
蜡梅科植物次生木质部导管分子形态特征
2022-07-08 05:43李红芳张小玲
广西林业科学 2022年3期
李红芳,薛 成,张小玲
(1.咸阳职业技术学院,陕西西安 712000;2.渭南职业技术学院,陕西渭南 714000;3.中国科学院植物研究所 系统与进化植物学国家重点实验室,北京 100093;4.清华大学附属中学秦汉学校,陕西咸阳712000)
蜡梅科(Calycanthaceae)是第三纪孑遗植物,该科植物是著名的观花树种,且具有药用价值,为落叶或常绿灌木或小乔木,主要分布于温带,仅两属,7 ~9 种[1-2]。其中,蜡梅属(Chimonanthus)植物和夏蜡梅属(Calycanthus)的夏蜡梅(Calycanthus chinensis)为中国特有植物,也是我国二级重点保护植物。
蜡梅科的系统位置一直存在争议。恩格勒(Engler)系统将蜡梅科置于毛茛目(Ranales),《中国植物志》[1]采用这种分类方式;克郎奎斯特(Cronquist)系统中,蜡梅科隶属于樟目(Laurales)[3];哈钦松(Hutchison)系统将蜡梅科置于蔷薇目(Rosales)[4];在根据分子数据建立的APG 系统中,蜡梅科归属于樟目(Laurales),属基部被子植物的木兰分支[5-6],在基部被子植物中演化程度较高,目前这种分类方式得到普遍认可。
蜡梅科的系统学研究多依据早期的形态学、解剖学和孢粉学等证据[7-10]。虽然近年来有一些基因研究[2,5],但其解剖学数据依然发挥着不可忽视的作用。导管作为被子植物的特征之一,其形态是系统学研究的重要部分。对于蜡梅科木质部解剖结构的研究不多,曾有一些对Idiospermum植物的研究[11-12];对蜡梅属植物和夏蜡梅的研究较少,仅Metcalfe[12]对其进行了报道。近年来,扫描电子显微镜(SEM)开始应用于导管研究,越来越多导管分子特征被发现,如纹孔膜、原本无导管类群的植物中发现导管[13-15]等。因此,有必要对蜡梅科植物的导管分子进行进一步研究。
本研究选择两个中国特有种,即蜡梅(Chimonanthus praecox)(蜡梅属)和夏蜡梅(夏蜡梅属),利用扫描电镜对其次生木质部导管分子进行观察,并将结果进行展示,以期为蜡梅科植物的解剖学和系统学研究提供更多的资料和依据。
