
日本刀角瓢虫对不同番茄品种的产卵选择性及其影响因素
2022-07-12 02:44梅文娟张前荣丁雪玲卢学松姚凤銮何玉仙翁启勇
昆虫学报 2022年6期
梅文娟, 林 硕, 张前荣, 丁雪玲, 郑 宇,卢学松, 姚凤銮,*, 何玉仙,*, 翁启勇,*
(1. 福建省农业科学院植物保护研究所, 闽台作物有害生物生态防控国家重点实验室, 福州 350013;2. 福建省作物有害生物监测与治理重点实验室, 福州 350013; 3. 福建省作物有害生物绿色防控工程研究中心,福州 350013; 4. 福建省农业科学院作物研究所, 福州 350013)
番茄Lycopersiconesculentum作为病虫害多发的蔬菜作物,受烟粉虱Bemisiatabaci危害尤其严重(周涛等, 2010; 应丰泽等, 2015; 李娇娇等, 2018)。番茄中研究较多的抗虫性状主要是番茄叶毛特别是腺毛及其产生的次生代谢物(Simmons and Gurr, 2005; 郭广君, 2013)。众多研究表明,植物腺毛的分泌物通常会对昆虫产生驱避和毒害作用(Simmonsetal., 2004; Medeiros and Tingey, 2006),或具有黏性,对昆虫如烟粉虱、二斑叶螨Tetranychusurticae、小花蝽Oriusalbidipennis及捕食螨Phytoseiulusmacropilis等的存活、取食和生长发育产生消极影响 (Satoetal., 2011;Lucinietal.,2015;
Banihashemietal., 2017; Morimoto, 2019; Santegoetsetal., 2021)。番茄叶毛可以分为非腺毛和腺毛两大类,其中非腺毛包括Ⅱ, Ⅲ和Ⅴ型非腺毛,腺毛包括Ⅰ, Ⅳ, Ⅵ和Ⅶ型腺毛(Luckwill, 1943; Channarayappaetal., 1992; 郭广君, 2013)。在番茄腺毛中,Ⅳ型腺毛抗虫性的研究较多,它对银叶粉虱Bemisiaargentifolii、番茄潜麦蛾Tutaabsoluta及二斑叶螨都有显著抗性(Liedletal., 1995; Malufetal., 2010; Lucinietal., 2015)。
除了抵抗害虫外,番茄叶毛,尤其是腺毛也会削弱捕食者的搜索效率、捕食和繁殖能力(De Clercqetal., 2000; Kennedy, 2003; Banihashemietal., 2017; Bielzaetal., 2020)。例如,由于番茄上的腺毛更丰富,致使小花蝽在番茄植株上捕获的猎物数量显著少于在蚕豆和玉米上的(Coll and Ridgway, 1995)。番茄叶毛降低了捕食者加州新小绥螨Neoseiuluscalifornicus对二斑叶螨的捕食量(Cédolaetal., 2001)。番茄腺毛分泌物会黏住斑腹刺益蝽Podisusmaculiventris的足,影响其运动和捕食,甚至困死它们(Lambert, 2007)。小黑瓢虫Delphastuscatalinae在叶毛密度高的番茄品种上的产卵量显著低于叶毛密度低的番茄品种上的(Heinz and Zalom, 1996)。尽管如此,番茄腺毛对捕食性昆虫产卵选择行为的影响作用仍少见报道。
日本刀角瓢虫Serangiumjaponicum属鞘翅目(Coleoptera)瓢虫科(Coccinellidae),是烟粉虱的重要捕食性天敌,在叶片背面捕食烟粉虱且嗜食烟粉虱卵(姚凤銮等, 2018; Yaoetal., 2021)。本课题组前期研究发现日本刀角瓢虫对腺毛不发达的茄子品种的产卵选择性与茄子品种叶毛密度呈正相关关系(梅文娟等, 2021)。但是,日本刀角瓢虫对腺毛发达的番茄品种的产卵选择性还不得而知。因此,我们选择4个具有不同叶毛密度和类型的番茄品种作为供试寄主植物,观察其叶片上腺毛和非腺毛数量和形态,测定日本刀角瓢虫的产卵选择性,验证番茄叶毛类型及密度、日本刀角瓢虫的亲代和子代的表现、捕食及附着力与其产卵选择性之间的关系,以期进一步明确番茄叶毛尤其是腺毛对日本刀角瓢虫产卵选择的影响。
1 材料与方法
1.1 供试植物和昆虫
1.1.1供试番茄品种:在各番茄品种植株8叶期,高35~40 cm时(播种后8-9周),通过扫描电镜观察各番茄品种自下往上第7片羽状复叶背面叶毛密度后,从8个番茄品种(表1)中选取4个具有不同叶毛密度的番茄品种,分别是以色列超级金刚(Yiseliechaojijingang)(下面简称金刚Jingang)(四川种都高科种业有限公司)、砧木101(Zhenmu 101)、闽粉樱1号(Minfenying 1)(下面简称粉樱Fenying)和砧木301(Zhenmu 301)(福建省农业科学院作物研究所)。将无病、饱满的种子浸种催芽后播种于花盆(内径13 cm,高12 cm)中。每盆种植1株幼苗,待植株长到8叶期,株高35~40 cm时(播种后8-9周),去掉顶端叶芽和底部第1-3片羽状复叶待用。实验用苗均培养于温室中,温室环境条件为温度27±1℃、相对湿度65%±5%、光周期 14L∶10D。
