
新疆两个主栽核桃品种叶片结构和光合特性的比较
2022-07-20 09:40韩立群李丽莉
经济林研究 2022年2期
张 玉,韩立群,,赵 钰,于 栋,李丽莉,周 龙,马 凯
(1. 新疆农业大学 园艺学院,新疆 乌鲁木齐 830052;2. 新疆农业科学院 a. 园艺作物研究所;b. 农业农村部新疆地区果树科学观测试验站,新疆 乌鲁木齐 830091)
核桃Juglans regiaL.系胡桃科核桃属多年生落叶果树,位居世界四大干果(核桃、扁桃、板栗、腰果)之首,是我国重要的木本油料树种,具有很高的经济价值[1]。核桃仁富含多种不饱和脂肪酸和矿质元素等营养物质,具有抗衰老、提高免疫力等保健功效[2-3]。新疆维吾尔自治区是我国核桃种植大省区,核桃产业是新疆特色林果业之一,发展特色林果业是带动经济增长的新动力,不仅能促使农民增收致富,而且能促进新疆林果产业提质增效,所以,如何提高核桃产量的问题是目前亟待解决的现实问题。而果树产量与其基因、器官形态和组织结构等内在因素有关,而光合作用是影响果树产量的决定性因素,因此,提高植株的光合速率是提高果树产量的关键措施[4]。
光合作用是植物生长发育、产量形成与增加的关键影响因素[5],叶片是进行光合作用的主要器官,其结构和功能与光合作用的关系均十分密切。有关研究结果表明:植株种内光合速率的大小是由测定方法和叶片内部显微结构的异同造成的[6],叶片的解剖结构与其光合特性紧密相关,其净光合速率与叶片的栅栏组织、海绵组织的厚度及气孔密度均呈正相关,即栅栏组织和海绵组织越厚,净光合速率就越高[7];邱彦芬等[6]在研究橡胶树时发现,胞间CO2浓度与角质层和叶片厚度均呈极显著负相关;任顺梅等[8]在研究地石榴时发现,净光合速率与气孔导度、蒸腾速率均呈极显著正相关,而与胞间CO2浓度呈极显著负相关。
目前,有关不同核桃品种光合特性日变化规律的研究报道较多:宗建伟等[9]、王陆军等[10]和刘鹏等[11]研究了不同核桃品种光合特性的变化规律;杨莹等[12]对红、绿核桃叶片的性状特征及光合日变化趋势进行了比较分析;李秋煜等[13]研究了早实和晚实核桃的光合特性。目前,关于核桃光合作用与环境因子之间变化关系的研究报道较多[14-15],然而,有关相同生境下不同核桃品种光合特性与叶片的显微结构和叶绿素相对含量之间的关系的研究报道并不多见。为给新疆地区早实核桃的丰产和优质栽培提供理论依据,本研究对新疆地区现有优良核桃品种‘温185’和‘新新2 号’的叶片形态结构与光合特性等方面进行了比较分析,现将研究结果分析报道如下。
1 材料与方法
1.1 试验地概况
试验地位于新疆维吾尔自治区喀什地区叶城县农业农村部果树科学观测站。该地区属于温带干旱气候,光热量资源丰富,日照时间长,降水稀少,昼夜温差大,年平均气温11.3 ℃,年平均降水量54 mm,平均年无霜期约228 d。试验地的土壤是沙壤土,土壤中的有机质含量为2.954 g/kg,其pH 值为7.3 ~8.0。
1.2 材 料
试材为7 年生的‘温185’和‘新新2 号’实生大树,南北向定植,常规管理,株行距均为6 m×3 m。
