根系细胞主要的细胞器之一,是感受外界胁迫相关信号的主要部位[7-8]。研究表明,逆境胁迫会导致线粒体的结构和功能发生明显变化[9]。如:在盐碱胁迫下,番茄根系线粒体内H2O2和MDA含量增加,线粒体膜通透性增大[10];在干旱胁迫下,银沙槐幼苗叶片中线粒体膜完整性低,出现内部嵴消失、空泡化等现象[11];在铝胁迫下,导致花生根尖线粒体电子传递受阻,氧自由基含量增加,造成线粒体膜质过氧化,根尖细胞发生程序性死亡[12]。生物炭(粉状颗粒化的木炭)具有孔隙结构发达、吸附性能好、离子交换能力较强等特点[13-14],可有效吸附极性及非极性化合物[15],施入土壤后可影响除草剂在土壤环境中的迁移和转化[16]。Wei等[17]研究表明,生物炭对莠去津和西玛津有较高的吸附亲和力,可有效去除残留,减轻污染。李玉梅等[18]研究表明,施入生物炭后,异噁草松对甜菜生长的抑制作用得以缓解,根系吸收养分和水分的能力增强,表现为根长增加,体积增大,根尖数增多现象。虽然有关除草剂影响作物生长发育的研究报道较多,但关于除草剂导致作物根系线粒体膜结构和功能损伤以及生物炭减缓其伤害的机制等尚无详细研究报道。本文以草莓植株为试验材料,研究乙草胺诱发ROS对草莓根系线粒体膜电位、膜通透性等生物膜结构和功能的影响,明确添加生物炭缓解乙草胺伤害的生理机制,为生产上有效应对乙草胺类除草剂胁迫提供借鉴。
1 材料与方法
1.1 试验材料
供试材料为日本草莓品种“宝交早生”(Fragaria×ananassaDuch.‘Hokowase’)。浓度90%乙草胺乳油(购自吉林美联化学品有限公司),生物炭(粒径为0.10~0.15 mm)。
1.2 试验处理
将三叶一心的草莓假植幼苗定植于内径宽和高均为16.0 cm的塑料钵中。栽培基质为园土∶泥炭∶有机肥=3∶1∶1(体积比),每盆装入基质1.25 kg,每盆栽植草莓苗1株。草莓植株在塑料钵中定植30 d后,选择长势基本一致的草莓植株进行试验。试验设乙草胺处理(为突出乙草胺的毒害作用,本试验中加大了乙草胺的使用浓度,即每钵浇灌0.2 mol/L的乙草胺100 mL,编号为D)、生物炭+乙草胺处理(在栽植草莓植株前,每钵基质添加125.00 g生物炭,与园土、泥炭、有机肥混合均匀;浇灌0.2 mol/L乙草胺100 mL,编号为DB)及清水对照(未施加乙草胺和生物炭,编号为CK)3个处理。每处理每重复1株(盆),重复30次。按试验设定剂量将除草剂溶液浇灌塑料钵基质中,对照施入等量的蒸馏水。分别于处理后1 d、3 d、5 d、7 d和9 d,取草莓根系前端1~2 cm分生区部位根段样品,用于各生理指标的测定。