1.1.2供试虫源:烟粉虱(约200头)采自福建省福州市郊区番茄(品种:爱好)田(26.133°N, 119.345°E),在室内棉花(石远321)苗上饲养4代;日本刀角瓢虫(约50头)采自福建省福州市郊区甘薯田(26.134°N, 119.344°E),在室内以接种烟粉虱的棉花苗饲养120代。 室内环境条件为温度27±1℃、相对湿度65%±5%、光周期 14L∶10D。
1.2 番茄品种叶片背面和日本刀角瓢虫跗节超微结构观察
选取处于8叶期的4个番茄品种的第7片羽状复叶进行扫描电镜(Zeiss Supra55)观察,每个番茄品种观察3株植株。避开主、次叶脉,选取叶脉之间的部分(10 mm×10 mm)进行制样。制样过程依次为:5%戍二醛固定样品4 h以上;磷酸缓冲液清洗样品3次,每次间隔10~15 min;1%锇酸固定4 h;蒸馏水清洗3次,每次间隔10~15 min;酒精逐级脱水(50%, 70%, 80%, 90%和100%,每次间隔 10~15 min,其中100%置换3次);环氧丙烷置换2次;临界点干燥(日立HITACHI HCP-2);上台、喷金(EIKO IB-5);上机观察。将番茄叶片背面叶毛分为腺毛Ⅰ, Ⅳ, Ⅵ和Ⅶ型以及非腺毛Ⅱ, Ⅲ和Ⅴ型(Luckwill, 1943; Simmons and Gurr, 2005),观察100倍镜下照片,统计15个不同位置1 mm2面积内的叶毛类型及数量。
选取在4个番茄品种叶片上饲养10~15 d的日本刀角瓢虫雌成虫各3头,解剖下后足跗节,然后上台、喷金(EIKO IB-5)、上机电镜观察跗节各部分形态(Yaoetal., 2021)。被污染后的日本刀角瓢虫跗节上的刚毛或爪上在扫描电镜下可观察到块状粘附物质。
1.3 日本刀角瓢虫对不同番茄品种的产卵选择性测定
采用叶碟法和植株法分别测定日本刀角瓢虫对番茄叶片和植株的产卵选择性(Yaoetal., 2021)。
1.3.1叶碟法:在直径33 mm培养皿内事先倒入12 g/L的琼脂水溶液并放置凝固(下称琼脂培养皿),再用打孔器分别将4个番茄品种的新鲜、干净叶片打成直径33 mm的叶碟,然后再将各小圆片正面朝下沿着培养皿内壁边缘铺在直径33 mm的琼脂培养皿中。再取100对混合日龄烟粉虱成虫置于其中,倒置并让烟粉虱产卵2 d,选卵量大于800粒的叶碟制成卵盘。剔除各叶片上的烟粉虱成虫,再将叶碟正面朝下沿着培养皿内壁边缘铺在直径90 mm的琼脂培养皿中,每皿内4个番茄品种的叶碟各放1片,叶碟间不接触,随机排列。每皿投入1头处于产卵期的10-15日龄的日本刀角瓢虫雌成虫,该虫饲养于棉花苗上(无特殊说明的话,下述供试日本刀角瓢虫均来源于棉花苗上),然后将培养皿倒置于温度27±1℃、相对湿度65%±5%、光周期14L∶10D(下述试验条件若无特殊说明则与此相同)的培养箱,24 h后检查4个番茄叶片上日本刀角瓢虫的产卵量。
1.3.2植株法:选取长势一致、高度相近的4个番茄品种植株,留下植株高35~40 cm处的一片羽状复叶。将各品种植株单株放入养虫笼内(36 cm×45 cm×45 cm, 下同),并接入500头混合日龄的烟粉虱产卵2 d后番茄叶片上的烟粉虱卵数量在800~1 200粒间。然后将这4个品种的番茄植株放在上述规格养虫笼的4个角落,再在养虫笼的中间地面上释放1头处于产卵期的10-15日龄的日本刀角瓢虫雌成虫,2 d后检查各番茄植物上日本刀角瓢虫卵的数量。
以上处理中,每头日本刀角瓢虫仅试验1次,各处理重复12次。
1.4 日本刀角瓢虫在不同番茄品种叶片上的生长发育和繁殖力测定
1.4.1生长发育:将日本刀角瓢虫置于直径33 mm培养皿(琼脂上铺有正面向下、带有600~800粒烟粉虱卵的各番茄品种叶片)内产卵,孵出幼虫单头饲养,每日更换一次猎物。每个处理重复50~65粒卵,每天定时观察记录各虫态发育历期和死亡数,直至羽化为成虫,并对新羽化成虫称量鲜重。
1.4.2繁殖:成虫羽化后,集中饲养于直径90 mm的琼脂培养皿中(培养皿中铺就了正面朝下、带有5 000粒左右烟粉虱卵的相应番茄品种叶片)。每日定时更换叶片,7 d后将日本刀角瓢虫单头饲养于直径33 mm的带有600~800粒烟粉虱卵的卵盘中(Yaoetal., 2021)。在解剖镜下观察记录日本刀角瓢虫的每日产卵量,连续观察记录30 d。每处理至少观测10头雌成虫。
1.5 日本刀角瓢虫在不同番茄品种叶片上的捕食能力和附着力测定
1.5.1捕食能力:在直径33 mm的卵盘中(烟粉虱卵量800~1 200粒)投入1头饥饿24 h的10-15日龄的雌成虫,24 h后在解剖镜下清点烟粉虱卵剩余数量,每处理至少观测6头雌成虫。
1.5.2附着力:利用自制离心式昆虫微力测试仪(Yaoetal., 2021),测试日本刀角瓢虫在4个番茄品种叶片上的附着力。首先把供试叶片叶背朝上平铺固定在测试转盘上(直径25.0 cm),然后将10-15日龄日本刀角瓢虫雌成虫称重后放置到供试叶片上,再缓慢加速,直到该虫脱离叶片。在这期间运用工业相机(WP-UT130/M, 深圳华谷动力科技有限公司)的连续拍照模式,记录日本刀角瓢虫在叶片上的附着情况。利用激光转速计[UT373, 优利德科技(中国)股份有限公司]记录转盘转速。最后选取虫体脱离叶片前后瞬间的图像,读取电机转速,并结合脱离前后瞬间的图像测量出虫体临界脱离半径,计算附着力。公式如下:
F=mrn2π2/900。
其中,F表示测试对象临界脱离测试材料表面时所受离心力,单位为N;m表示测试对象的质量,单位为kg;r表示测试对象脱离测试材料时的离心运动半径,单位为m;n表示测试对象脱离测试材料时驱动测试平台的电机转速,单位为r/min(王立新和翟利刚, 2014)。每个番茄品种上各测10头雌成虫,每头试虫仅测试1次。
1.6 日本刀角瓢虫对不同番茄品种叶片气味的趋性测定
日本刀角瓢虫对不同气味源选择反应分为两部分: (1)比较日本刀角瓢虫雌成虫对4种番茄健康叶片的选择性; (2)比较日本刀角瓢虫雌成虫对带有200~300粒烟粉虱卵和100~200头低龄若虫的4种番茄叶片的选择性。测试时实验室温度27±1℃、相对湿度65%±5%。 每个处理观察80头瓢虫。
测试前,接通大气采样仪的电源,摘取1片待测番茄羽状复叶,叶片面积为250~300 mm2,放入味源瓶中,然后通气10 min,空气依次通过活性碳瓶、蒸馏水瓶,味源瓶(或空白对照)进入多臂嗅觉仪中,使气味充满多臂嗅觉仪,以保证测试结果。测试时,调节气体流量计,使各臂的气流速度相同,均为0.3 L/min,打开日光灯,使光线均匀照射在嗅觉仪上。挑选10头活动性良好的10-15日龄日本刀角瓢虫雌成虫饥饿24 h后,置于嗅觉仪中心活动室内。若日本刀角瓢虫顺气流爬动,进入嗅觉仪管臂并停留5 min,即认定为有效选择。若日本刀角瓢虫未作出上述行为,则记为无选择(徐桂萍, 2011; 张辉洁等, 2012; 刘修堂等, 2013),每头日本刀角瓢虫雌成虫只记录首次有效选择。观察记录10头日本刀角瓢虫雌成虫在3 h内的选择情况。供试日本刀角瓢虫仅使用1次,试验重复8次。每次测试后味源瓶随机调换位置,并用100%乙醇和蒸馏水擦洗活动室、梨形玻璃球以及味源瓶(包括空白对照),以消除味源残留。
1.7 日本刀角瓢虫卵在不同番茄品种上的被捕食风险观测
1.7.1叶碟法:将4个番茄品种叶片按1.3节中的方法做成叶碟,铺在直径90 mm琼脂培养皿中。根据日本刀角瓢虫靠近叶脉产卵的习性,用阿拉伯胶将10粒24 h内产下的日本刀角瓢虫卵粘在叶碟背面的叶脉侧边。每皿投入1头10-15日龄的日本刀角瓢虫雌成虫,24 h后检查叶碟上剩余的日本刀角瓢虫卵量。
1.7.2植株法:选取1.3节中描述的番茄植株,并在4个番茄品种叶片背面的叶脉侧边上,用阿拉伯胶粘上10粒日本刀角瓢虫卵。然后,将这些植株放在上述规格养虫笼的4个角落,再在养虫笼的中间地面上释放1头10-15日龄的日本刀角瓢虫雌成虫,24 h后检查各番茄植株上日本刀角瓢虫卵的数量。
以上处理中,每头日本刀角瓢虫仅试验1次,各处理重复试验10次。
1.8 数据分析
将日本刀角瓢虫的产卵选择性、日本刀角瓢虫对4个番茄品种气味的选择、日本刀角瓢虫卵在4个番茄品种上的被捕食风险的数据以百分比形式记录,再进行Friedman检验(Lockwood, 1998)。日本刀角瓢虫在不同番茄品种叶片上的生长发育时间、成虫体重、单雌产卵量、附着力、烟粉虱卵被捕食量以及番茄叶毛密度采用单因素方差分析(One-way analysis of variance)。如果数据不能满足正态性和同质性,则采用非参数Kruskal-Wallis检验。日本刀角瓢虫在4个番茄品种上的存活率采用卡方(χ2)检验分析。单因素方差分析采用Tukey法进行多重比较,而Kruskal-Wallis检验则采用Bonferroni校正法。在相关性分析中,选择在不同处理间具有显著差异的数据进行皮尔逊Pearson相关系数分析(Scheirsetal., 2000)。所有统计分析均在R 语言(版本: 4.0.2)上进行。
2 结果
2.1 不同番茄品种叶片背面的超微结构
扫描电镜下观察发现,4个番茄品种叶片背面有3种非腺毛和4种腺毛,均呈线性,腺毛顶端具有腺体细胞(图1)。非腺毛包括Ⅱ型非腺毛(图1: C)、Ⅲ型非腺毛(图1: E)和Ⅴ型非腺毛(图1: G)。腺毛包括Ⅰ型腺毛(图1: B)、Ⅳ型腺毛(图1: D)、Ⅵ型腺毛(图1: F)和Ⅶ型腺毛(图1: H)。
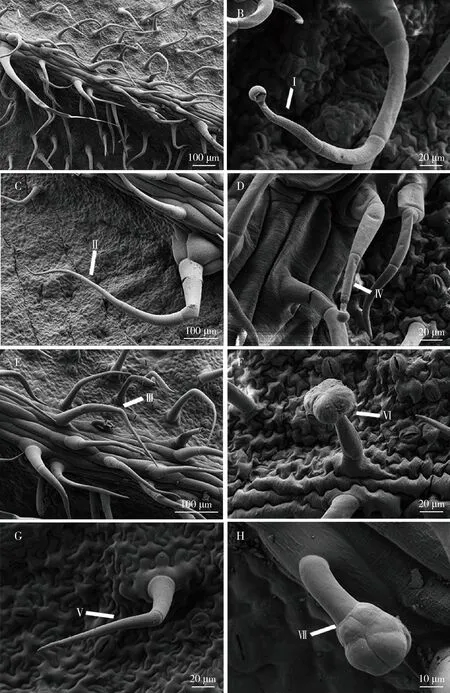
图1 4个番茄品种叶片背面叶毛扫描电镜图Fig. 1 Scanning electron microscopy images of trichomes on the abaxial leaf surface of four tomato varietiesA: 叶片表面概貌 General view of leaf surface; B: Ⅰ型腺毛Type Ⅰ glandular trichome; C: Ⅱ型非腺毛Type Ⅱ non-glandular trichome; D: Ⅳ型腺毛Type Ⅳ glandular trichome, E: Ⅲ型非腺毛Type Ⅲ non-glandular trichome; F: Ⅵ型腺毛Type Ⅵ glandular trichome; G: Ⅴ型非腺毛Type Ⅴ non-glandular trichome; H: Ⅶ型腺毛Type Ⅶ glandular trichome.