1.3 方 法
1.3.1 叶片形态及显微结构的观测
叶片形态的观测:2021 年7 月,选择长势一致的‘温185’和‘新新2 号’核桃树,每个品种各选3 株样树,分别从核桃样树的东、南、西、北4 个方向各标记2 片中部新梢叶片的旗叶,共设8 次生物学重复,每隔15 d 用游标卡尺测量旗叶的叶长(即叶片顶端到叶柄与叶片连接处的长度)和叶宽(即叶片最宽处的宽度),并参照吉德娟等[16]和王磊等[17]的计算方法分别计算叶形指数和叶面积:叶形指数=叶长/叶宽;叶面积=叶长×叶宽×2/3。
叶片解剖结构的观察:用孔径为0.7 cm 的打孔器分别对‘温185’和‘新新2 号’叶片的旗叶进行打孔。利用常规石蜡切片法[18]制片,将其叶片放入70%的FAA 固定液中保存以备用,脱水、浸蜡和包埋,利用切片机(LEACA CM1950)切片,切片厚度为10 μm,用1%的番红和1%的固绿进行染色,用中性树胶封片,并标注好样品的名称和采样时间,然后,用显微镜(NIKON eclipse Ts2)观察并拍照,利用Image J 软件测量叶片上表皮、下表皮、栅栏组织、海绵组织的厚度和叶片厚度等微观结构指标,并计算出栅海比、叶片组织疏松度和叶片组织紧密度:栅海比=栅栏组织/海绵组织;叶片组织疏松度=海绵组织/叶片厚度;叶片组织紧密度=栅栏组织/叶片厚度。
1.3.2 光合参数的测定
于2021 年7 月22—24 日(天气晴朗),选择健康且长势一致的样树测定其光合参数,每株样树各测定5 片树冠中层不同方向且健康的完整旗叶,设3 次生物学重复。采用SPAD-502 和Li-6800 光合仪分别测定旗叶的叶绿素相对含量(SPAD 值)和净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)、光合有效辐射强度(PAR)等光合参数与温度(T)、CO2浓度等环境因子,光合参数的测定时刻为9:00—21:00时,每隔2 h 测定1 次,取平均值,并根据戴秋月等[19]所用的方法计算水分利用效率(RWUE):
RWUE=Pn/Tr。
1.4 数据统计与分析
运 用SPSS 26.0 和Microsoft Excel 2010 软 件进行数据处理和单因素方差分析(显著性水平为0.05)与相关性分析及制作图表;运用Image J 软件在显微镜(20 倍)下测量上表皮、下表皮、栅栏组织、海绵组织的厚度和叶片厚度等指标。
2 结果与分析
2.1 不同核桃品种叶片形态与显微结构的观测结果
不同日期测定的不同核桃品种的叶长和叶宽如图1 所示,其叶形指数和叶面积如图2 所示。从图1 和图2 中可以看出,‘温185’的叶长、叶宽和叶面积均略大于‘新新2 号’的;‘新新2 号’的叶形指数略大于‘温185’的。两个核桃品种的叶长存在显著性差异(P<0.05),而其叶宽、叶形指数和叶面积均不存在显著性差异(P>0.05)。
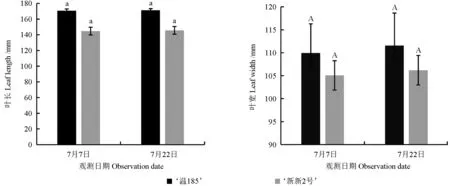
