
红豆杉叶际共生菌参与紫杉醇合成的宏基因组学分析
2022-07-04 06:56董华恒温欣于淼陈营
农业与技术 2022年12期
董华恒 温欣 于淼 陈营
(烟台大学生命科学学院,山东 烟台 264005)
红豆杉是红豆杉目红豆杉科红豆杉属的一种乔木,又称紫杉,也称赤柏松,是世界上公认的濒临灭绝的天然珍稀抗癌植物。红豆杉的药用价值主要体现在其提取物——次生代谢衍生物——紫杉醇。紫杉醇是从红豆杉的树皮中分离提纯的天然次生代谢产物,在临床上已经广泛用于乳腺癌、卵巢癌等的治疗。紫杉醇作为一个具有抗癌活性的二萜生物碱类化合物,其广泛而显著的生物活性、全新独特的作用机制等使其受到了植物学家、化学家、药理学家、分子生物学家的极大青睐,使其成为20世纪下半叶举世瞩目的抗癌明星和研究重点[1,2]。但是从红豆杉树皮中分离提纯的紫杉醇数量少,远远满足不了临床的需要[3,4],也有学者采用化学合成的方法,但其合成路线复杂、反应条件难以控制,合成率低[5],微生物发酵法生产虽具有生产周期短、操作简单的优势,但是目前已知的微生物生产紫杉醇产量很低,不能够满足产业化生产要求,因此获得能够适用于发酵生产的工业菌株是实现发酵生产紫杉醇的前提。
宏基因组学是一种直接对微生物群体中包含的全部基因组信息进行研究的手段。其规避了对样品中的微生物进行分离培养,提供了一种对不可分离培养的微生物进行研究的途径,更真实地反映样本中微生物组成、互作情况,同时在分子水平对其代谢通路、基因功能进行研究[6,7]。本实验基于宏基因组技术分析红豆杉叶发酵液的微生物菌群,分析其主要代谢通路和合成紫杉醇的相关酶和微生物,以期为后续实验提供优势菌种,从而缩短提取时间,提高紫杉醇的提取率。
1 材料与方法
1.1 材料与试剂
1.1.1 原料
采摘加拿大曼地亚红豆杉针叶和枝条,置于冰箱内冷藏备用。
1.1.2 培养基
富集培养基:酵母膏0.5g,(NH4)2SO43g,NaCl 5g,KH2PO40.4g,MgSO40.3g,FeSO40.01g,蒸馏水1000mL。
1.2 实验方法
1.2.1 样品制备
取适量实验室留存的红豆杉枝叶样品清洗晾干后取其叶片绞碎,样品置于300mL锥形瓶中。配制1000mL富集培养基,121℃灭菌20min。向锥形瓶中加入10g针叶碎片,35℃、140rpm震荡培养,直至培养液浑浊并且针叶碎片出现明显降解,为原代共生菌富集液。取原代富集液按5%接种含10%红豆杉叶的富集培养基,35℃、140rpm震荡培养4~5d,为一代菌群富集液。同样操作至三代菌群富集液。每代菌群富集液为3个平行。
在超净台中分别吸取上述最终菌群富集液的3个平行样品4mL,置于灭过菌的离心管中,8000rpm离心5min。取出后倒掉上清液,留底部沉淀,命名为h1、h2、h3,冷冻保存后送于公司进行微生物宏基因组分析。
1.2.2 宏基因组测序