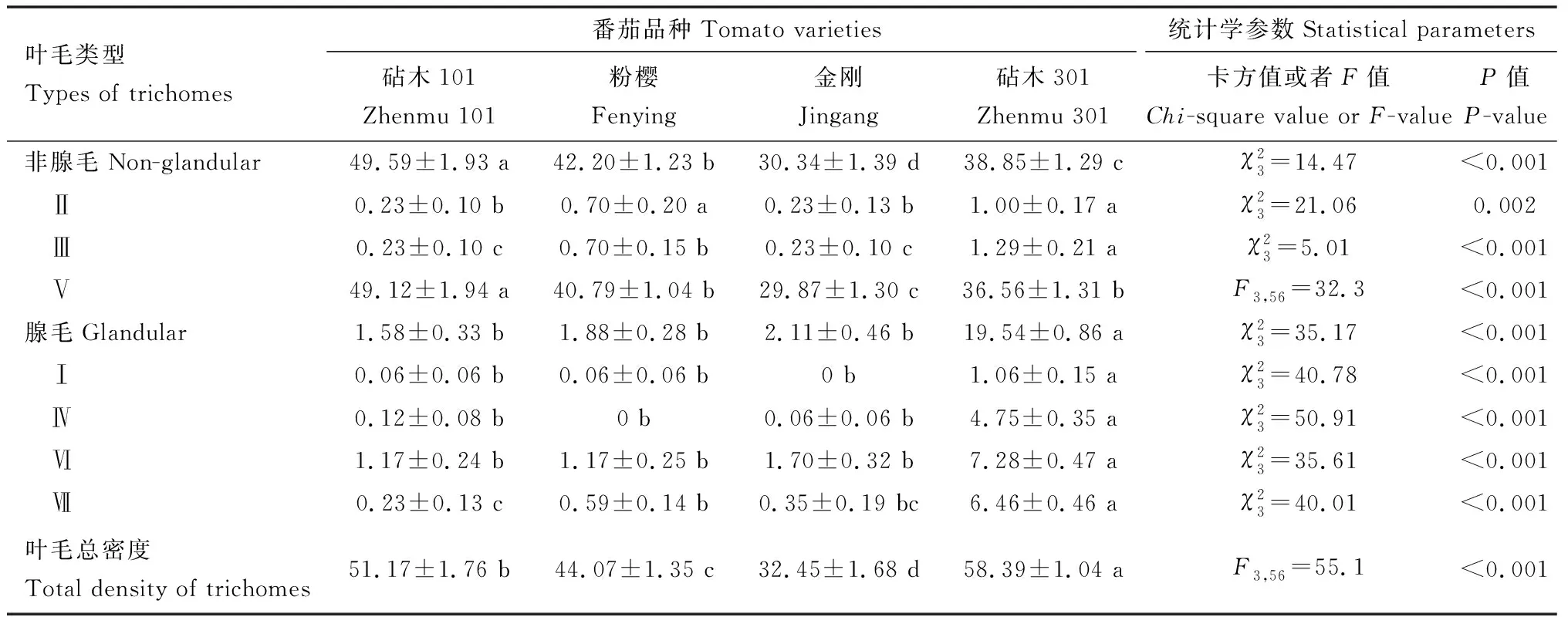
表1 4个番茄品种叶片背面叶毛类型及密度(根/mm2)Table 1 Types and densities (per mm2) of trichomes on the abaxial leaf surface of four tomato varieties
2.2 日本刀角瓢虫对不同番茄品种的产卵选择
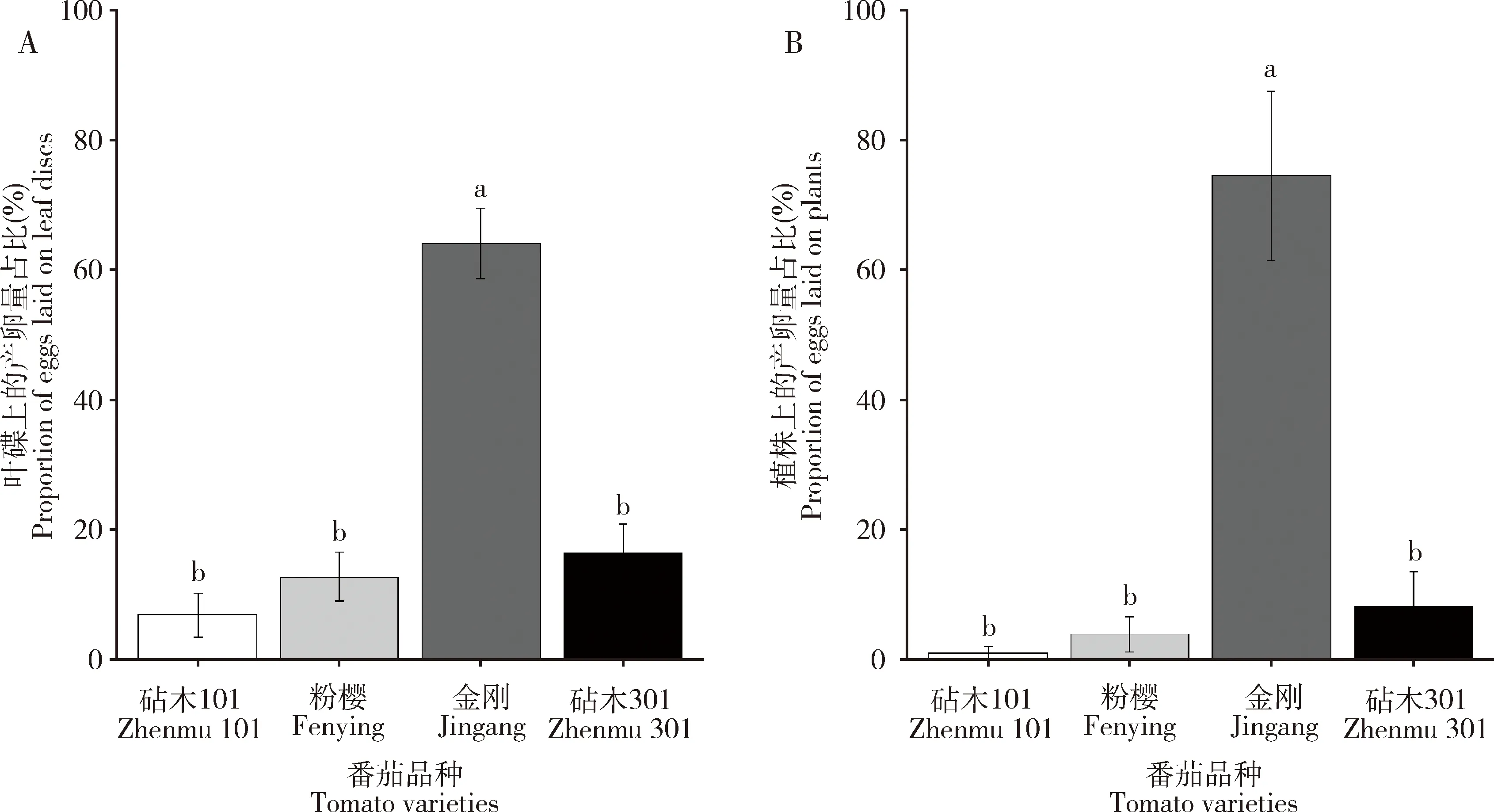
图2 日本刀角瓢虫雌成虫在带烟粉虱卵的4个番茄品种叶碟(A)和植株(B)上的产卵量占比Fig. 2 Proportions of eggs laid by female adults of Serangium japonicum on leaf discs (A) andplants (B) of four tomato varieties bearing Bemisia tabaci eggs图中数据为平均值±标准误;柱上不同小写字母表示差异显著(P<0.05, Tukey氏多重比较)。下图同。Data in the figure are mean±SE. Different small letters above bars indicate significant difference (P<0.05, Tukey’s multiple range comparison). The same for the following figures.
