
氮钾配施对冬油菜角果皮光合作用及光合器官氮分配的影响
2022-06-17 03:56周杨果陆志峰丛日环李小坤鲁剑巍
植物营养与肥料学报 2022年5期
李 静,周杨果,陆志峰,丛日环,李小坤,任 涛,鲁剑巍
(华中农业大学资源与环境学院/农业农村部长江中下游耕地保育重点实验室,湖北武汉 430070)
光合作用利用光能将CO2转化为碳水化合物,贡献了90%以上的作物生物量及产量[1]。叶片作为高等植物主要光合器官,是植物生长最重要的碳获取源。然而,越来越多的研究表明,非叶器官含有发育良好的叶绿体,对碳同化有很大贡献,是植物光合作用的重要补充[2-3]。冬油菜(Brassica napusL.)是我国重要的油料作物,其生长过程中存在明显的光合器官演替过程,在开花前叶片是主要的光合器官,为油菜冠层结构形成及后期角果建成贡献了大量碳水化合物。花后油菜叶片大量凋落,角果成为此时最主要的光合碳获取源。前人研究表明,尽管角果层面积指数(PAI)略低于叶面积指数(LAI),但角果冠层拦截了约80%的入射辐射,并在角果填充阶段贡献了80%~95%的总碳增益[4-5]。此外,籽粒产量的60%~70%来源于角果皮的碳固定[6],由此可见,角果光合作用对油菜产量形成具有重要意义。
角果为源库一体,其光合作用是油菜产量形成的基础。氮是光合系统构建的关键因子,直接或间接地影响光合作用。前人在水稻叶片上的研究表明,叶片氮含量增加后叶绿体变长变厚,细胞壁变薄,叶肉细胞和叶绿体面向细胞间隙的比例增加,提高了气孔导度和叶肉导度,从而改善CO2在叶片中的传导[7],但过高的叶片氮含量并未进一步提高光合作用,致使光合氮利用效率(PNUE)明显降低,这主要与施氮后叶绿体增大导致叶肉导度和Rubisco酶含量间的比值降低密切相关[8]。PNUE是氮素生理利用效率的特征之一,受光合系统各组分氮分配比例的调控,提高PNUE有助于提升作物氮利用效率[9],而PNUE的提高需要优化叶片中不同形态氮素的分配比例[10]。与氮不同,钾并非光合作用关键酶或蛋白的组成元素,钾在植物体内主要通过维持细胞渗透压及大分子物质的电荷平衡,进而影响酶的活性、气孔运动、CO2传导以及光合产物转运等影响光合作用[11]。胡文诗等[12]研究表明,钾肥施用提高了角果皮光合面积,降低了中果皮细胞壁厚度、增加了细胞及叶绿体面向细胞空隙面积,从而降低了CO2的传输阻力,提高了角果光合速率。