1.3 测定方法
1.3.1 根系分生区线粒体的提取及其特性的测定 线粒体的提取参照杨玖英[19]的方法;线粒体膜电位(Δψm)测定参照Braidot[20]等方法;线粒体膜通透性的测定参照金超芳[21]等方法;线粒体细胞色素c/a(Cyt c/a)测定参照Tonshin及马怀宇[22]等方法。
1.3.2 根系分生区膜脂过氧产物及保护性酶活性的测定 丙二醛(MDA)含量采用硫代巴比妥酸(TBA)法测定[23];过氧化氢酶(CAT)活性采用KMnO4滴定法测定[24];超氧化物歧化酶(SOD)活性采用氮蓝四唑(NBT)显色法测定[25];过氧化物酶(POD)活性采用愈创木酚法测定[26];超氧阴离子(O2·-)含量采用羟胺法测定[27];过氧化氢(H2O2)含量采用四氯化钛法测定[28]。
1.3.3 根系活力及质膜H+-ATPase活性测定 根系活力采用TTC法测定[29],单位为mg/(g·h);质膜H+-ATPase测定参照Blumwald等[30]方法,以单位质量的质膜在单位时间水解ATP所释放的无机磷(Pi)量来表示活性,单位为μmol/(mg·h)。
1.4 数据统计与分析
采用Excel 2010 软件进行原始数据处理,Sigma Plot 10.0软件绘图,SPSS软件进行差异显著性分析。
2 结果与分析
2.1 根系线粒体膜功能
线粒体膜电位(Δψm)是评线粒体功能的特征指标。施加处理D后,根系线粒体膜Δψm于处理后7 d内呈持续下降的趋势,比CK低64.3%,后期无明显变化。施加处理DB后,根系线粒体膜Δψm的下降幅度显著减小。说明,乙草胺对草莓根系线粒体膜功能产生伤害,而土施生物炭能在一定程度上缓解除草剂乙草胺对草莓植株的伤害(见图1A)。
线粒体细胞色素Cyt c是呼吸电子传递链的重要组成部分,遇到胁迫伤害易从线粒体内膜脱落,而细胞色素Cyt a则不易脱落,可用Cyt c/a来反应Cyt c量的变化。与CK相比,前期处理D和DB根系线粒体内膜上细胞色素Cyt c/a含量明显下降,均于处理7 d后达最低值,后期Cyt c/a变化不明显,处理DB的线粒体Cyt c/a降幅显著小于处理D(见图1B)。表明,乙草胺导致草莓根系线粒体内膜上的Cyt c不断流失,对线粒体的呼吸电子传递功能产生了严重的影响,而土施生物炭能够有效减缓乙草胺对根系线粒体膜功能的伤害。