2.3 日本刀角瓢虫在不同番茄品种叶片上的生长发育、繁殖力
表2 非选择试验中日本刀角瓢虫在4个番茄品种上的未成熟期发育历期和存活率、羽化后成虫体重及雌成虫繁殖力和捕食量Table 2 Developmental duration and survival rates of immature stages, adult body masses, and fecundities andpredation amounts of female adults of Serangium japonicum on four tomato varieties in no-choice tests
2.4 日本刀角瓢虫在不同番茄品种叶片上的捕食、附着力及跗节超微结构

图3 日本刀角瓢虫在4个番茄品种叶片上的附着力Fig. 3 Attachment force of Serangium japonicumon leaves of four tomato varieties
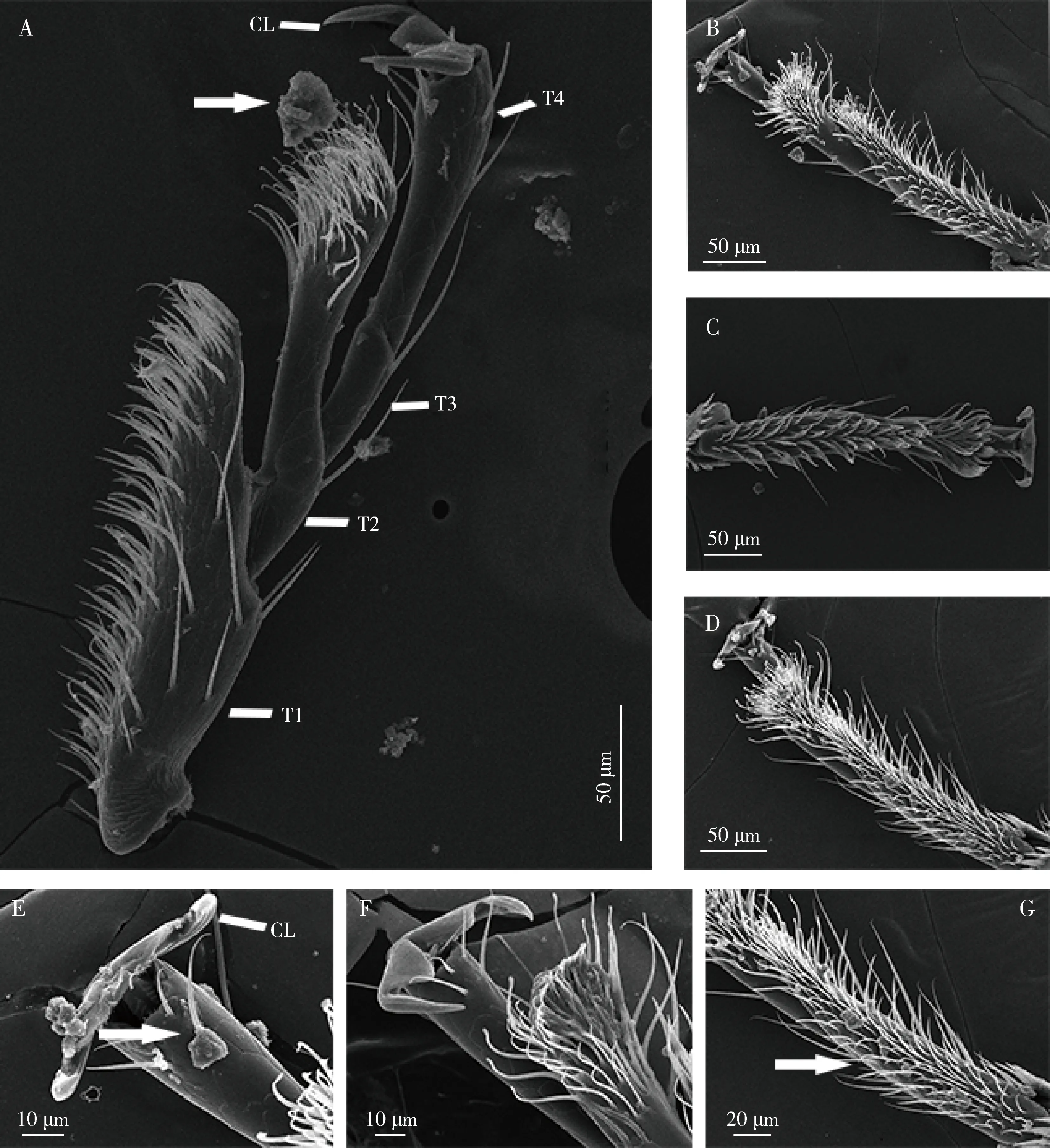
图4 日本刀角瓢虫雌成虫在4种番茄品种植株上饲养10-15 d后足跗节被污染情况的扫描电镜图Fig. 4 Scanning electron microscopy images of the contaminated hind-leg tarsi of female adults of Serangium japonicum rearedon plants of four tomato varieties for 10-15 dA-D: 分别是在金刚、粉樱、砧木101和砧木301番茄植株上饲养的日本刀角瓢虫跗节Hind-leg tarsus of S. japonicum reared on tomato varieties Jingang, Fenying, Zhenmu 101 and Zhenmu 301, respectively. E-G: 分别为瓢虫爪、第2跗节刚毛和第1跗节刚毛被污染情况Contaminated claw, and setae on the 2nd and the 1st tarsomere of S. japonicum, respectively. CL: 爪Claws; T1-T4: 分别为第1-4跗节1st-4th tarsomere, respectively. 箭头指示污染物Arrows indicate the contamination.