1 材料与方法
1.1 材料
选取蜡梅和夏蜡梅3年生以上枝。蜡梅取自陕西师范大学校园(108°95′E,34°20′N),夏蜡梅取自西安市植物园(108°96′E,34°20′N),均为人工栽培植株。凭证标本存放于陕西师范大学生命科学学院植物标本室(SANU)。
1.2 方法
离析法:将材料用FAA 固定,切成1 cm 长、火柴棍粗细的细条,用Jeffrey 离析液(10%铬酸∶10%硝酸= 1∶1),40 ℃下离析约48 h。离析后喷金镀膜,在Hitachi S-570扫描电子显微镜下观察。
石蜡切片法:材料进行石蜡包埋后,石蜡切片机切片,脱蜡,CO2临界点干燥,喷金镀膜后在Hitachi-700扫描电子显微镜下观察。
1.3 数据处理
在显微镜下测量导管分子的长度与直径。采用SPSS Version 22.0 软件进行以下分析:计算均值和变异系数;采用Student's t-test对数量性状进行差异显著性分析;采用Pearson相关性检验对数量性状进行相关性分析。
2 结果与分析
导管作为被子植物的特征之一,在解剖学和系统学上均有重要意义。普遍认为单穿孔板、直径变大和长度变短是导管演化的方向,网状或梯状穿孔板、细而长是导管相对原始的性状[16-18]。导管和穿孔板的形态特征一定程度上可显示一个类群的演化水平,也是系统学分类的重要形态学证据之一。
2.1 导管分子形态特征
2.1.1 蜡梅导管分子形态观察
蜡梅次生木质部中,导管分子端壁均为单穿孔板(图1)。穿孔板倾斜角度不同,多数穿孔板较倾斜(图1a ~g),极少数穿孔板较平截(图1h)。
穿孔板大多具尾,尾普遍较长,尾长度与形状有差异。尾长度变化较大,形状不一(图1a ~h)。有的穿孔板尾很短(图1a);有的穿孔板尾很长(图1b),单侧尾长度通常接近导管分子的主体长度,即尾长度占整个导管分子的1/2 ~2/3。除少数很短的尾(图1a)外,穿孔板尾的形状大致分为两种,一种前端较尖(图1d ~f),数量较多;另一种前端较圆(图1b,g)。也有穿孔板不具尾(图1h,j),通常较平截(图1h),数量很少。
导管分子两端的穿孔板形态可能相同(图1a),也可能不同(图1 c,g ~h)。穿孔板形态不同时,表现为一端穿孔板具尾,另一端不具尾(图1h),或一端穿孔板具长尾,另一端具短尾(图1 c,g)。