图1 不同核桃品种叶长和叶宽的测定结果Fig. 1 Determination results of leaf length and leaf width of different walnut varieties
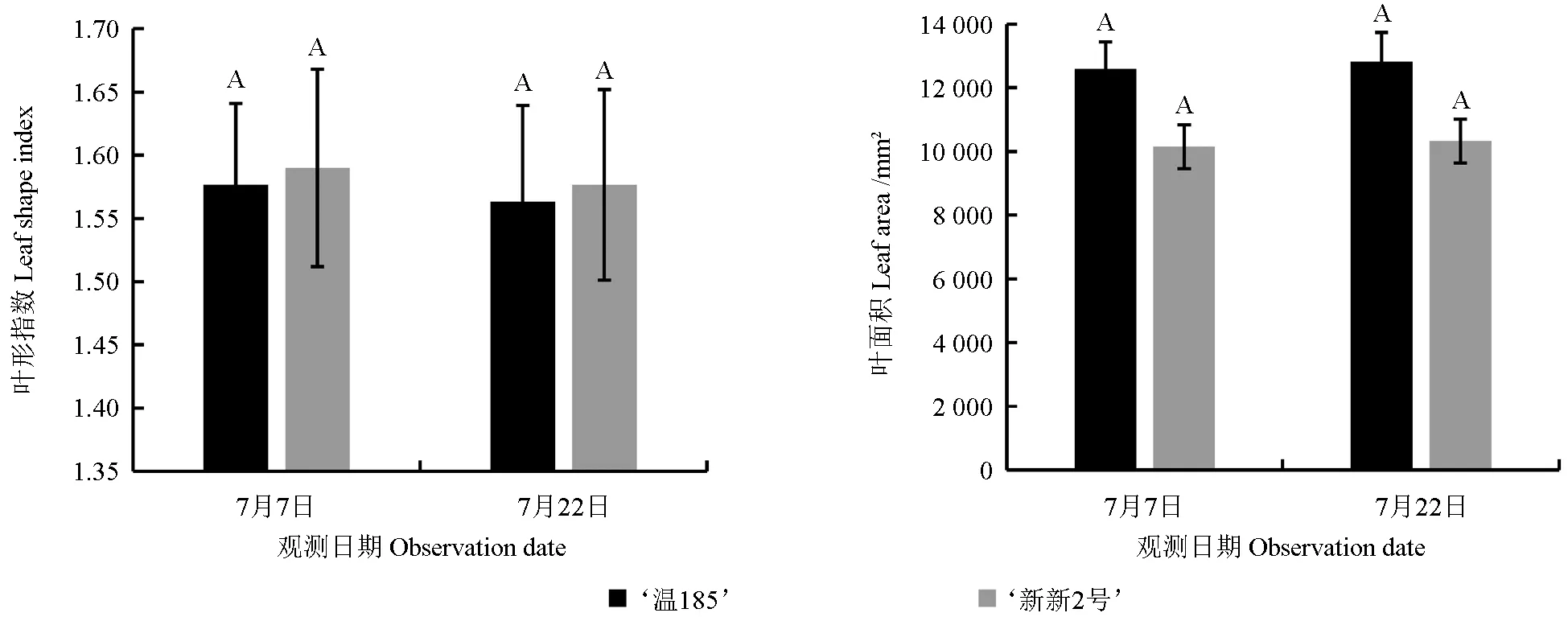
图2 不同核桃品种叶形指数和叶面积的测定结果Fig. 2 Comparison results of leaf shape index and leaf area of different walnut varieties
观察到的‘温185’和‘新新2’的叶片纵切面的显微结构如图3 所示。从图3 中可以看出,从外向内可以观察到,核桃叶片的最外层是由大小不等且透明的1 列椭圆形细胞构成上下表皮的,其内层是由颜色较深的2 列长条状细胞组成的栅栏组织,有多个由不规则细胞组成的海绵组织分布在栅栏组织与下表皮之间。‘温185’的海绵组织明显少于‘新新2 号’的,‘新新2 号’叶片的空腔大于‘温185’的。
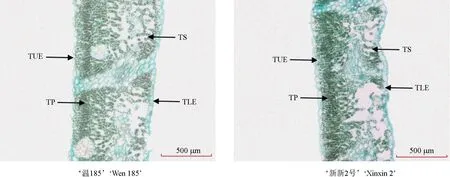
图3 不同核桃品种的叶片显微结构Fig. 3 Leaf microstructure of different walnut varieties
采用单因素检验法对‘温185’和‘新新2 号’叶片的解剖结构指标值进行单因素方差分析,结果见表1。由表1 可知,‘温185’的上表皮厚度、下表皮厚度、栅栏组织厚度、叶片厚度、栅海比、叶片组织紧密度均大于‘新新2 号’的;‘新新2号’的海绵组织厚度大于‘温185’的;‘新新2 号’的海绵组织和叶片组织疏松度均大于‘温185’的。‘温185’和‘新新2 号’的8 个叶片解剖结构指标值均无显著差异(P>0.05)。
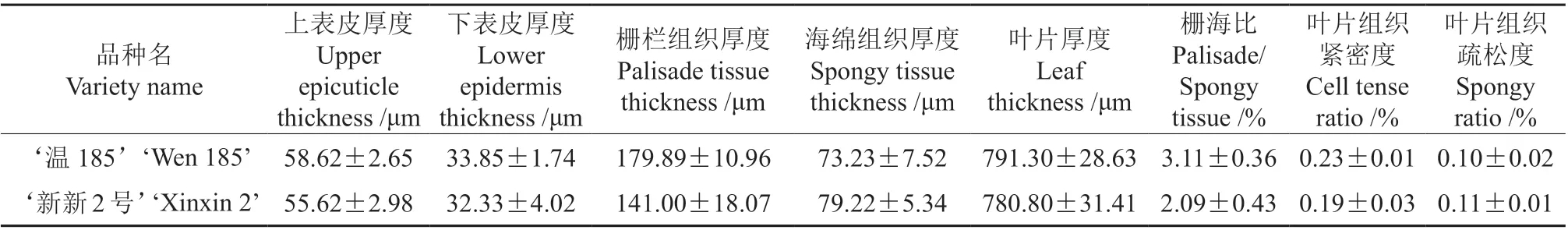
表1 不同品种核桃叶片解剖结构的观测结果†Table 1 Significance analysis of leaf anatomical structure of two varieties of walnut
2.2 不同核桃品种叶绿素相对含量和光合参数的比较分析
2.2.1 不同核桃品种叶绿素相对含量的比较分析
叶绿素是植物细胞内参与光合作用的主要色素,直接参与光合作用过程,叶绿素含量的高低决定着植物光合能力的强弱[20]。不同核桃品种的叶绿素相对含量(SPAD 值)如图4 所示。从图4中可以看出,‘温185’和‘新新2 号’的叶绿素相对含量不同,但其含量差异不明显,‘温185’的叶绿素相对含量略高于‘新新2 号’的。

图4 不同核桃品种的叶绿素相对含量比较Fig. 4 Comparison of chlorophyll relative content in different walnut varieties
2.2.2 不同核桃品种各个光合参数的比较分析
2.2.2.1 各个环境因子的比较分析