2.5 日本刀角瓢虫对不同番茄品种叶片气味的趋性
日本刀角瓢虫对4个番茄品种健康叶片和受烟粉虱为害叶片的气味均无特别偏好(图5)。日本刀角瓢虫对各番茄品种的叶片气味选择比例都稳定在5%~15%之间。在对健康叶片气味选择中,日本刀角瓢虫选择空白对照的比例(18.75%)小于未选择的比例(31.25%),但彼此之间无显著差异(P>0.05);在对烟粉虱为害叶片气味选择中,日本刀角瓢虫选择空白对照的比例(20%)、选择叶片的比例(5%~14%)和未选择的比例(46.25%)存在着显著差异(P<0.05),日本刀角瓢虫未选择的比例显著大于其他(P<0.05),日本刀角瓢虫选择空白对照的比例也显著大于对叶片的选择(P<0.05)。
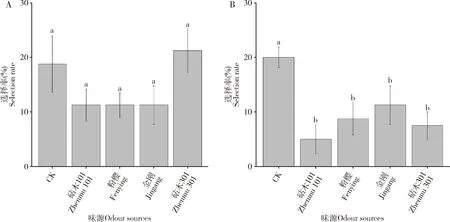
图5 日本刀角瓢虫对4个番茄品种健康叶片(A)和受烟粉虱为害叶片(B)气味选择Fig. 5 Selection rates of Serangium japonicum to the odours from healthy leaves (A)and Bemisia tabaci-infected leaves (B) of four tomato varietiesCK: 空白对照Blank control.
2.6 日本刀角瓢虫卵在不同番茄品种上的被捕食风险
在叶碟法中,日本刀角瓢虫卵在4个番茄品种上的被捕食比例差异显著(F3,36=15.98,P=0.001)(图6: A),日本刀角瓢虫卵在金刚上的被捕食风险(65.83%)显著高于在其他3个番茄品种上的(砧木101: 0, 粉樱: 1.67%, 砧木301: 7.5%)(P<0.05)。日本刀角瓢虫卵在砧木101、粉樱、砧木301间上的被捕食比例无显著差异(P>0.05)。在植株法中,瓢虫卵在4个番茄品种上的被捕食比例差异显著(F3,36=17.58,P<0.001)(图6: B),且以金刚上的(65.43%)显著高于其他3个番茄品种上的(<10%),砧木301上的(28.72%)显著高于粉樱(4.07%)和砧木101(1.78%)上的(P<0.05)。
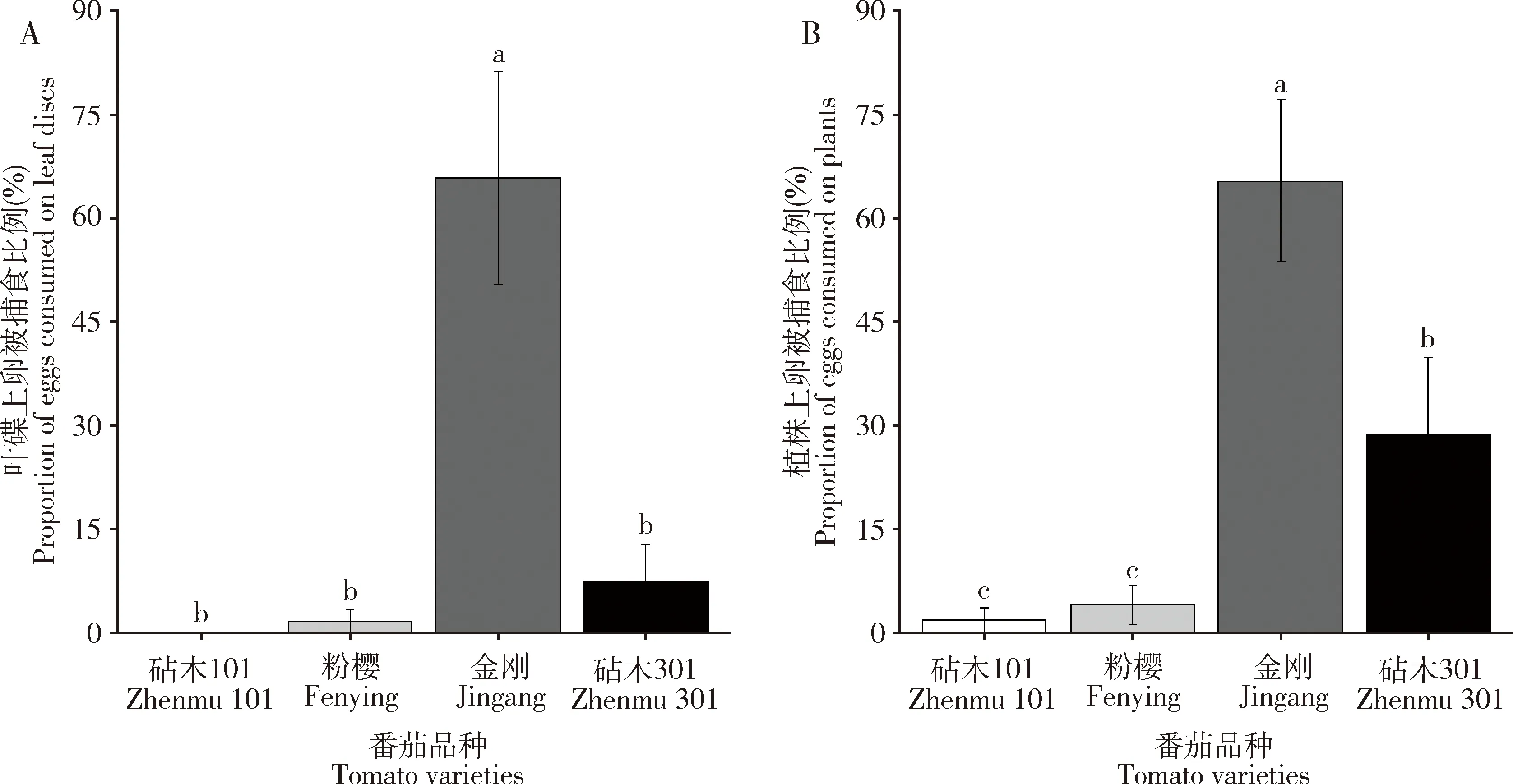
图6 4个番茄品种叶碟(A)和植株(B)上的日本刀角瓢虫卵被同类捕食比例Fig. 6 Proportions of eggs consumed by cannibalistic Serangium japonicum on leaf discs (A)and plants (B) of four tomato varieties
2.7 不同番茄品种叶毛密度、日本刀角瓢虫产卵选择性以及日本刀角瓢虫表现之间的相关性分析
选取存在显著差异的变量分别与日本刀角瓢虫在不同番茄品种上的产卵量占比(产卵选择性)进行相关性分析(表3)。日本刀角瓢虫后代的生长发育、成虫的繁殖、捕食能力和附着力均与日本刀角瓢虫雌成虫的产卵量占比无显著相关性。日本刀角瓢虫在不同番茄品种上的产卵量占比与其卵被捕食比例(被捕食风险)(叶碟法:r=0.99,P=0.003; 植株法:r=0.99,P=0.011)存在着显著的正相关关系。
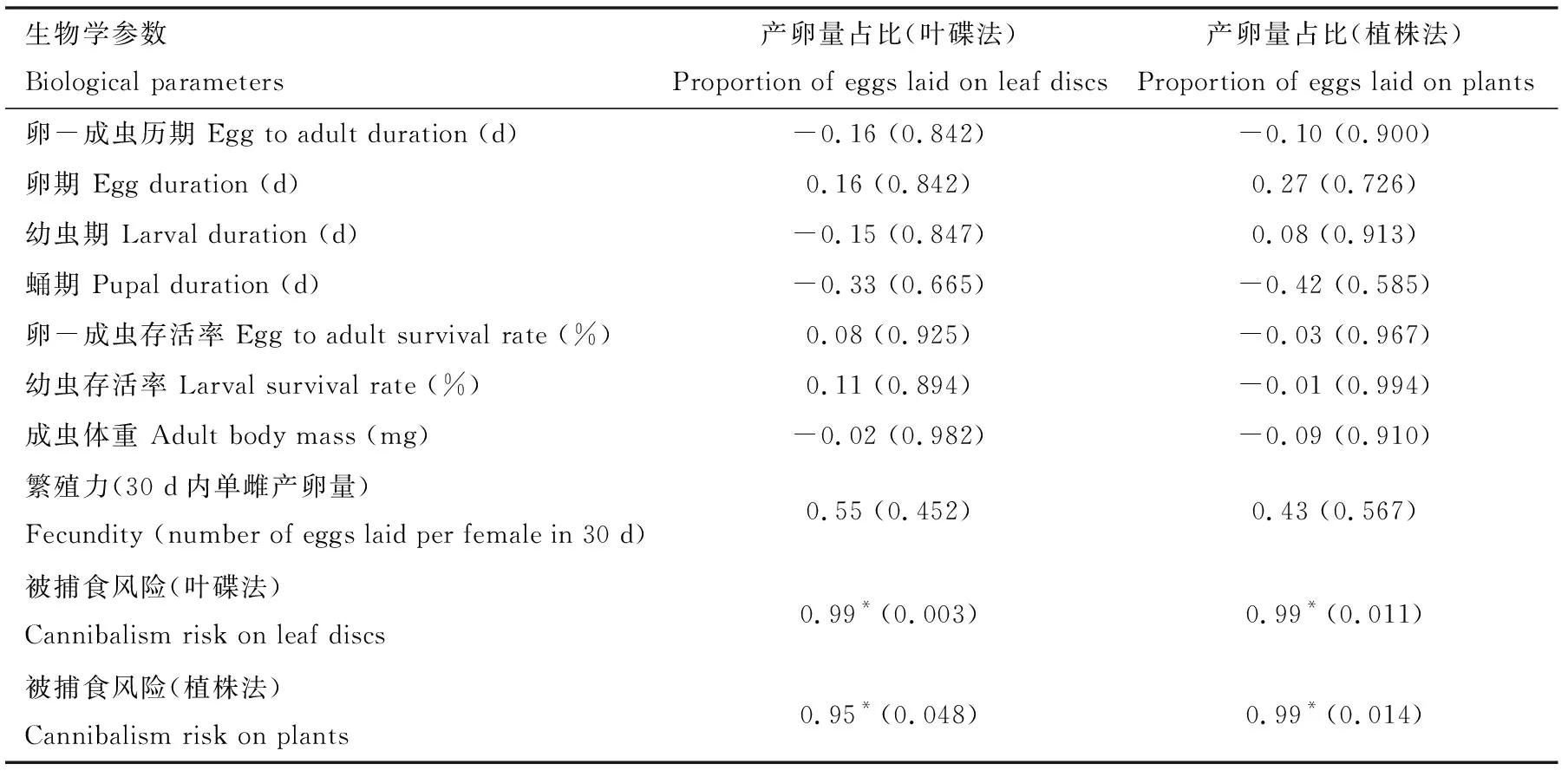
表3 日本刀角瓢虫在4个蕃茄品种上的产卵量占比与子代及成虫表现的相关性Table 3 Correlations between the proportion of eggs laid of Serangium japonicum and its offspringand adult performance on four tomato varieties