渗透性转换孔(MPTP)是连接线粒体内外膜的通道,其开放程度可反映线粒体膜的通透性。与CK相比,在处理1~9 d,处理D和DB的根系线粒体膜透性(MPT)呈持续,处理DB的升幅显著小于处理D(见图1C)。可见,乙草胺对线粒体膜通透性产生了抑制作用,而土施生物炭能有效减弱乙草胺对线粒体膜通透性的负面影响。
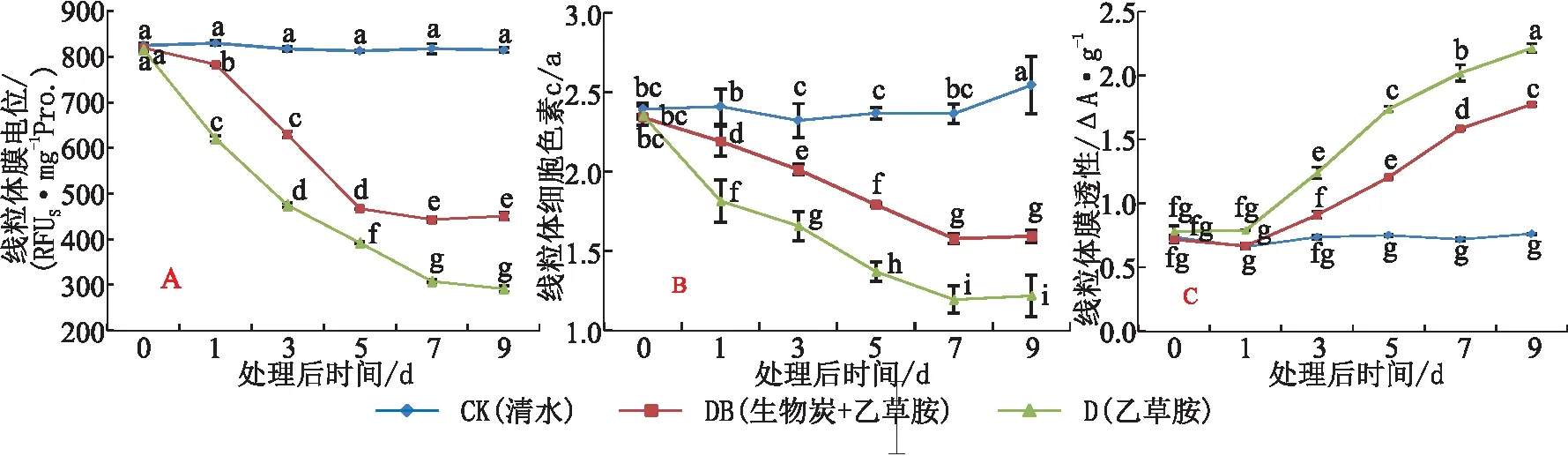
图1 乙草胺及生物炭对草莓根系线粒体功能的影响
2.2 根系活力及H+-ATPase
与CK相比,乙草胺(处理D)使草莓根系活力持续下降,处理9 d后,根系活力仅为对照的54.8%,土施生物炭(处理DB)削弱了乙草胺引起的草莓根系活力下降(见图2A)。与CK相比,处理D和DB均使草莓根系细胞质膜H+-ATPase活性呈现明显下降趋势,其中,处理D下降相对明显(见图2B)。
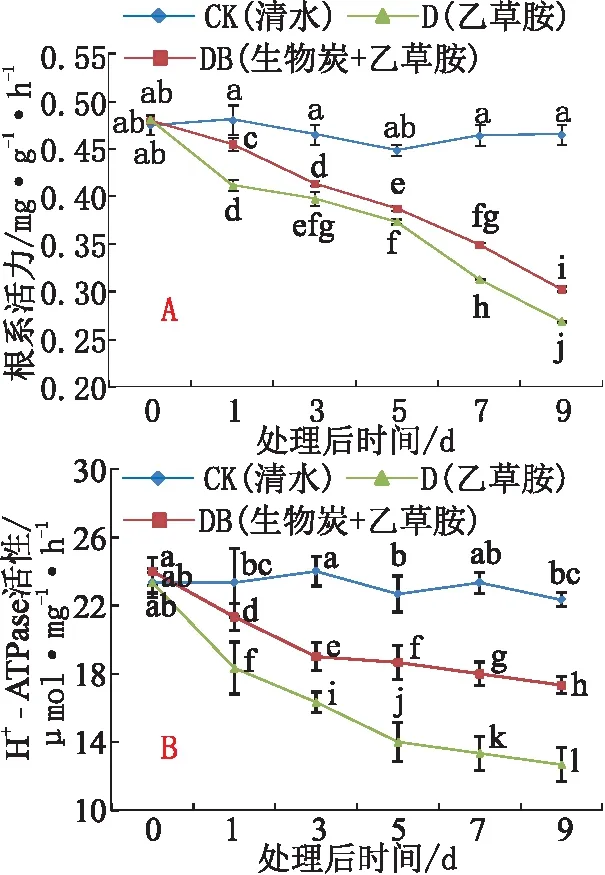
图2 乙草胺及生物炭对草莓根系活力及H+-ATPase的影响
2.3 根系丙二醛含量
丙二醛(MDA)是植物受到逆境胁迫所生成膜质过氧化物质,其含量常常用来表征植物机体内膜质过氧化程度。与CK相比,在乙草胺胁迫下(处理D和DB)的根系MDA含量均呈现持续下降趋势。其中,处理DB的MDA含量在后期(5 d及以后)下降缓慢,高于处理D(见图3)。表明,乙草胺胁迫明显造成草莓根系膜质伤害,且随处理时间延长,根系膜质过氧化程度趋于严重,而土施生物炭可有效减缓乙草胺胁迫造成的伤害。
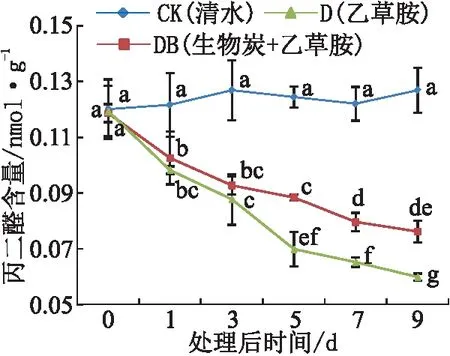
图3 乙草胺及生物炭处理对草莓根系丙二醛含量的影响
2.4 根系活性氧含量
与CK相比,随着胁迫时间延长,处理D的草莓根系超氧阴离子自由基(O2·-)含量明显上升,处理第5 d后,O2-含量较CK增长14.9%,处理9 d后O2-含量达到最高值;5 d后,处理DB的草莓根系O2-含量比CK增长9.1%,处理后期O2-含量变化不明显。
处理3 d及以后,处理DB的O2·-含量始终低于处理D(见图4A)。与CK相比,随着胁迫时间的持续,处理D的根系过氧化氢(H2O2)含量明显上升,处理7 d时H2O2含量达到最高值,较CK增长32.5%;处理DB的根系H2O2含量变化与处理D相似,但始终低于处理D(见图4B)。说明,乙草胺胁迫对草莓植株根系的伤害严重,对ROS产生有较大的影响,而土施生物炭能有效缓解活性氧对草莓根系的伤害。