光合作用是叶绿素吸收和利用光能,将环境中的二氧化碳和水合成植株生长发育所需的营养物质,因此光合作用与环境因子密切相关[21]。不同核桃品种的光合有效辐射、空气温度和CO2浓度的日变化趋势如图5 所示。从图5 中可以看出,‘温185’和‘新新2 号’核桃的光合有效辐射(PAR)与气温的日变化趋势均相似,均呈现出先上升至平稳状态最后下降的变化趋势;其PAR 在15:00时均最大,均为1 809 μmol·m-2·s-1;气温在17:00时达到最高,为38.96 ℃;其大气中CO2的浓度在上午9:00时均最大,为386.4 μmol·mol-¹,在9:00—11:00 时均呈明显的下降趋势,在11:00—17:00 时均表现为上下波动状态,而在19:00—21:00 时均趋于平缓。
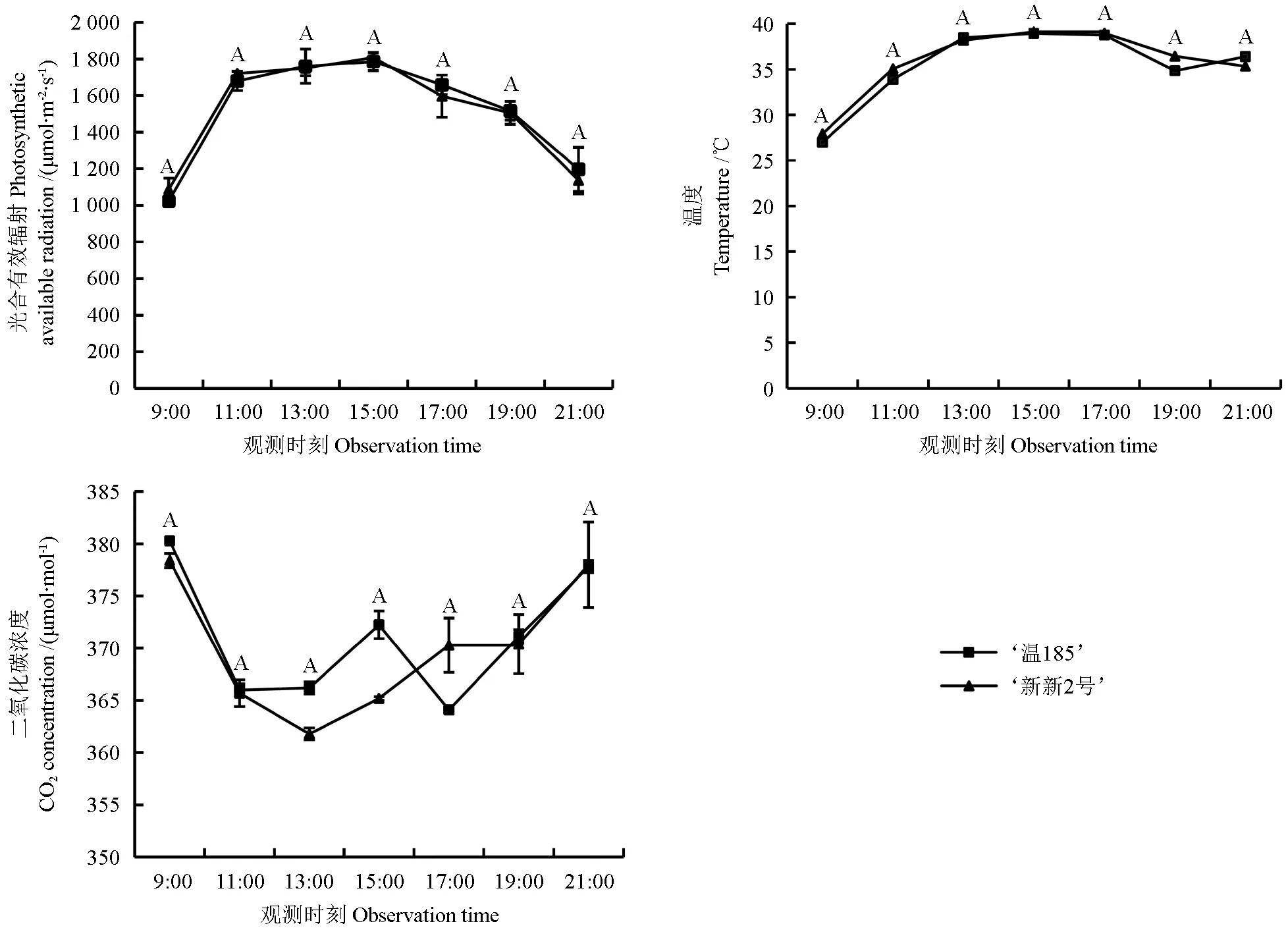
图5 不同核桃品种的光合有效辐射、温度和CO2浓度的日变化趋势Fig. 5 Diurnal variation trend of photosynthetic available radiation, temperature and CO2 concentration of different walnut varieties
2.2.2.2 各个光合参数的比较分析
‘温185’和‘新新2 号’核桃品种的净光合速率日变化趋势如图6 所示。从图6 中可以看出,‘温185’和‘新新2 号’的净光合速率变化曲线均为单峰型,且‘温185’的净光合速率日均值(9.89 μmol·m-2·s-1)大于‘新新2 号’的(8.68 μmol·m-2·s-1); 但 是,‘ 新 新2 号’ 的最大净光合速率出现的时间比‘温185’的早,‘温185’的净光合速率在13:00 时达到了峰值(13.12 μmol·m-2·s-1),而‘新新2 号’的净光合速率在11:00 时就达到了峰值(11.56 μmol·m-2·s-1),随着时间的推移,两者的净光合速率在达到峰值后均缓慢减小;且2 个品种间其净光合速率在13:00、15:00、17:00 和21:00 时测定的值均存在显著差异(P<0.05)。
气孔是叶片内部与外界O2、CO2等气体交流的主要通道[22],气孔导度(Gs)的大小可以反映植物光合作用的强弱[23]。‘温185’和‘新新2 号’核桃品种的气孔导度日变化趋势如图7 所示。从图7 中可以看出,‘温185’的Gs日均值(0.223 mol·m-2·s-1)大于‘新新2 号’的Gs日均值(0.170 mol·m-2·s-1),且两者的气孔导度均呈现出先增加再减小的变化趋势;‘温185’与‘新新2 号’的气孔导度峰值出现的时刻不一致,‘温185’的气孔导度峰值(0.270 mol·m-2·s-1)出现在13:00 时,而‘新新2 号’的气孔导度峰值(0.044 mol·m-2·s-1)出现在11:00 时;2 个品种间其气孔导度在13:00、15:00、17:00、19:00 和21:00 时测定的值均存在显著差异(P<0.05),而在其余时刻测定的气孔导度均无显著性差异(P>0.05)。