日本刀角瓢虫在不同叶片上的繁殖力与4个番茄品种上Ⅰ型腺毛密度(r=-0.95,P=0.048)呈显著负相关,而与其他叶毛类型的密度无显著相关性(表4)。
3 讨论
本研究中,日本刀角瓢虫在4个番茄品种上的产卵选择差异显著,在叶碟和植株法中,日本刀角瓢虫都偏好在金刚品种上产卵(图2)。但是金刚品种上叶毛总密度和非腺毛密度在4个番茄品种中最低(表1),因此,日本刀角瓢虫雌成虫可能偏好选择叶毛密度低的番茄叶片进行产卵。这与我们在茄子品种间观测到日本刀角瓢虫偏好在叶毛密度最高的黑到底品种上产卵的结果(梅文娟等, 2021)相反,究其原因,可能与番茄上存在大量的腺毛有关。
番茄腺毛会产生并储存大量的化合物,这些化合物在植物防御中起着重要作用(Shepherdetal., 2005; Shepherd and Wagner, 2007; Peifferetal., 2009; Tookeretal., 2010)。番茄上的Ⅰ, Ⅳ和Ⅵ型腺毛的密度与红蜘蛛的数量和产卵量呈负相关(Savietal., 2019)。植物腺毛除了抵抗害虫外,还可能对捕食性天敌产生不利影响(Sletvoldetal., 2010)。例如,番茄腺毛显著降低捕食螨的捕食作用(Cédolaetal., 2001; Satoetal., 2011)。在本研究中,Ⅰ型腺毛密度与日本刀角瓢虫繁殖力呈显著负相关关系(表4),且在日本刀角瓢虫偏好产卵的金刚品种上无Ⅰ型腺毛(表1),说明Ⅰ型腺毛很可能可以抑制日本刀角瓢虫雌成虫的产卵。Ⅳ型腺毛是番茄抗性品种上腺体叶毛的重要组成部分,其分泌物具毒性可以对植食性昆虫产生直接致死作用(Williamsetal., 1980; Kennedy, 2003; de Resendeetal., 2008; Keskin and Kumral, 2015)。在测试的4个番茄品种上,仅在粉樱品种上未发现Ⅳ型腺毛,那么,日本刀角瓢虫后代在粉樱品种上相对良好的生长发育状态很可能与此相关。Ⅰ型和Ⅳ型腺毛对日本刀角瓢虫生长发育、繁殖的作用值得进一步深入探讨。
多项研究指出,番茄腺毛的分泌物也可能通过改变昆虫的附着能力影响昆虫的运动和捕食,进而影响昆虫的产卵选择行为。斑腹刺益蝽、盲蝽Macrolophuspygmaeus和毛肩花蝽Oriusniger,以及二星瓢虫Adaliabipunctata幼虫都会受到腺毛分泌物的粘附而减弱运动能力,增加用于清理的时间,从而削弱捕食能力(Shah, 1982; Economouetal., 2006; Lambert, 2007; Riddick and Simmons, 2014)。本研究发现,日本刀角瓢虫在番茄叶片上的爬行速度较为缓慢。扫描电镜观察发现日本刀角瓢虫在番茄叶片上爬行后,其跗节上的爪及刚毛受到严重污染(图4),导致日本刀角瓢虫在番茄叶片上的附着力(64~115 μN)较在光滑的亚克力板上的附着力(233.63 μN)(Yaoetal., 2021)显著下降,这可能也驱使日本刀角瓢虫逃离腺毛丰富的番茄品种。