氮和钾在CO2传导及同化中既有相似性又有差异性和互补性,在CO2传导上,K+和NO3-均是维持细胞渗透压,调节气孔开闭的重要物质[13-14]。氮、钾均可调控叶绿体形状和分布,增加叶肉细胞和叶绿体面向细胞间隙的面积比例,降低CO2的液相传输阻力,提高叶肉导度[7,15]。在CO2同化上,氮直接参与CO2固定,高氮供应增加叶片Rubisco酶含量,施钾可以提高Rubisco酶活性[16]。此外,Hou等[17]在水稻中的研究表明,钾肥施用降低单位面积叶片氮含量,但增加了光合系统氮分配比例,进而提高叶片净光合速率和PNUE。那么在油菜角果这样一个“源、库”结合紧密的非叶光合器官中,钾如何协调光合氮分配进而改善光合作用、提高角果PNUE仍需进一步探究。为此,开展了不同氮钾肥配施对冬油菜角果形态、光合特性及光合氮分配影响的田间试验,研究了不同氮、钾营养条件下角果皮光合特性和光合系统氮分配的变化,并分析了PNUE与角果皮养分含量及光合系统不同组分分配比例间的关系,为深入理解氮钾配施提高角果光合效率的机制提供理论依据。
1 材料与方法
1.1 试验点概况
试验在湖北省武穴市梅川镇(30°06'N,115°36'E)进行,供试土壤为花岗片麻岩母质发育的水稻土,0—20 cm耕层土壤pH 5.76,含有机质32.1 g/kg、全氮1.75 g/kg、速效钾54.5 mg/kg、速效磷13.4 mg/kg、有效硼0.48 mg/kg。试验点前茬作物为水稻。
1.2 试验设计
采用氮钾两因素试验设计,氮肥(N)用量为0、90、180、270 kg/hm24 个水平,记做 N0、N90、N180、N270;钾肥(K2O)用量设0和120 kg/hm22个梯度,用K0及K120表示。试验共计8个处理分别为:N0K0、N0K120、N90K0、N90K120、N180K0、N180K120、N270K0、N270K120。每个处理3次重复,完全随机区组排列,小区面积为20 m2。
各处理磷、硼肥用量相同,分别为P2O590 kg/hm2、硼砂9 kg/hm2。供试肥料分别为尿素(含N 46%)、过磷酸钙(含P2O512%)、氯化钾(含K2O 60%)和硼砂(含B 11%)。氮肥按基肥∶越冬肥∶薹肥 = 6∶2∶2比例分3次施用,其他肥料均一次性基施。
供试油菜品种为华油杂9号,采用育苗移栽的方式种植。在2018年9月中下旬播种育苗,选取4~5片叶(苗龄约35天)大小均一的油菜幼苗移栽到试验田,移栽密度为11.25×104株/hm2。所有试验小区布置完成后在整个试验外围设保护区,保护区内亦种植移栽油菜,在试验过程中,所有田间管理,包括除草剂施用和病虫害防治等,均采用当地栽培管理方法。
1.3 测定项目与方法
1.3.1 角果光合指标的测定与计算 选取油菜盛花期同一天开花标记的主序角果(开花后35天左右,单株标记10~15个角果),此时角果位于主序中上部,长和宽均已达最大值(图1),每个小区选择3株油菜,每株油菜选取3个标记角果用于光合参数的测定。

图1 不同氮钾肥处理的角果对比照片Fig. 1 Comparative photos of silique under different N and K fertilizers