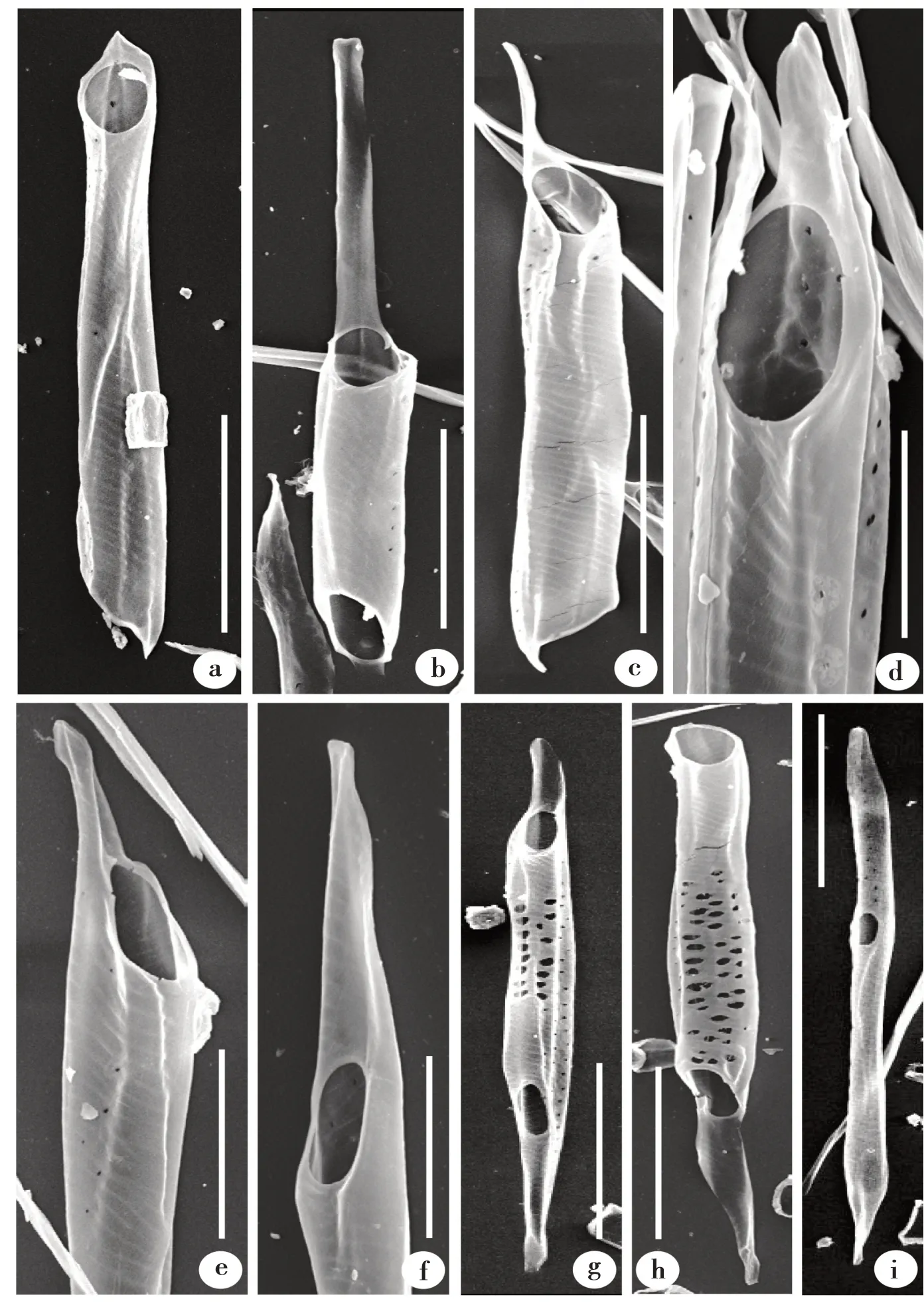
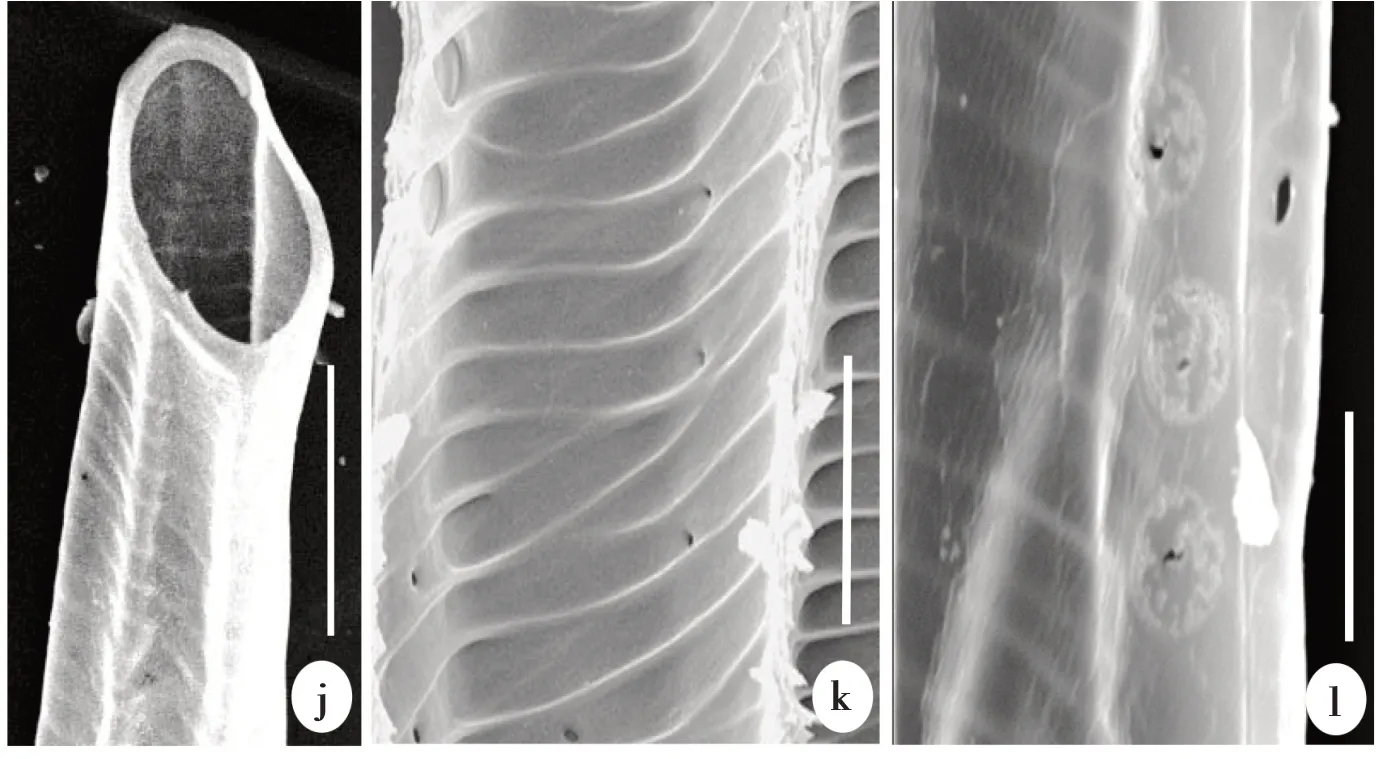
图1 蜡梅导管分子电镜照片Fig.1 SEM photos of vessel elements in C.praecox
观察中还发现有穿孔板位于导管侧壁(图1i),即侧壁穿孔板。导管内壁具螺纹加厚(图1k ~l),从导管内部切片上可更清楚地看到其形态(图1k)。
2.1.2 夏蜡梅导管分子形态观察
夏蜡梅导管分子端壁均为单穿孔板(图2),多数侧壁不具有明显螺纹加厚。穿孔板不具尾(图2a ~d)或具尾(图2e ~k)。不具尾的穿孔板通常较平截(图2a),有些有不同程度倾斜(图2b ~d)。穿孔板口部边缘内敛(图2a),石蜡切片法也观察到这种情况(图2c)。不具尾的穿孔板数量较多。
穿孔板尾的长度多数中等,形态较一致(图2g ~j)。少数穿孔板具短尾(图2e ~f)或长尾(图2k),这些穿孔板倾斜,但角度不大。导管分子两端的穿孔板形态可能相同(图2l),也可能不同(图2m)。石蜡切片法可观察到完整的穿孔板(图2n),其内部纹饰可见轻微加厚的较细螺纹(图2p)。
2.2 两种植物导管分子形态特征比较
蜡梅和夏蜡梅的导管分子均为单穿孔板,但两者有明显差异(图1 ~2,表1)。

夏蜡梅导管分子中,不具尾穿孔板占比较大,约为37%;穿孔板的尾普遍比蜡梅短,仅少数穿孔板具长尾;蜡梅导管分子中,穿孔板的尾普遍较长,形态各异,仅少数穿孔板不具尾。
蜡梅导管分子中的穿孔板多数倾斜,极少数平截;夏蜡梅导管分子中的穿孔板倾斜角度小,且较一致,平截的穿孔板较多。
导管内壁上,蜡梅具有明显的螺纹加厚,与木兰科(Magnoliaceae)的螺纹加厚较相似[19-20]。夏蜡梅的螺纹加厚不明显;在石蜡切片上,可观察到其内部纹饰不呈螺旋形。
2.3 统计分析
蜡梅导管分子长度为257 ~471 µm,平均为367.0µm,变异系数为15.9%(小于20%),变异幅度小,离散程度低;直径为21 ~60 µm,平均为40.4µm,变异系数为27.0%(大于20%),变异幅度较大,离散程度高(表1)。

表1 导管分子特征及统计分析Tab.1 Characters of vessel elements and statistical analysis