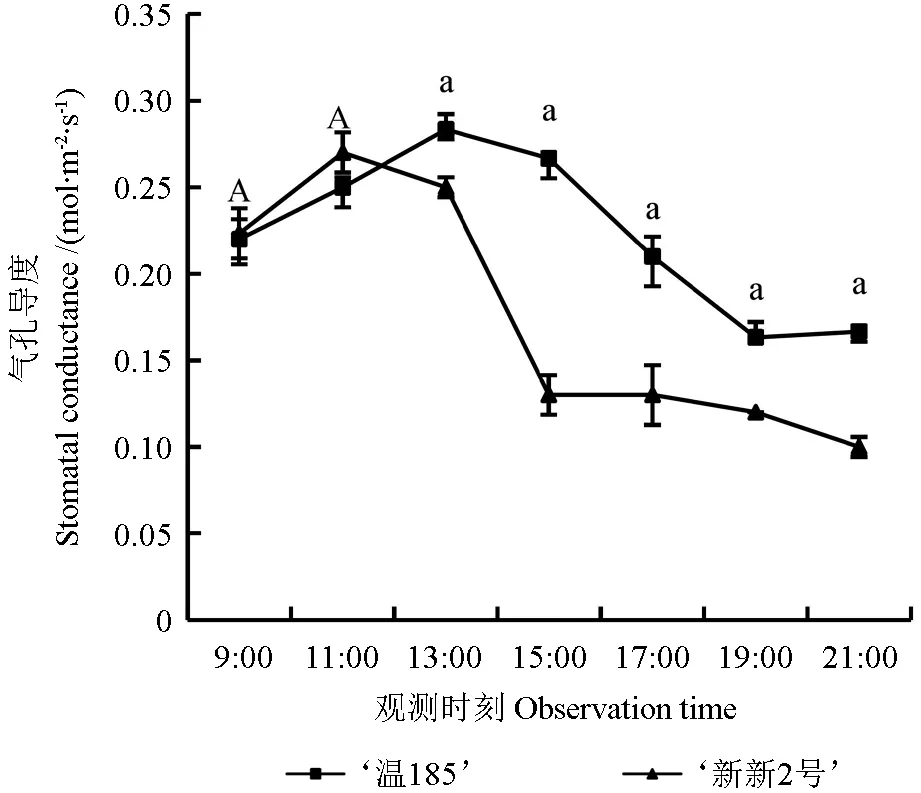
图7 不同核桃品种的气孔导度的日变化趋势Fig. 7 Diurnal variation trend of stomatal conductance of different walnut varieties
胞间CO2浓度是反映植株调控和影响光合作用的关键因子[24]。‘温185’和‘新新2 号’核桃品种的胞间CO2浓度(Ci)的日变化趋势如图8 所示。从图8 中可以看出,‘温185’的Ci日均值(261.06 μmol·mol-1)大于‘新新2 号’的Ci日均值(241.02 μmol·mol-1);‘温185’的胞间CO2浓度呈现出先下降再缓慢增加最后缓慢减小的变化趋势,‘新新2 号’的胞间CO2浓度则呈现出先快速下降再缓慢下降的变化趋势;2 个品种间其胞 间CO2浓 度 在13:00、15:00、17:00、19:00 和21:00 时测定的值均存在显著差异(P<0.05),而其余时间测定的胞间CO2浓度均无显著差异(P>0.05)。
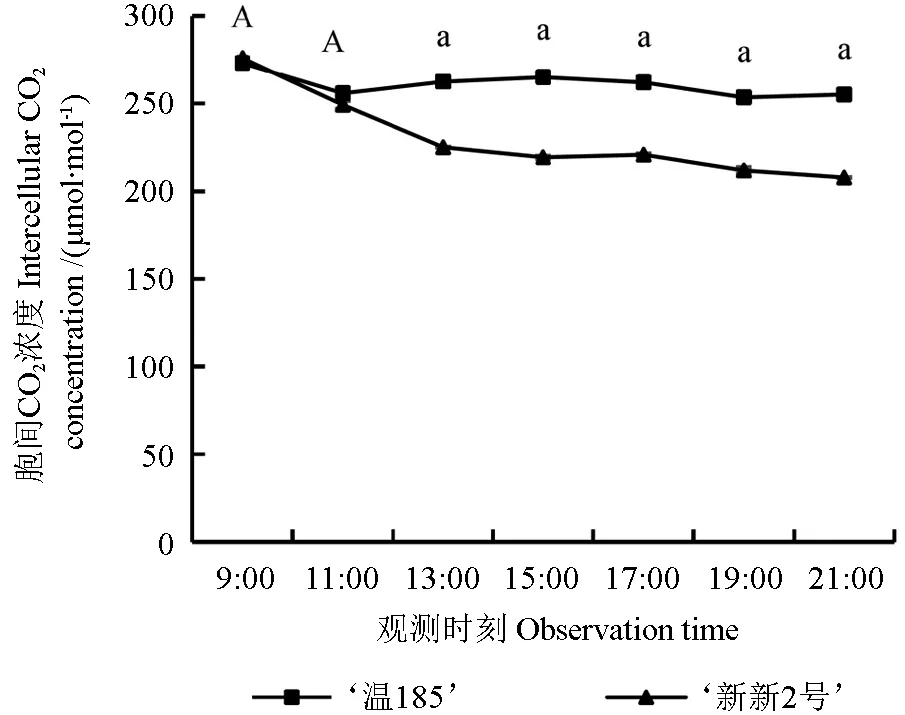
图8 不同核桃品种的胞间CO2浓度的日变化趋势Fig. 8 Diurnal variation trend of intercellular CO2 concentration in different walnut varieties
蒸腾速率是指植物通过蒸腾作用产生蒸腾拉力在单位时间内散失水蒸汽的量,是反映植物含水量的关键生理指标[25]。‘温185’和‘新新2 号’的蒸腾速率(Tr)日变化趋势如图9 所示。从图9 中可以看出,‘温185’和‘新新2 号’的蒸腾速率均呈现出先增加再减小的变化趋势,其日变化曲线类型均为单峰型;‘温185’的Tr日均值(0.007 7 mol·m-2·s-1)大于‘新新2 号’的Tr日均值(0.005 6 mol·m-2·s-1);‘温185’和‘新新2 号’的Tr峰值均出现在13:00 时,分别为0.015 0 和0.007 5 mol·m-2·s-1。‘温185’和‘新新2 号’间其蒸腾速率在13:00、15:00、17:00 和21:00 时测定的值均存在显著差异(P<0.05),而在其余时刻测定的值均无差异显著性(P>0.05)。
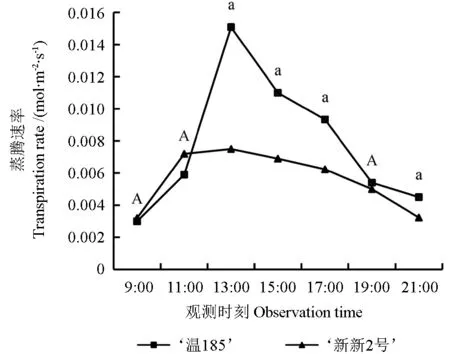
图9 不同核桃品种的蒸腾速率的日变化趋势Fig. 9 Diurnal variation trend of transpiration rate of different walnut varieties
2.2.2.3 叶片水分利用率的比较分析