使用LI-COR6400XT便携式光合测定系统测定饱和光强下气体交换参数和CO2响应曲线。于饱和光强1200 μmol/(m2·s)下测定各处理油菜角果净光合速率(An)、气孔导度(gs)和胞间CO2浓度(Ci)。叶室内CO2浓度设定为400 μmol/mol,角果温度和叶室内空气湿度分别控制在25℃和55%~65%,流速为500 μmol/s。待角果光活化10~20 min稳定后,记录各气体交换参数。保持上述参数不变,外接CO2注入系统测定CO2响应曲线,设置CO2浓度梯度为400、300、200、150、100、80、50、400、600、800、1000、1200、1500 和 1800 μmol/mol,待参数稳定后记录各CO2浓度下气体交换参数,每个处理3次重复。参考Long等[18]的方法计算最大羧化速率(Vcmax)和最大电子传递速率(Jmax);羧化效率(CE)为Ci小于200 μmol/mol时An-Ci响应曲线拟合线的斜率;参考Ethier等[19]的方法,采用非直角双曲线模型对CO2响应曲线进行拟合,计算叶肉导度(gm)。
1.3.2 角果皮特征参数的测定 光合参数测定结束后,每个处理选取至少15个标记角果带回实验室,用游标卡尺量取角果长、宽后,将角果分为果壳和籽粒,角果皮剥开重物压平置于黑色棉布上,黑色棉布上放置25 cm2白色纸板作对照,用数码相机拍照获取图片,通过image-pro plus 6.0软件计算果皮面积。后置于105℃杀青30 min,60℃恒温烘干、称重,磨细过0.5 mm筛。
养分含量测定:采用H2SO4-H2O2消化,流动分析仪(AA3, Seal Analytical Inc., Southampton, UK)测定角果皮中氮含量,火焰光度计测定钾含量。
叶绿素的测定:剪取0.1 g果皮于比色管中,加95%乙醇定容至25 mL,黑暗浸提48 h。分别在665、649 nm处测定浸提液吸光值,计算叶绿素a、叶绿素b浓度及总叶绿素含量[20]。
1.3.3 角果皮光合系统氮分配估算 根据前人研究结果[21],将参与光合作用的氮素(Npsn)分为3个主要部分:卡尔文循环中用于羧化反应的蛋白质,归为羧化系统(Ncb);用于电子传输的蛋白质,划为电子传递组分(Net);用于光系统I、II和其他捕光色素蛋白复合物中的光捕获蛋白构成捕光系统(Nlc)。模型假设Rubisco活化数量与最大羧化速率(Vcmax)成正比。通过Rubisco的比活性(Vcr),即每单位Rubisco的最大羧化速率估算氮在羧化系统的分配比例(PNcb):
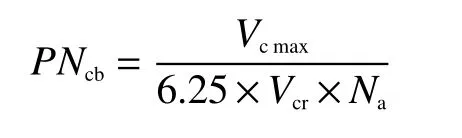
式中,6.25是Rubisco酶氮转化系数;Vcr在25℃为20.78 [CO2μmol/(g·s), Rubisco][22];Na为单位面积角果皮氮含量(g/m2)。