本实验样本采用Illumina Novaseq平台对测序样本进行双端测序。采用Illumina测序平台测序获得的原始数据(Raw Data)存在一定比例低质量数据,为了保证后续分析的结果准确可靠,需要对原始的测序数据进行预处理,包括质控和去宿主序列,获取用于后续分析的有效序列(clean data)。3组数据经过过滤后剩余的序列数占Raw reads 的百分比相近。测序质量基本合格,可以用于后续分析。
1.2.3 物种组成分析
为了研究样品物种组成及多样性信息,用Kraken2对所有样品的全部的有效序列进行注释分类。继续用Bracken对Kraken2得到的分类结果进行分类后贝叶斯重新估算丰度来估算宏基因组样本的物种水平或属级丰度。
1.2.4 功能数据库注释
使用HUMAnN2软件,将质控和去宿主之后的序列与蛋白质数据库进行比对,过滤掉比对失败的reads,统计UniRef90各个蛋白的相对丰度,根据UniRef90 的ID和各个功能数据库ID的对应关系,统计各个功能数据库对应功能相对丰度。
2 结果与讨论
2.1 红豆杉叶际共生菌群的多样性分析
本实验首先是以红豆杉叶为唯一碳源进行富集培养,使富集到的菌群均为红豆杉叶际共生菌,以排除其他杂菌的干扰,确保这些检测到的菌种与紫杉醇的合成代谢相关。
由每个样本在各分类水平注释的相对程度可知,本实验样品的所有序列都能够确定到种水平上。由图1可以看出,3个样本的绝对优势菌门为Ascomycota(子囊菌门)存在较高的相对丰度。在属水平上,该样本的优势菌属有Wickerhamomyces(威克罕默氏属)、Clavispora(棒孢酵母属)、Meyerozyma(季也蒙念珠菌)、Aspergillus(曲霉属)。在种水平上,Wickerhamomycesanomalus(威克罕默氏菌)、Clavisporalusitaniae(葡萄牙棒孢酵母)、Meyerozymaguilliermondii(季也蒙念珠菌)、Aspergillusnidulans(构巢曲霉)为样本的优势菌种,且威克罕默氏属的威克罕默氏菌为绝对优势菌种,存在较高的相对丰度。上述检测结果与Flores-Bustamante ZR等[8]的研究结果一致,即紫杉醇的生产者主要属于子囊菌门,故这些菌种可能与紫杉醇的合成有关。

图1 各个样品在门、属、种水平上的相对分布情况柱形图注:相对丰度前20的物种。
2.2 红豆杉共生菌主要代谢通路及紫杉醇合成的分析
对样本的基因进行KEGG注释,统计其可能参与或涉及的代谢途径。由图2可知,样本的功能基因在Level1层级有6大类的代谢通路,分别为细胞过程(Cellular Processes)、环境信息处理(Environmental Information Processing)、遗传信息处理(Genetic Information Processing)、人类疾病(Human Diseases)、新陈代谢(Metabolism)、生物体系统(Organismal Systems),其主要位于新陈代谢的代谢途径中。