叶片的水分利用率可以反映植物对水分的利用效果及对逆境的适应能力[26]。‘温185’和‘新新2 号’的水分利用率日变化趋势如图10 所示。从图10 中可以看出,‘温185’的水分利用率日均值(9.89 μmol·mol-1)大于‘新新2 号’的(8.68 μmol·mol-1);‘温185’和‘新新2 号’的水分利用率均在9:00 时达到最大值;9:00—11:00 时测定的水分利用率,‘温185’大于‘新新2 号’;13:00—17:00 时测定的水分利用率,‘新新2 号’大于‘温185’;17:00 时以后,‘新新2 号’的水分利用率随着时间的推移而缓慢增加,而‘温185’的水分利用率却呈现出先增加再减小的变化趋势。除了13:00 时测定的水分利用率2 个品种间存在显著差异(P<0.05)之外,其余时刻测定的水分利用率2 个品种间均不存在显著差异(P>0.05)。

图10 不同核桃品种的水分利用率的日变化趋势Fig. 10 Diurnal variation trend of water use efficiency of different walnut varieties
2.2.3 不同核桃品种的光合参数、SPAD 值与叶片显微结构指标之间的相关性分析
‘温185’与‘新新2 号’的净光合速率与各光合参数和SPAD 值之间的相关系数分别见表2和表3,‘温185’与‘新新2 号’的净光合速率与叶片各个显微结构指标之间的相关系数分别见表4 和表5。相关分析结果表明,‘温185’和‘新新2 号’的净光合速率与其蒸腾速率、气孔导度均呈极显著正相关;2 个品种的净光合速率与其光合有效辐射均呈显著正相关,而与其CO2浓度和叶片厚度均呈显著负相关。‘新新2 号’的净光合速率与胞间CO2浓度呈极显著正相关;而‘温185’的净光合速率与胞间CO2浓度无相关性。‘温185’的净光合速率与其栅栏组织和叶片组织疏松度均呈显著正相关,而‘新新2 号’的净光合速率与其栅栏组织和叶片组织疏松度均无相关性。
3 结 论
测定结果表明,核桃品种的叶长、叶宽和叶面积,‘温185’均略大于‘新新2 号’;‘新新2 号’的叶形指数略大于‘温185’的,其叶宽、叶形指数和叶面积的生长量差异均不显著。显微观察结果表明,‘温185’和‘新新2 号’的叶片均由上表皮、下表皮、栅栏组织和海绵组织构成;采用单因素检验法对‘温185’和‘新新2 号’叶片各个解剖结构指标的显著性进行分析,结果表明,叶片的上表皮厚度、下表皮厚度、栅栏组织厚度、叶片厚度、栅海比、叶片组织紧密度,‘温185’均大于‘新新2 号’;而叶片的海绵组织厚度,‘新新2 号’大于‘温185’。
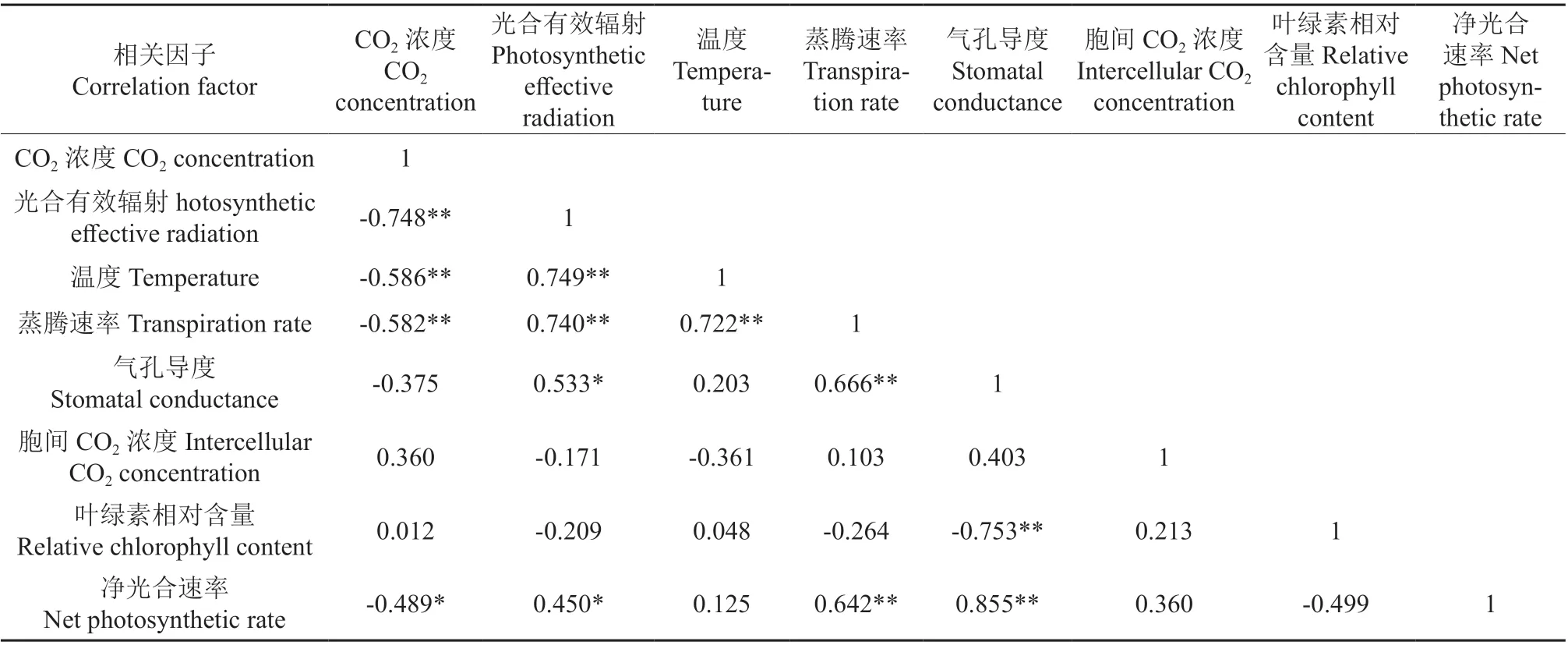
表2‘温185’的净光合速率与各光合参数和SPAD值之间的相关系数†Table 2 Correlation coefficient between net photosynthetic rate of ‘Wen 185’ and photosynthetic parameters and SPAD value
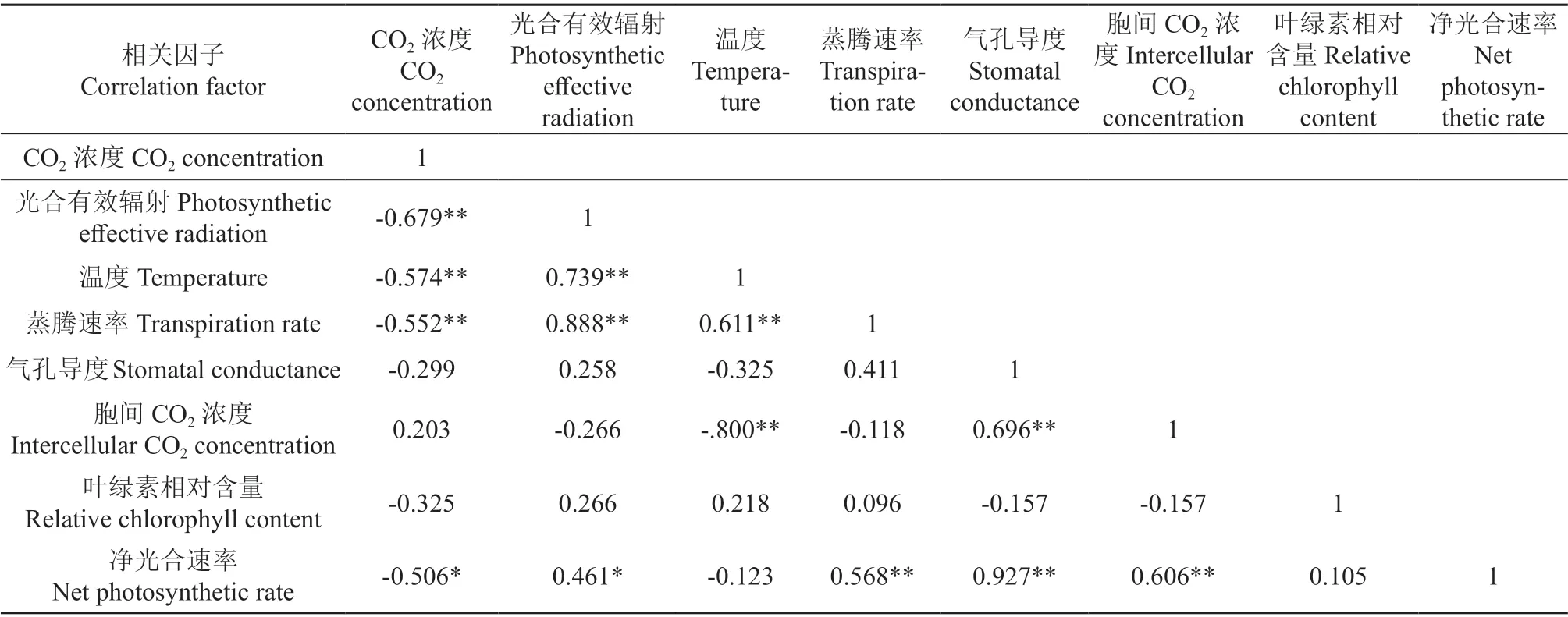