模型假设氮在电子传递组分的投入与最大电子传递速率(Jmax)成正比。通过单位细胞色素f在单位时间的最大电子传输速率(Jmc)估算氮在电子传递组分的分配比例(PNet):
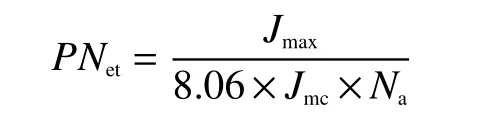
式中,8.06是每单位细胞色素f氮转换系数[23];Jmc在 25℃ 为 155.65 [e-μmol/(μmol·s), Cytf][22]。
氮在捕光系统的分配比例(PNlc)为:
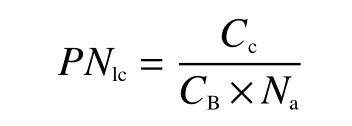
式中,Cc为叶绿素含量(mmol/m2);CB为结合光系统I (PSI)、光系统II (PSII)、捕光复合物 (LHCII)中的叶绿素,值为2.15 (mmol/g, N)[21]。
光合系统氮分配比例(PNpsn)为PNcb、PNet和PNlc三者之和;非光合组分氮分配比例(PNnon-psn)为1-PNpsn。
1.4 数据统计分析
试验数据利用Excel软件进行计算处理,采用SPSS 18.0软件中的双因素方差分析程序进行数据的统计分析,根据分析结果中F值大小,评价各因素及其交互作用的影响。采用Origin 9.0作图。
2 结果与分析
2.1 不同氮钾肥用量对冬油菜角果形态的影响
氮钾肥施用均能增加单株角果数,且存在显著的交互作用(表1)。与不施氮处理相比,施用氮肥后单株角果数平均增加了161.6个,增幅为143.9%;与不施钾处理相比,施钾后单株角果数平均增加72.4%;氮钾配施下,单株角果数较不施氮钾处理提高了1.7~3.0倍;氮肥施用对角粒数无显著影响,但施钾后角粒数显著增加,增幅为11.3%,氮钾肥施用对角粒数变化的影响具有显著交互作用,氮钾配施后角粒数平均增加了6.9%;氮肥施用对角果长和角果面积的提升作用不如钾肥(表1),与不施氮肥处理相比,氮肥施用后角果长和角果面积平均提高了2.3%和10.8%,而钾肥施用后角果长和角果面积平均增加了26.9%和29.8%;相反,钾肥施用对角果宽度的提升作用不如氮肥,钾肥施用后角果宽度平均只增加了1.7%,而氮肥施用后角果宽度平均增幅可达25.2%;氮钾肥配施有利于角果长、宽和角果面积的综合提升,氮钾肥配施后角果长和角果面积较不施氮钾肥处理分别增加了12.1%~30.2%和9.9%~43.8%,而角果宽平均只增加了6.5%;氮肥施用对角果皮比叶重(LMA)影响不显著,相反,不同氮肥用量下钾肥施用均能降低角果皮LMA,与不施钾处理相比,钾肥施用后LMA平均降低6.2%。
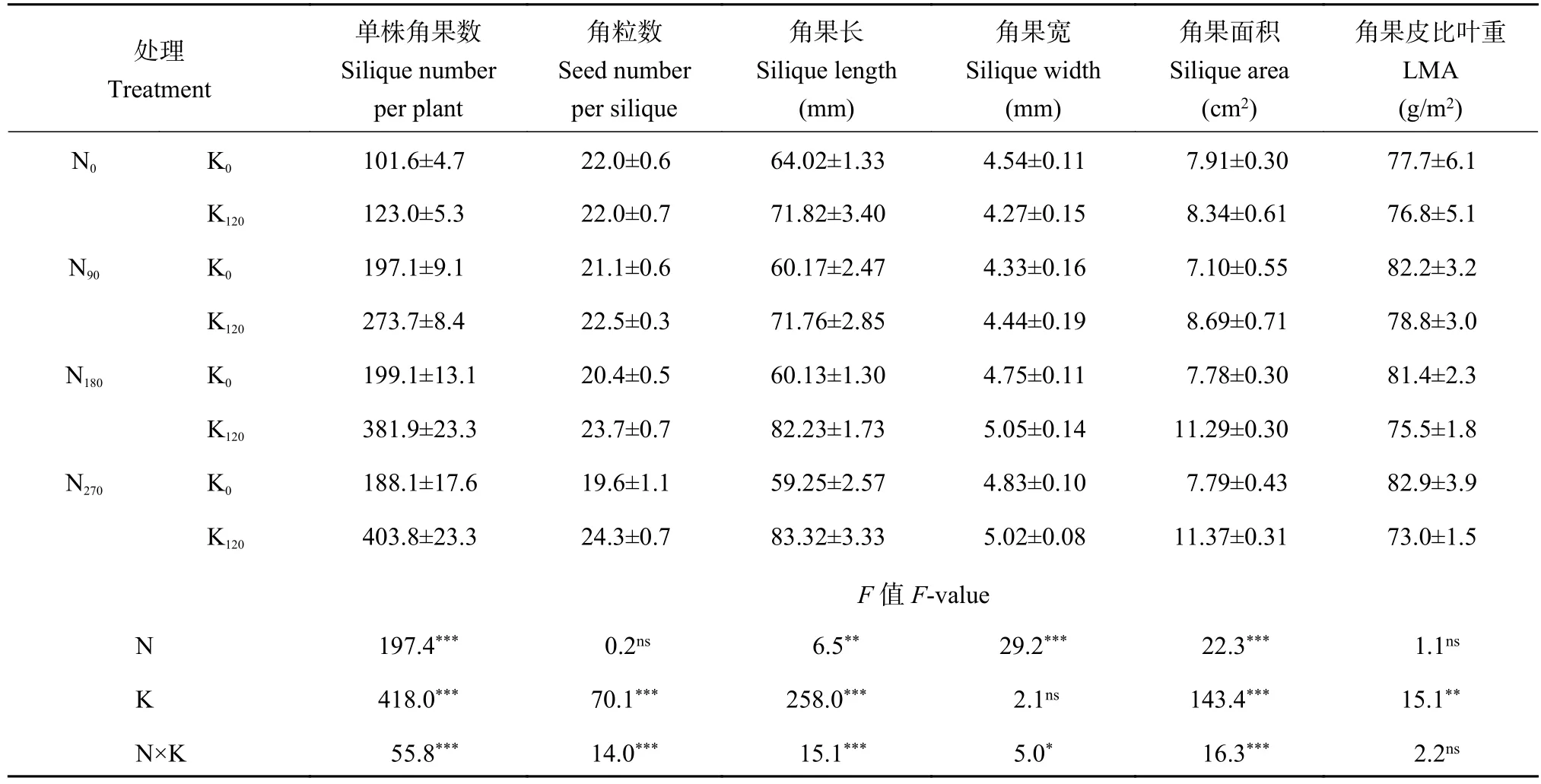