此外, Ⅴ型非腺毛有利于昆虫雌虫的定居和产卵(Gurr and McGrath, 2002; Simmons and Gurr, 2005; McDowelletal., 2011)。Alba等(2009)研究证实了番茄的Ⅴ型非腺毛密度与二斑叶螨的产卵量以及潜叶蛾Phthorimaeaoperculella幼虫的存活率存在正相关关系。而且在番茄非腺毛中,通常Ⅴ型非腺毛密度最高(McDowelletal., 2011; 郭广君, 2013; Savietal., 2019),本研究亦发现Ⅴ型非腺毛在4个番茄品种的叶毛数量中占绝大多数(表1)。然而,在本研究中,Ⅴ型非腺毛与日本刀角瓢虫产卵选择无明显相关关系,尤其是日本刀角瓢虫产卵偏好的金刚上的Ⅴ型非腺毛密度还显著低于其他3种番茄品种的Ⅴ型非腺毛密度(表1; 图2)。因此,可能原因是尽管Ⅴ型非腺毛对于吸引日本刀角瓢虫产卵具有积极作用,但腺毛对日本刀角瓢虫产卵选择的排斥作用更强烈。
日本刀角瓢虫对4个番茄品种的叶片气味无明显偏好(图5),说明番茄的叶片气味并不是影响日本刀角瓢虫产卵选择的重要因素。本课题组前期研究与Tian等(2020)的研究报道均表明日本刀角瓢虫对植物气味不敏感。另外,由于4个番茄品种上烟粉虱2日龄卵的长度和宽度之间均无显著差异(未发表数据)。因此,猎物的营养状态很可能不是影响日本刀角瓢虫产卵选择性的关键因子。当然,本研究中烟粉虱经历转寄主饲养后,即先从番茄上采集,再用棉花饲养多代,再接到不同品种的番茄上,可能导致烟粉虱所产下的卵在不同品种间无显著差异。因此,采用长期饲养在各自番茄品种上的烟粉虱来制作卵盘,供日本刀角瓢虫取食,那么就能消除可能存在的烟粉虱转寄主适应的影响,从而更好地反映日本刀角瓢虫对不同番茄品种的选择性和适应性。
与日本刀角瓢虫成虫偏好在金刚上产卵不同,日本刀角瓢虫子代在粉樱品种上由卵发育至成虫羽化的历期最短,幼虫存活率最高,羽化后成虫体重也最大;亲代雌成虫在金刚品种上的产卵量及对烟粉虱卵的捕食量均与粉樱和砧木101品种上的差异不显著(图2)。因此,日本刀角瓢虫雌成虫和幼虫对番茄叶片的需求和适应性可能是有差异的。Scheirs和De Bruyn(2002)认为应该综合考虑后代的生长发育以及成虫的繁殖力才能更好地理解昆虫的产卵选择。然而,在我们的研究中,如果综合考虑子代和亲代的表现,日本刀角瓢虫在粉樱品种上的生存适合度似乎并不比在金刚品种上的差,而且日本刀角瓢虫卵在4个番茄品种叶碟和植株上的被捕食风险与日本刀角瓢虫的产卵选择性成显著正相关关系(表4),说明日本刀角瓢虫对番茄叶片的产卵选择并不是基于为了保护后代免受同类个体的捕食,也暗示日本刀角瓢虫喜欢呆在金刚植株上,或者在金刚植株上的活动(包括产卵)更不容易受到干扰。可见,在番茄等具丰富腺毛的植物上,日本刀角瓢虫的产卵选择更可能考虑亲代的生存需求,比较符合最适取食理论(Scheirsetal., 2000)。
综上,对于具备腺毛的叶片,日本刀角瓢虫可能倾向于选择叶毛少的叶片进行产卵。该结果也与我们在烟草上的发现一致,烟草腺毛丰富,日本刀角瓢虫成虫及幼虫都不能在烟草上进行长时间的生命活动。所以日本刀角瓢虫亲代成虫的繁殖以及后代幼虫的生长发育可能更多地受到腺毛的影响。区分腺毛和非腺毛有助于正确认识叶毛对天敌行为的影响机制,为生产上天敌实际应用提供一定的指导意义。
猜你喜欢
红蜻蜓(2020年9期)2020-09-26
作文大王·笑话大王(2018年12期)2018-03-23
现代农业科技(2017年13期)2017-08-09
农业科技与装备(2017年1期)2017-06-09
现代园艺(2016年7期)2017-01-09
现代园艺(2016年9期)2016-07-16
小学生导刊(低年级)(2016年5期)2016-05-27
小朋友·快乐手工(2015年10期)2015-11-02
创新作文(5-6年级)(2015年9期)2015-10-21
红蜻蜓(2015年4期)2015-06-01