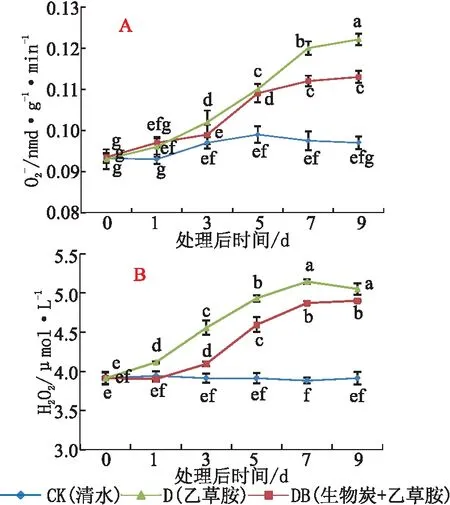
图4 乙草胺及生物炭对草莓根系ROS含量及H2O2含量的影响
2.5 根系保护性酶活性
超氧化物歧化酶(SOD)是参与超氧阴离子自由基(O2·-)分解的唯一酶类。与CK相比,随着乙草胺胁迫(处理D和DB)时间增加,草莓根系SOD活性均呈逐渐上升趋势,且处理D和DB的SOD活性差异不显著。在胁迫处理9 d后,处理D和DB的根系SOD活性分别比CK增长32.8%和27.7%(图5A)。
过氧化物酶(POD)作为胞内酶又可作为胞外酶来参与H2O2的分解,是众多涉及清除ROS类物质的最重要酶类之一。与CK相比,随着乙草胺胁迫(处理D和DB)时间的增加,根系POD活性呈现逐渐上升趋势,于处理9 d后达最高水平,后期(处理7 d及以后)处理DB的POD活性显著低于处理D(见图5B)。说明,乙草胺胁迫造成草莓根系伤害,引起根系保护性酶POD活性增加,而土施生物炭则显著缓解乙草胺对根系的伤害。
过氧化氢酶(CAT)清除剂也是生物防御体系的关键酶之一。与CK相比,随着胁迫时间增加,处理D的根系CAT活性呈波动下降趋势,处理9 d后的CAT活性达最低值,较CK降低了17.1%;处理DB的草莓根系CAT活性呈现持续缓慢下降趋势,处理9 d后的CAT活性仅比CK降低6.1%(见图5C)。表明,土施生物炭明显缓解了乙草胺的伤害作用。
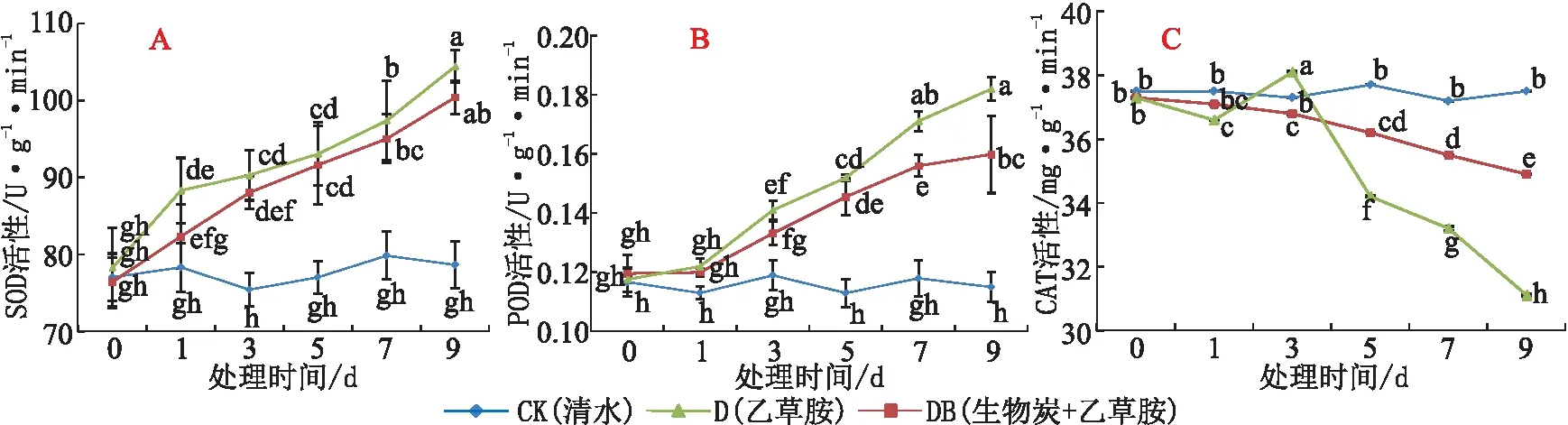
图5 乙草胺及生物炭对草莓根系保护性酶活性的影响
3 讨论
在正常生理状态下,植物组织中的ROS产生与清除处于平衡状态。当植物受到逆境胁迫时,植株体内会产生过量ROS,引起细胞膜系统受到损伤[31],引起膜脂过氧化物产物(MDA)激增[32-33]。温银元等[27]研究发现,受到扑草净胁迫后,西府海棠幼苗根系中抗氧化酶SOD、POD等含量均显著升高,O2·-和H2O2含量显著高于对照。在本研究中,乙草胺胁迫引起草莓根尖部位的SOD和POD活性提高,与前人研究结果相似;而添加生物炭后,利用其较强的吸附能力,明显减缓了乙草胺对草莓根系的直接伤害,保护了细胞膜结构[34]。刘炜[35]等研究指出,低温逆境下,小麦根系细胞质膜Ca2+-ATPase会迅速失活。本研究也发现,草莓受到乙草胺胁迫后,质膜H+--ATPase活性呈现下降的趋势,可能是根系应对胁迫的一种适应性反应;随着胁迫时间的延长,H2O2和O2·-大量积聚,膜脂过氧化加剧,导致质膜H+-ATPase活性进一步下降,质膜功能损伤。