表1 不同氮钾配施处理油菜的角果形态指标Table 1 Silique morphological parameters in oilseed rape as affected by N and K application
2.2 不同氮钾肥用量对冬油菜角果皮氮钾养分及叶绿素含量的影响
钾肥施用可以降低角果皮氮含量(表2),与不施钾处理相比,施钾后角果皮氮含量平均降低了19.5%;同样地,氮肥施用也能降低角果皮钾含量,与不施氮处理相比,施氮后角果皮钾含量平均降低了20.9%;氮钾肥施用对角果皮氮钾比有显著影响,氮肥施用后角果皮氮钾比平均增加了186.1%,而钾肥施用可显著降低角果皮氮钾比,平均降幅为50.9%;角果皮叶绿素含量受氮肥供应量的显著影响,而钾肥施用对角果皮叶绿素含量的影响不大,与不施氮处理相比,施氮后叶绿素a、b含量平均增加了96.6%和75.2%;氮钾肥施用对角果皮叶绿素a/b的值有明显影响,且存在显著的交互作用,与不施氮处理相比,氮肥施用后叶绿素a/b的值平均增加了12.0%,同样地,钾肥施用后叶绿素a/b的值平均提高了9.4%。

表2 不同氮钾配施油菜角果皮氮钾养分及叶绿素含量Table 2 N and K nutrients and chlorophyll content of silique wall as affected by N and K application
2.3 不同氮钾肥用量对冬油菜角果皮光合特性的影响
如表3所示,氮肥施用后角果皮An平均提高了56.6%,钾肥施用后An平均增加了42.0%;氮钾配施较不施氮钾肥处理An平均提升了115.0%;角果皮气孔导度(gs)和叶肉导度(gm)均受氮钾营养的调控且存在显著的交互作用。在不施钾时,氮肥施用后gs降低了19.6%,而供钾充足时氮肥施用对gs影响不显著,相反,钾肥施用后gs提高了30.5%。氮肥施用对gm的影响同样受钾肥供应量的影响,在不施钾时,氮肥施用后gm提高了39.8%,而在钾供应充足时施氮gm增幅可达77.4%;氮钾配施后gs和gm分别较不施氮钾处理平均高11.1%和158.8%。从CO2同化上看,随着胞间CO2浓度的升高,角果皮净光合速率逐渐增加(图2)。不同氮肥施用量下配施钾肥均能提高角果皮CO2同化特性,且随氮肥施用量的增加,钾对CO2同化特征参数的提升作用更加明显(表3)。最大羧化速率(Vcmax)、最大电子传递速率(Jmax)和羧化效率(CE)受氮钾肥施用的显著影响,与不施氮处理相比,氮肥施用后Vcmax、Jmax和CE分别提高了44.0%、45.1%和57.4%,同样地,钾肥施用后Vcmax、Jmax和CE分别较不施钾处理增加了35.9%、31.4%和43.1%;而与不施氮钾肥处理相比,氮钾配施后Vcmax、Jmax和CE增幅可达88.2%、88.7%和113.0%;进一步分析不同氮钾肥施用量下角果皮光合氮利用效率(PNUE)的差异发现,PNUE受氮钾肥施用的显著影响,氮肥施用后PNUE降低了21.1%,而钾肥施低了21.1%,而钾肥施用后PNUE提高了65.7%,且钾肥施用对PNUE的提升作用随氮肥施用量的增加而提高。