主要功能基因二级代谢途径有20类,其中富集差异表达基因最多的8个KEGG通路包括氨基酸代谢(Amino acid metabolism)、碳水化合物代谢(Carbohydrate metabolism)、辅因子和维生素的代谢(Metabolism of cofactors and vitamins)、信号转导(Signal transduction)、翻译(Translation)、折叠、分类和降解(Folding,sorting and degradation)、运输和分解代谢(Transport and catabolism)、内分泌系统(Endocrine system)。

图2 KEGG代谢通路的Level1和Level2层级丰度柱形图
其中,氨基酸代谢和碳水化合物代谢最为活跃。在该样本中检测到了一些高丰度碳代谢基因,包括pgm(磷酸葡萄糖变位酶)、己糖激酶(HK)的基因、galM(醛糖-1-差向异构酶)等,共11个。糖酵解/糖异生途径的特征基因明显多于其他碳代谢途径,这说明糖酵解和糖异生途径在该样本的碳循环过程中比较活跃,可以为生命活动提供能量[9]。除此之外,氨基酸代谢和碳水化合物代谢所产生的一些中间产物如乙酰辅酶A、丙酮酸、甘油醛-3-磷酸等与紫杉醇的合成相关。紫杉醇类化合物是通过异戊二烯前体合成的,萜类的异戊二烯结构单元可以通过位于细胞质中的甲羟戊酸途径(MVA)或位于质体中的磷酸甲基赤藓糖醇途径(MEP)合成。MVA途径起始于乙酰辅酶A,MEP途径则起始于丙酮酸和甘油醛-3-磷酸,这2种途径再经过共同的中间体IPP最终合成紫杉醇[5]。
通过查询NCBI数据库以及相关文献[8,9]找到了合成紫杉醇的一些关键酶以及能分泌这些酶的微生物[10-12],如分泌去乙酰巴卡蛋白III-10β-O-乙酰转移酶(Deacetyl Baccatin III-10β-O-acetyltransferase,DBAT)的菌种有Aspergillus candidus(念珠曲霉)、Fusarium solani(茄病镰刀菌)等;分泌香叶酰香叶酰二磷酸合成酶(Geranylgeranyl diphosphate synthase,GGPPS)的菌种有Clavisporalusitaniae(葡萄牙棒孢酵母)、Aspergillus luchuensis(芦竹曲霉)、Fusarium solani(茄病镰刀菌)等;Fusarium solani (茄病镰刀菌)也是能分泌紫杉二烯合酶(Taxadiene synthase,TS)的菌种。
上述菌种均属于本实验的优势菌门子囊菌门,其中包括排名第2的优势菌种Clavispora lusitaniae(葡萄牙棒孢酵母),以及排名第4的优势菌属Aspergillus(曲霉属)与该实验结果相对应。这些优势菌种的存在为后续实验菌种的筛选提供了理论基础。
2.3 红豆杉共生菌主要碳水化合物活性酶与紫杉醇活性的关系