夏蜡梅导管分子长度为273 ~394 µm,平均为340.6µm,变异系数为13.3%(小于20%),变异幅度小,离散程度低;直径为34 ~61 µm,平均为51.1µm,变异系数为32.6%(大于20%),变异幅度较大,离散程度高。
单因素方差分析t检验结果显示,两种植物导管分子的平均长度差异不显著,平均直径差异显著(P<0.05)。
长度与直径的相关性检验结果显示,两种植物导管分子的长度与直径均呈极显著相关(P<0.01)。李红芳等[21]关于番荔枝科(Annonaceae)植物导管分子长度与直径的研究结果显示,导管分子的直径与长度不相关。导管分子的直径与长度是平行演化,各自独立进行,还是有相关性的共同演化,还需进行更多的研究来证实。
夏蜡梅导管分子的平均长度小于蜡梅,但差异不显著;蜡梅最短的导管分子比夏蜡梅短,最长的比夏蜡梅长,导管分子长度的变异系数大于夏蜡梅,离散程度比夏蜡梅高。夏蜡梅导管分子的最大直径与最小直径均大于蜡梅,导管分子直径的平均值显著大于蜡梅(P<0.05)。Metcalfe[12]研究中,蜡梅的直径和长度与本试验结果接近,直径稍大;美国夏蜡梅(Calycanthus floridus)的长度更长,直径比蜡梅小。
从以上蜡梅与夏蜡梅导管分子的比较可以看出,夏蜡梅的导管分子演化水平高于腊梅;两种植物导管分子端壁均为单穿孔板,具有种间或属间差异,穿孔板形态和导管的直径均有明显区别。这些解剖学特征具有分类学意义,一定程度上有助于进行种类鉴定。
3 讨论与结论
关于蜡梅科的系统位置,有很多争议,现在普遍认可其归属于樟目。在APG 系统中,蜡梅科的系统位置位于基部被子植物中的木兰分支[5],在基部类群中是较进化的类群。蜡梅与夏蜡梅导管分子端壁均为单穿孔板,为穿孔板演化的最高水平,该结果与蜡梅科在基部被子植物中的系统位置一致。
蜡梅科通常被认为包含2 ~4 属[22]。Takhtajan、Thome 和Dahlgren 等学者认为,蜡梅科包含4个属,分别为蜡梅属(Chimonanthus)、美国夏蜡梅属(Calycanthus)、中国夏蜡梅属(Sinocalycanthus)和Idiospermum[10,23-26]。其中,Idiospermum被一些学 者认为应当独立成科[3,27],但更多的人认为其应归属于蜡梅科[12,23,28-29],主张将其归入美国夏蜡梅属[12,23]。