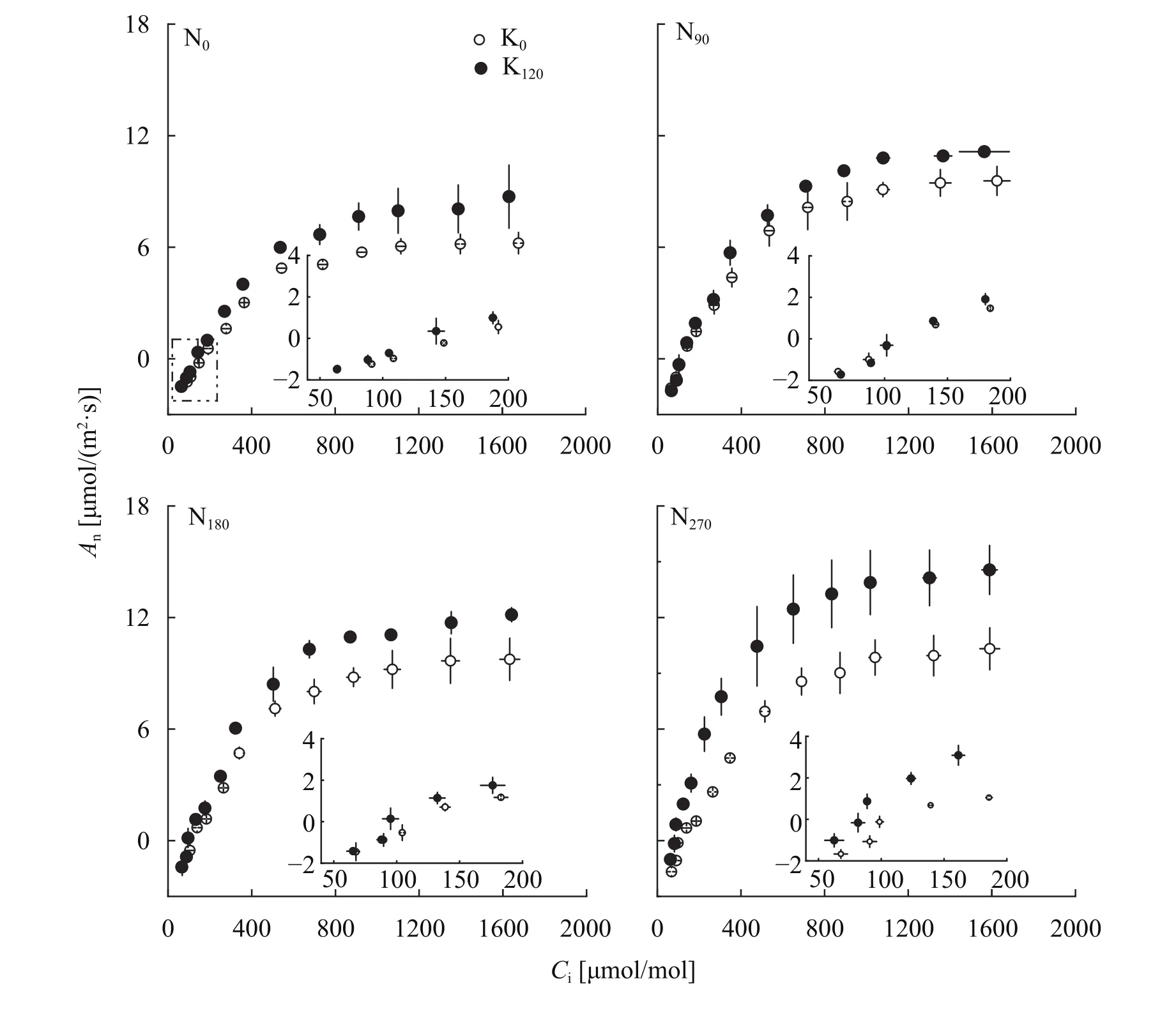
图2 不同氮钾肥用量下油菜角果CO2同化响应曲线Fig. 2 CO2 assimilation response curve of oilseed rape silique under different N and K application rates

表3 不同氮钾配施处理角果光合特征参数Table 3 Silique photosynthesis parameters as affected by N and K application
2.4 不同氮钾肥用量对冬油菜角果皮光合系统氮分配的影响
氮钾肥施用不仅影响角果皮氮含量(表2),还对角果皮中光合氮即羧化系统氮(Ncb)、电子传递系统氮(Net)、捕光系统氮(Nlc)和非光合氮(Nnon-Npsn)库容及比例产生显著影响(图3)。角果皮中氮主要以Ncb及Nnon-Npsn为主,分别占角果皮全氮的12.0%~34.7%和47.3%~79.9%,而参与光能捕获及电子传递的氮占比较小。氮钾肥施用显著影响角果皮光合氮(Npsn)及非光合氮分配,与不施氮处理相比,氮肥施用后Npsn库容平均增加了51.1%,但光合系统氮分配比例(PNpsn)却降低了11.2个百分点,其中Ncb与Net分配比例分别降低了8.4和2.5个百分点,而Nlc分配比例在氮肥施用后增加了0.34个百分点;与氮肥施用不同的是,钾肥施用后可以显著提高角果皮光合系统氮库容量和分配比例,与不施钾处理相比,钾肥施用后角果皮Npsn库容量平均提高了28.7%,PNpsn增加了15.6个百分点,进一步细分钾对光合作用各部分氮分配比例提升作用差异发现,钾肥施用后对Ncb库容增加提升作用最大,增幅可达35.9%,其次是Net,施钾后其增幅也可达31.4%,而钾肥施用对Nlc库容的提升作用相对较小,施钾后Nlc只提高了4.2%;与不施氮钾肥处理相比,尽管氮钾配施后光合系统氮分配比例平均只提高了2.9个百分点,但光合系统氮库容量却提升了90.7%,远高于单施氮肥(51.0%)或单施钾肥(28.7%)对角果皮光合氮库的提升。
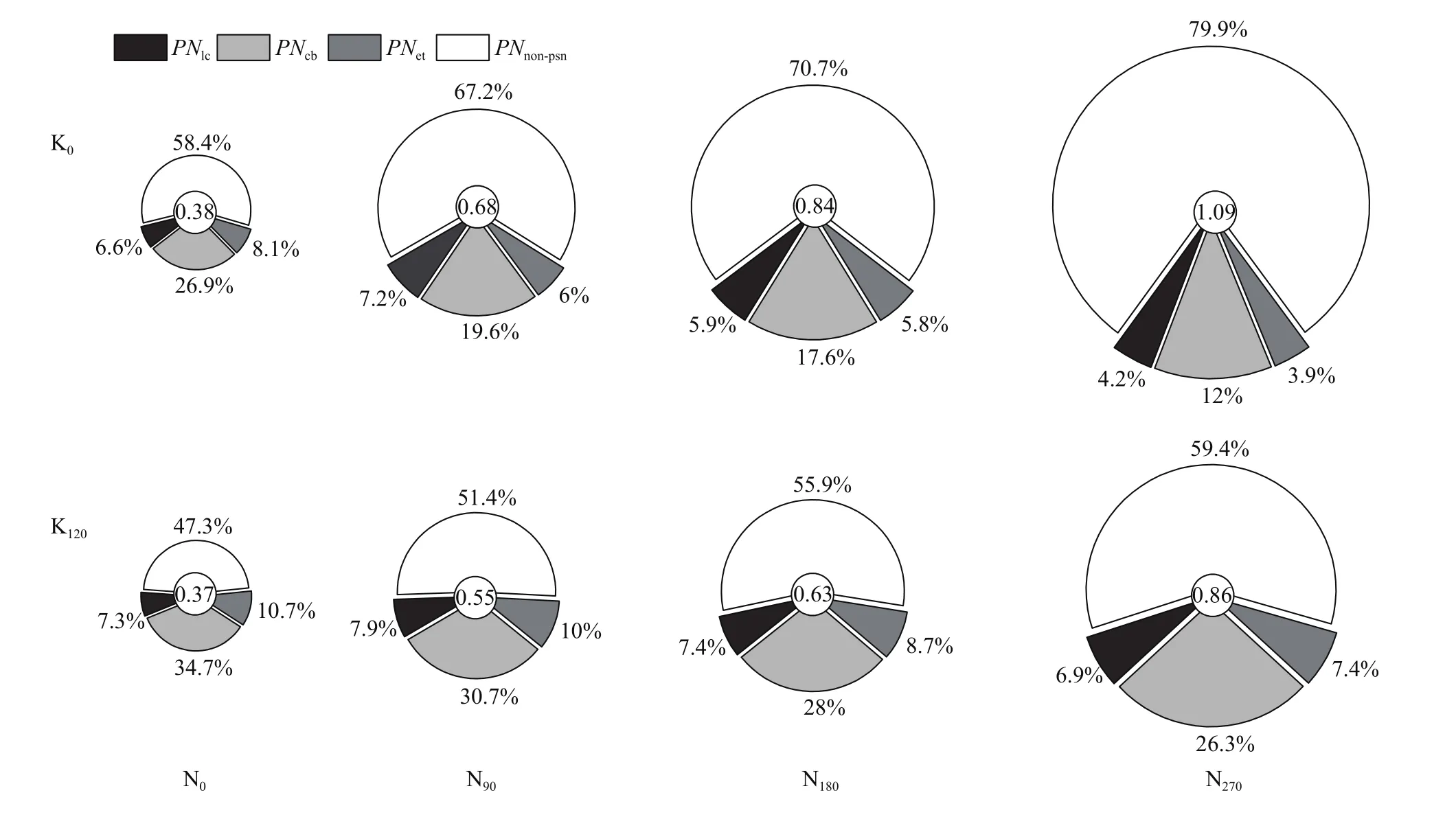
图3 不同氮钾肥供应对角果皮光合系统氮分配的影响Fig. 3 Effects of different N and K supply on N allocation within the photosynthetic apparatus of silique wall
2.5 光合氮利用效率、叶片氮分配比例与角果皮氮钾养分含量间的关系