表3‘新新2号’的净光合速率与各光合参数和SPAD值之间的相关系数Table 3 Correlation coefficient between net photosynthetic rate and photosynthetic parameters and SPAD value of ‘Xinxin 2’
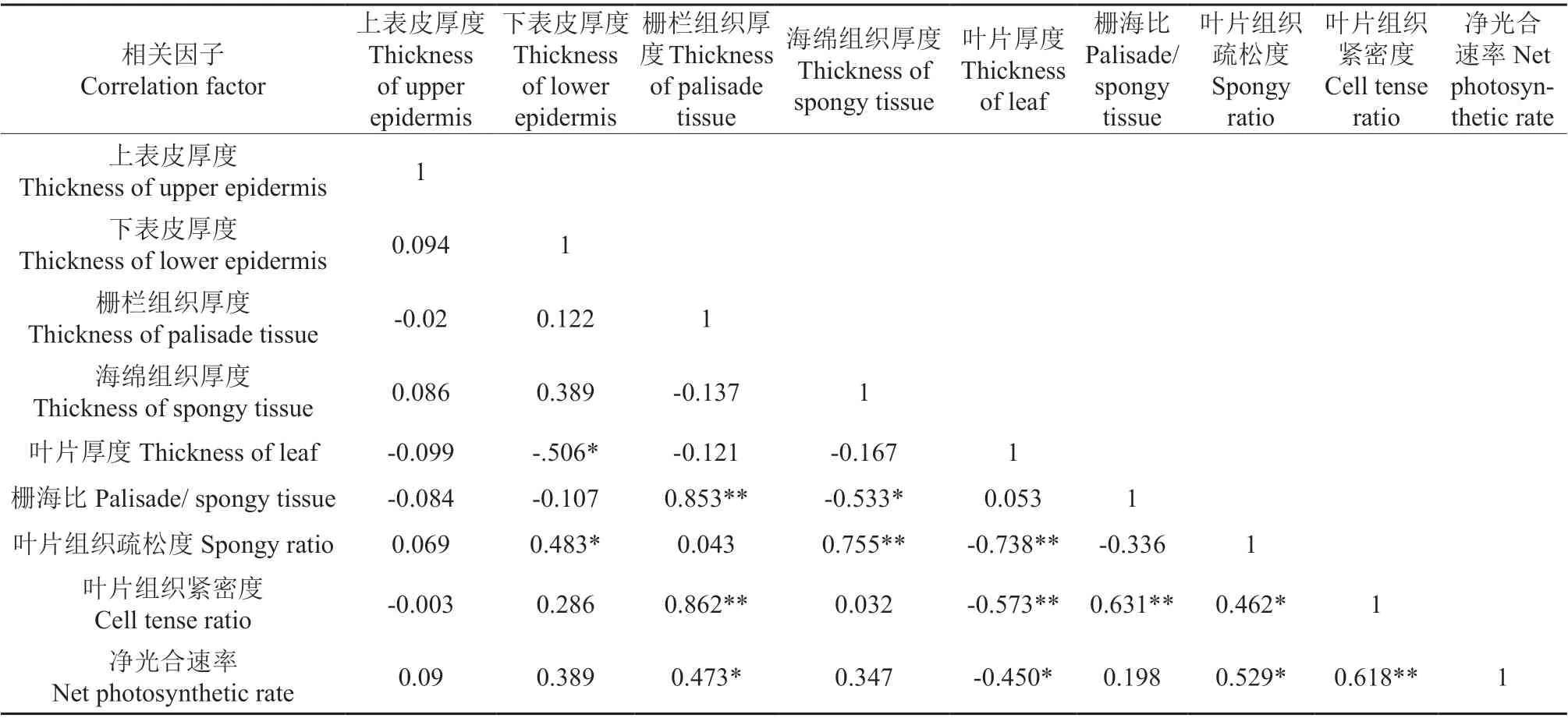
表4‘温185’的净光合速率与叶片各个显微结构指标之间的相关系数Table 4 Correlation coefficient between net photosynthetic rate of ‘Wen 185’ and various microstructure indexes of leaves
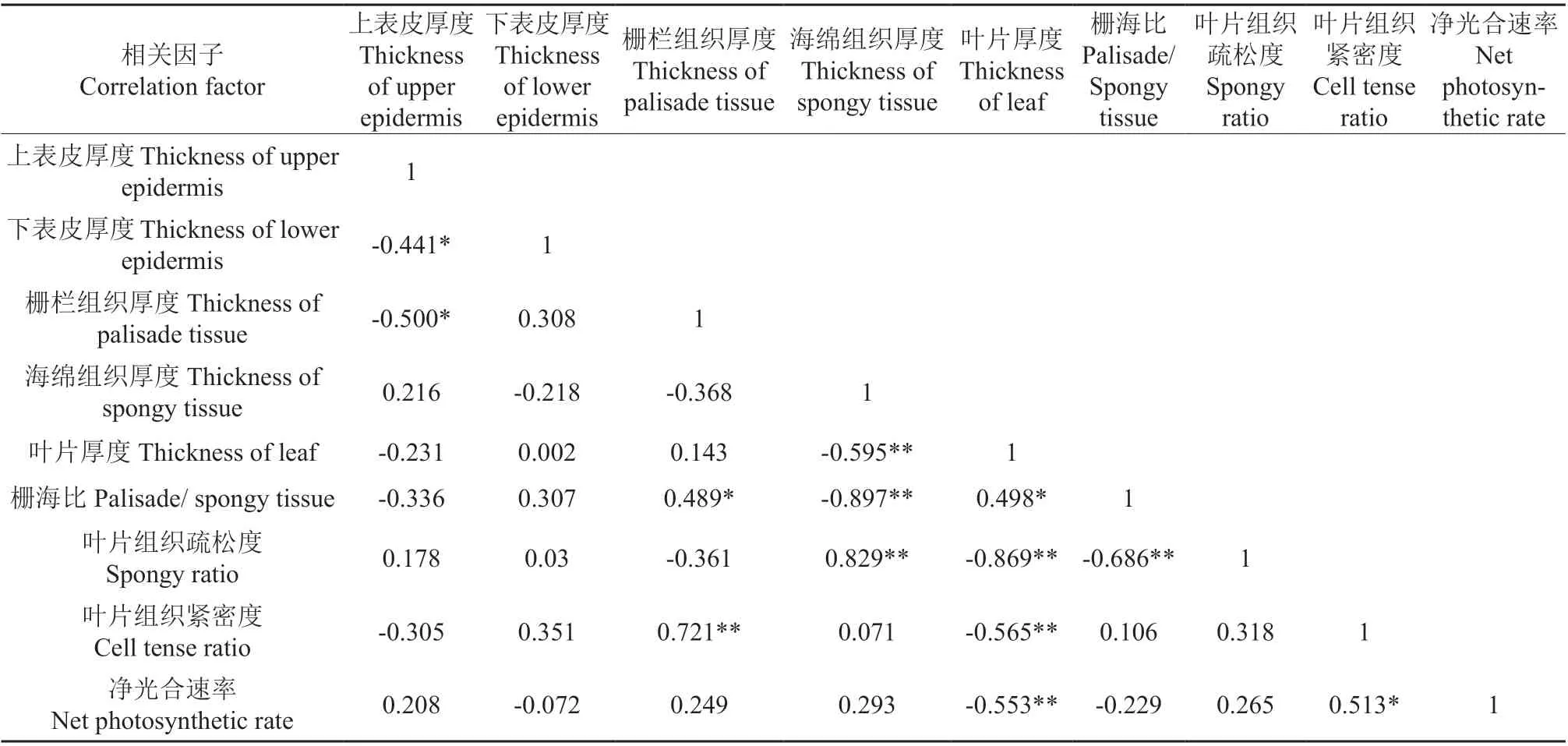
表5‘新新2号’的净光合速率与叶片各个显微结构指标之间的相关系数Table 5 Correlation coefficient between net photosynthetic rate and leaf microstructure indexes of ‘Xinxin 2’
‘温185’和‘新新2 号’所在环境各因子(光合有效辐射、温度和CO2浓度)的日变化趋势均基本一致;且‘温185’的净光合速率、气孔导度、胞间CO2浓度、蒸腾速率及水分利用率均大于‘新新2 号’的;且‘温185’和‘新新2 号’的净光合速率日变化曲线类型均为呈单峰型,表明2 个核桃品种都没有出现明显的光合午休现象。
2 个核桃品种的净光合速率与叶片栅栏组织厚度、叶片组织疏松度、蒸腾速率和气孔导度均呈正相关,说明核桃叶片的栅栏组织、叶片厚度、蒸腾速率、气孔导度及叶片的水分利用率都能影响其光合速率。此外,叶片栅栏组织的厚度、上表皮厚度、下表皮厚度和水分利用率均可作为植物抗旱性的评价指标。分析结果表明:‘温185’比‘新新2 号’更能适应南疆地区的气候条件。
4 讨 论
核桃树是喜光果树。‘温185’和‘新新2 号’作为新疆地区的主栽核桃品种,其树势均较强,树姿均较张开。植物叶片的微观构造是反映植物光合能力的结构基础[27],本研究通过分析核桃新梢叶片的生长势探究其光合特性。已有研究结果表明,叶片厚度、单位面积栅栏细胞数及表皮和表面角质层与植物的光合作用均显著相关[6-7]。研究中发现,‘温185’的栅栏组织、叶片厚度均大于‘新新2 号’的,且其净光合速率也大于‘新新2 号’的。相关性分析结果表明,‘温185’和‘新新2 号’的净光合速率与叶片厚度均呈负相关,而与叶片组织紧密度均呈正相关,反映出光合速率与叶片厚度和叶片组织紧密度均密切相关,‘温185’的净光合速率与叶片栅栏组织厚度和叶片组织疏松度均呈显著正相关,这与李艳丽等[28]对锥栗的研究结果一致。另外,叶片栅栏组织的厚度、上表皮厚度、下表皮厚度均可作为植物抗旱性的评价指标[29],并且栅栏组织的厚度与植物的抗旱性呈正相关[30]。研究结果表明,‘温185’叶片中栅栏组织的厚度大于‘新新2 号’的,说明‘温185’更能适应新疆地区的气候(典型的大陆性干旱气候)条件。