另一个有争议的地方为中国夏蜡梅属(Sinocalycanthus)是否应当独立成属。Thome、Dahlgren、Mabberley 和吴征镒等学者根据解剖学特征认为,中国夏蜡梅属应独立成属[23-25,30-31];有些学者不支持独立成属,认为应将其并入美国夏蜡梅属[12,28-29]。APG系统将蜡梅科分为蜡梅属和夏蜡梅属。本研究根据APG 系统,按照《中国植物志》的分类方式,将中国夏蜡梅属归入美国夏蜡梅属,并按中国习惯称之为夏蜡梅属(Calyanthus)。
多数观点认为夏蜡梅属(包括美国夏蜡梅属(Calycanthus)和中国夏蜡梅属(Sinocalycanthus))较原始,蜡梅属较进化;形态学和孢粉学等特征表明夏蜡梅在演化上处于较原始的位置[7,28,32]。李烨等[33]研究发现,蜡梅属植物的叶脉类型较原始;李烨等[10]对蜡梅科的叶表皮细胞进行研究,认为蜡梅属植物的气孔器较原始,夏蜡梅属(Sinocalycanthus)和美国夏蜡梅属植物的气孔器较进化。本研究中,夏蜡梅的导管分子比蜡梅短且粗,穿孔板倾斜角度比蜡梅小,且穿孔板中不具尾的比例较高,显示夏蜡梅导管分子的演化水平比蜡梅更高,此结论与李烨等研究结果一致。
原始性状与进化性状可能在同一物种中并不统一,可能互相嵌合,不能单纯依据导管或其他木质部特征去推论夏蜡梅和蜡梅的演化水平,但这些特征可为系统学研究提供重要的参考。蜡梅科作为被子植物的基部类群,其穿孔板的相关特征对于导管的演化研究也有重要意义。
猜你喜欢
艺术家(2021年10期)2021-12-20
小学生学习指导(高年级)(2021年10期)2021-11-02
科学大众(2021年6期)2021-07-20
农产品市场周刊(2021年2期)2021-05-04
军工文化(2021年2期)2021-03-30
中华养生保健(2020年2期)2020-11-16
云南档案(2019年7期)2019-08-06
中医眼耳鼻喉杂志(2019年3期)2019-04-13
小天使·六年级语数英综合(2018年1期)2018-10-08
澳门月刊(2018年1期)2018-01-17