进一步分析光合氮利用效率(PNUE)与角果皮氮钾养分含量及氮钾比关系可以看到(图4),PNUE与角果皮氮含量及氮钾比均呈极显著负相关关系,且与氮钾比间关系更为密切,而与角果皮钾含量呈极显著正相关关系。由此可见,PNUE受氮钾养分的综合调控,氮钾营养失衡PNUE急剧下降,而施钾可以极大地提高PNUE。此外,PNUE受光合系统中光合氮分配比例的调控,由分析可以看出,PNUE与光合系统中各组分比例均呈显著正相关关系,其中与羧化系统氮分配比例(PNcb)相关性最高,而与非光合系统氮分配比例(PNnon-psn)呈极显著负相关;同样地,角果皮光合系统氮分配比例受角果皮氮钾养分含量及比值的影响,捕光系统的氮分配比例(PNlc)、羧化系统氮分配比例(PNcb)和电子传递组分的氮分配比例(PNet)与角果皮氮含量呈极显著负相关,而非光合组分氮分配比例(PNnon-psn)与角果皮氮含量呈极显著正相关;相反,光合系统中不同组分氮分配比例与角果皮钾含量均呈极显著正相关,而非光合组分氮分配比例(PNnon-psn)与角果皮钾含量呈极显著负相关。与角果皮氮含量或钾含量相比,角果皮氮钾比对角果皮光合系统氮分配比例的影响要更大。由此可见,氮钾肥施用通过影响角果皮氮钾养分含量和比值从而调节角果皮光合系统氮分配比例进而调控PNUE,随着施氮量的升高角果皮分配于光合器官中的氮素比例下降,非光合组分氮分配比例上升,导致PNUE下降,而钾含量的增加提高了光合器官中的氮分配比例,降低了非光合组分氮分配比例进而提高PNUE。

图4 冬油菜角果皮光合氮利用效率、光合系统中氮分配比例与角果皮养分含量间关系Fig. 4 Relationship between photosynthetic N utilization efficiency, N distribution ratio in the photosynthetic system,and nutrient content of siliques wall in winter rape
3 讨论
调控光合面积和光合速率是提高作物生产力的主要途径。氮和钾是植物必需的大量营养元素,可协同调控植物光合面积和光合能力[24]。本研究中,氮钾配施可以提高角果皮光合面积,调节角果皮氮钾养分含量和比例,提高角果皮叶绿素含量,增加角果光能捕获,进而提高角果皮的净光合速率;此外,表征角果皮CO2同化能力的最大羧化速率(Vcmax)和羧化效率(CE)受氮钾营养的共同调控且表现出显著的交互作用。氮肥施用对角果皮CO2同化能力的影响受钾肥供应量的影响,在钾供应不足时,氮肥施用量的增加对Vcmax和CE的提升作用较小,这主要与角果皮中氮钾养分失衡加剧有关,在高N/K下,蛋白质合成受阻,低K浓度会导致游离氨基酸的积累,并破坏蛋白质上的电荷平衡,降低氮代谢和氮同化酶活性[25-26]。相反,钾供应充足可以提高氮同化酶及Rubisco酶含量和活性,提高Vcmax和CE,进而提高光合速率[27]。前人在叶片上的研究结果表明,氮钾配施可以协同提高CO2传导和同化能力,从而增强光合作用[17,24]。在角果皮中氮钾配施同样可以协调CO2传导、改善CO2同化能力,从而提高单个角果生产力。