碳水化合物活性酶数据库(CAZy)共包括糖苷水解酶、糖基转移酶、多糖裂合酶、碳水化合物酯酶、碳水化合物结合模块、辅助模块酶类6大类蛋白质家族。由CAZy功能注释结果可知,GT48、GH3、GT62、GH85、GH32、GH5等6类碳水化合物活性酶的活性在样本中较高,其中样本中的特征碳水化合物活性酶为GH5(糖苷水解酶家族5)。
糖苷水解酶在寡糖合成、烷基糖苷和芳香基糖苷的合成、氨基酸和多肽的糖基化以及抗生素的糖基化方面发挥了重要作用。糖基转移酶在生物体内催化活化的糖连接到不同的受体分子,如蛋白、核酸、寡糖、脂和小分子上[13]。糖基化是生物细胞中最重要的反应之一,这种天然的修饰存在于多种生物学活性不一样的天然化合物中,包括抗生素、抗癌药物、激素、生物碱以及黄酮等多种代谢产物[14]。该样品中还检测到了一些抗生素的合成代谢途径,如图3所示,新霉素和卡那霉素属于氨基糖苷类抗生素,这类抗生素的生物合成途径中,糖基化通常是修饰的最后一步,通过糖的位置、类型和数量的改变对抗生素的活性进行调节;图4中的新生霉素属于糖苷类抗生素,其通常含有一个或多个糖基,研究表明,糖基可以维持、调节和改变糖苷类抗生素的生物活性,糖基化可以增加糖苷类抗生素的极性和水溶性,也可以增强某些抗生素的化学稳定性[15,16]。糖苷水解酶的糖基化作用不仅可以改变抗生素的活性,还能改变紫杉醇的特性,在植物体内除了有游离状态的紫杉醇还有糖基化的紫杉醇,紫杉醇的糖基化可以大大提高其水溶性,从而便于其在临床水平上的应用。
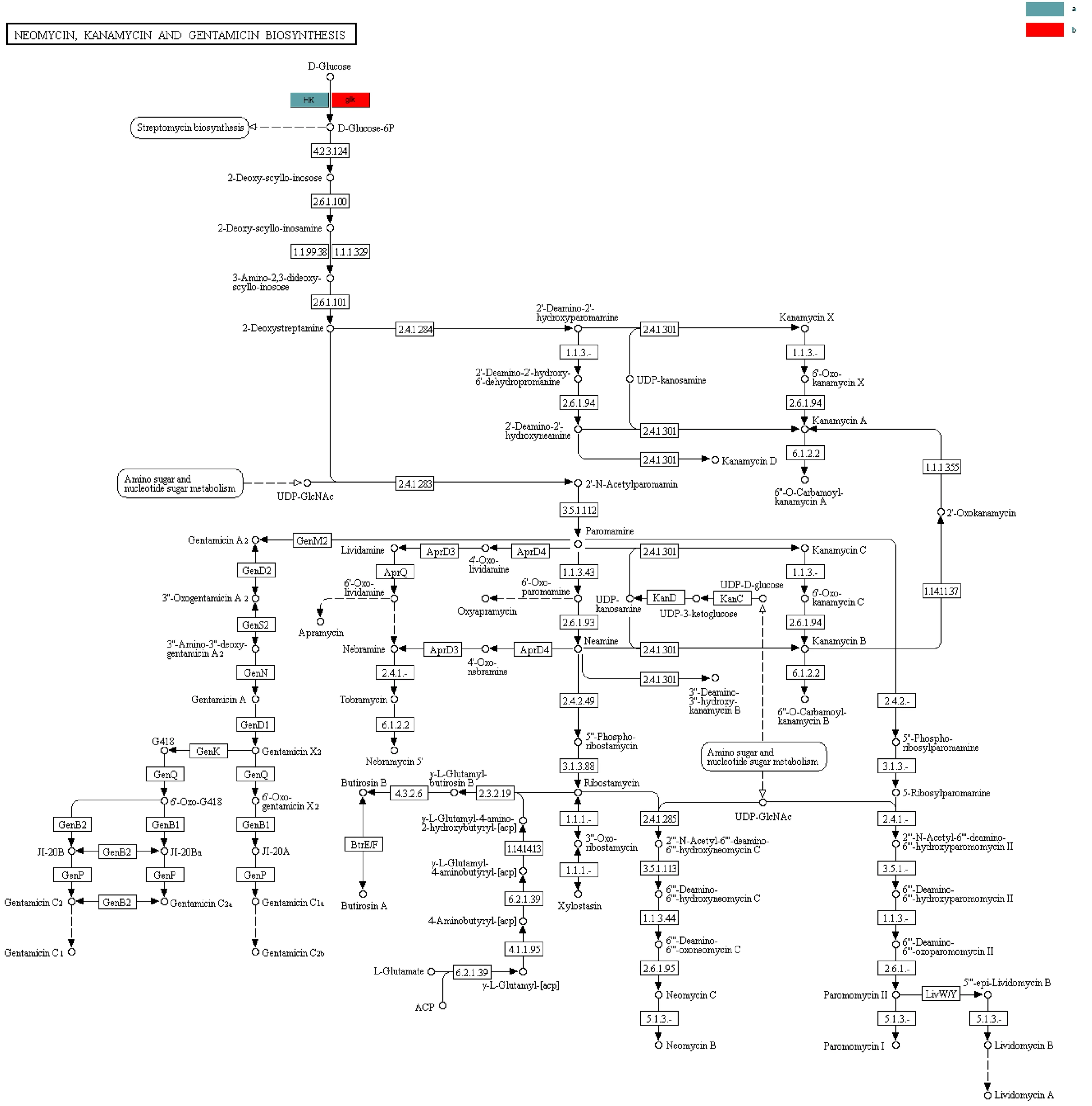
图3 合成新霉素、卡那霉素和庆大霉素的相关途径

图4 合成新生霉素的相关途径
3 结论
红豆杉是世界上公认的濒临灭绝的天然珍稀抗癌植物,其药用价值主要体现在其提取物——次生代谢衍生物紫杉醇,故获得合成紫杉醇的菌种,对缩短提取时间,提高紫杉醇的提取率具有重要意义。本实验以红豆杉叶为唯一碳源进行富集培养,以确保富集到的菌群均为红豆杉叶际共生菌,从而排除其他杂菌的干扰。后经宏基因组技术分析可得,本实验所检测到的菌种均处于紫杉醇的主要生产者——子囊菌门,且样本中也存在合成紫杉醇的菌种Clavispora lusitaniae(葡萄牙棒孢酵母),故通过分析推测样本中的菌种可以参与紫杉醇的合成代谢,并且其分泌的糖苷水解酶和糖苷转移酶可以使紫杉醇发生糖基化修饰,从而提高其水溶性,便于其在临床水平上的应用。
猜你喜欢
现代畜牧科技(2021年6期)2021-07-16
西江月(2018年5期)2018-06-08
广西林业科学(2016年4期)2016-03-16
物理化学学报(2015年7期)2015-12-30
医学研究杂志(2015年12期)2015-06-10
肝胆胰外科杂志(2015年4期)2015-02-27
应用化工(2014年3期)2014-08-16
中国粮油学报(2014年7期)2014-02-06
现代检验医学杂志(2014年1期)2014-02-06
中国洗涤用品工业(2011年3期)2011-03-20