线粒体是细胞能量代谢的场所,细胞氧化功能的中心,也是产生ROS的场所,其内含有抗氧化酶对机体有一定保护作用[36]。过量的ROS会引起线粒体膜氧化损伤,导致线粒体的电子传递受阻,内膜的完整性和呼吸电子传递链受到破坏,进一步破坏线粒体的抗氧化防御体系,使线粒体O2·-产生速率加快、H2O2含量增高、膜脂过氧化加剧[37]。在本研究中,施加乙草胺后,草莓根系线粒体的O2·-产生速率显著增加,根系线粒体膜电位(Δψm)持续降低。在植物根系中,线粒体呼吸链电子漏是O2·-产生的主要来源[38],而线粒体膜电位的下降又能导致线粒体呼吸链电子漏的增加[39]。施用乙草胺造成草莓根系线粒体膜电位降低,这可能是导致根系O2·-产生的重要原因之一。细胞色素Cyt c介导的呼吸链电子漏旁路,可以清除由于呼吸链漏电子而产生的O2·-和H2O2。徐建兴[40]研究发现,当植物遭受逆境胁迫时,线粒体内膜的完整性和呼吸电子传递链受到破坏,最终导致线粒体O2·-产生速率加快、H2O2含量增高。本研究结果显示,乙草胺胁迫处理的草莓根系线粒体Cyt c/a值明显低于CK,而生物炭+乙草胺处理的Cyt c/a含量明显高于乙草胺单独处理,根系中的H2O2含量也低于乙草胺单独处理。表明,在乙草胺胁迫下,加入生物炭可在一定程度上保护线粒体膜及其他细胞器膜,缓解乙草胺胁迫对于草莓植株根系线粒体功能的伤害,是植物根系线粒体膜免于除草剂乙草胺伤害的有效保护机制。
4 结论
乙草胺胁迫可导致草莓根系线粒体膜结构和功能受到伤害。通过土壤添加生物炭,可有效减缓乙草胺胁迫对草莓根系的伤害,增强根系抗氧化酶系统活性,维持植物根系线粒体膜整体功能,进而提高草莓植株对乙草胺胁迫的适应能力。
猜你喜欢
中国急救医学(2022年9期)2022-11-15
中华实用诊断与治疗杂志(2022年1期)2022-08-31
伴侣(2022年5期)2022-06-16
湖北农业科学(2019年22期)2019-12-23
农家致富顾问·下半月(2019年7期)2019-09-28
中国医药导报(2019年13期)2019-06-20
今日农药(2016年6期)2016-05-14
飞碟探索(2016年4期)2016-04-07
山东农药信息(2013年9期)2013-11-06
农民致富之友(2012年9期)2012-11-27