前人研究表明,优化光合系统中的氮分配比例,可以提高作物光合能力,从而提高氮素光合利用效率[28]。Mu等[29]在玉米上的研究发现,玉米叶片倾向于投入更多的氮用于维持电子传递,相应的减少捕光系统中氮分配比例以应对低氮环境。本研究中,氮肥施用增加了角果皮光合系统氮库,但降低了角果皮光合系统中氮分配比例从而降低了角果皮光合氮利用效率(PNUE);相反,钾肥施用后显著提高了角果皮光合系统氮库及分配比例,其中对羧化系统氮分配比例的提升作用最大。Warren等[30]指出高氮叶片中的单位Rubisco酶活性较低,叶片中增加的Rubisco酶更多以存储氮的形式存在,并没有催化能力。因此,随着施氮量的增加角果皮羧化系统中的分配比例的降低,并不是角果皮中Rubisco酶含量下降,而是有催化活性部分的比例较其他处理低,从而降低了PNUE。钾肥施用提高角果皮光合系统氮分配比例主要与羧化系统中Rubisco酶含量和活性提高密切相关。综上,氮肥施用增加了光合氮库,但降低了角果皮光合氮分配比例,而钾肥施用可以调动部分存储氮活性,增加光合系统氮分配比例,从而提高角果皮PNUE。
4 结论
角果形态受氮钾营养的综合调控,氮钾配施后角果长、宽及角果面积显著增加。角果An的提高与氮钾配施协调角果皮氮钾营养平衡、改善角果皮CO2传导及同化密切相关。施氮后增加了角果皮氮库容量,但降低了光合系统中各组分氮分配比例,从而降低PNUE。相反,钾肥施用显著提高了角果皮羧化、电子传递和捕光系统氮分配比例,进而提高角果An,改善PNUE。此外,氮钾配施对群体角果数的提升作用巨大。因此,在实际生产中要氮钾肥合理配施,在增加角果皮光合氮库的同时优化光合系统氮分配比例,从而最大程度地提升个体光合潜能,进而提高群体生产力以达到增产增效的目的。
猜你喜欢
山地农业生物学报(2022年5期)2022-10-15
农业工程学报(2021年23期)2021-03-11
小学生学习指导(低年级)(2019年9期)2019-09-25
安徽科技学院学报(2018年5期)2019-01-18
小学生优秀作文(低年级)(2018年12期)2018-12-13
热带农业科学(2018年2期)2018-06-21
速读·中旬(2017年11期)2017-11-11
理论导刊(2017年8期)2017-08-30
中华手工(2016年4期)2016-04-20
现代管理科学(2009年10期)2010-02-03