净光合速率是植物光合作用的直观体现,净光合速率的大小反映了植物同化物生产能力的大小[12]。有关研究结果表明,植物净光合速率的日变化曲线类型主要有双峰、单峰和不规则类型,‘温185’和‘新新2 号’的净光合速率日变化曲线类型均为单峰型,说明2个品种均未表现出明显的“午休”现象,这一结果与张志华等[31]的研究结果一致,而与前人对红富士[32]和樱桃[33]的研究结果均不一致。导致研究结果不一致的原因如下:一是品种不同;二是测定光合参数的仪器不同,本研究采用LI-6800 进行测定,而杨莹等[12]所用的光合仪器是LI-6400 XT 便携式光合仪;三是测定方法不同,张志华等[31]采用改良干重法测量光合参数,而本课题组成员则采用便携式光合仪测定叶片在某个时刻的光合参数,这样能更加准确地反映植物的光合特性。
环境因子(CO2浓度、光合有效辐射和温度)、气孔导度和蒸腾速率都是影响果树光合速率的重要因子[34]。研究中发现,随着温度和光合有效辐射的增加,核桃的净光合速率也有所增加,说明此时核桃树体开始生成机体所需的营养物质;核桃的净光合速率随着气孔导度和蒸腾速率的增加而增加,说明核桃的净光合速率与气孔导度和蒸腾速率均呈正相关,‘温185’和‘新新2 号’的净光合速率与蒸腾速率、气孔导度均呈正相关,这一分析结果与吴强等[27]对小黄杨的研究结论一致。同时,水分是植物进行光合作用的关键成分,水分利用率也是植物光合作用及抗旱性的关键评价因子。在相同的生境条件下,植株对水分的利用效率越大,说明植物在消耗等量水分的同时制造出的有机物越多,越利于植株在干旱逆境中生长[35],‘温185’的水分利用率日均值大于‘新新2 号’的,表明‘温185’的抗旱性强于‘新新2 号’的,‘温185’更能适应干旱条件。
本研究以‘温185’和‘新新2 号’为试验材料,通过测定其光合参数、观察其叶片显微结构、计算各个相关指标间的相关系数、参考前人研究结论来判断其抗旱能力,研究结果虽能反映出2 个核桃品种的抗旱能力,但还存在一定的不足,今后还可对核桃进行不同程度的干旱胁迫处理,并结合其表型、生理生化指标、叶片解剖结构及影响干旱的相关基因等指标,对其抗旱性进行综合评判;此外,果实是反映植株抗旱能力的直观体现,因此,还应在核桃果实不同的发育时期(果实速长期、果壳硬化期、种仁充实期和果实成熟期)对其进行干旱胁迫试验,并测定其光合参数与抗旱性指标,探究影响其干旱性的相关基因,并利用综合抗旱系数、抗旱指数和隶属函数综合评价值进行综合评价,这样才能更加科学、全面地评价不同核桃品种的抗旱性。
猜你喜欢
农业工程学报(2022年3期)2022-04-16
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
节水灌溉(2020年7期)2020-07-15
农业与技术(2020年12期)2020-07-04
中国农业科技导报(2020年12期)2020-03-15
农学学报(2019年1期)2019-02-21
新课程·下旬(2017年7期)2017-08-14
