
环境与植物激素对马铃薯块茎休眠与萌发的影响研究
2022-07-21 01:55李丽竹李俊花
中国种业 2022年7期
王 雪 王 谦 孙 源 李丽竹 李俊花 刘 浩 邓 纲
(云南大学资源植物研究院,昆明 650500)
马铃薯(Solanum tuberosumL.)是世界上最重要的粮食作物之一。因其特有的营养价值、经济价值、药用及保健价值和工业价值,对保障粮食生产安全和提高人民生活水平具有极其重要的作用。休眠是马铃薯块茎发育的特殊生理阶段,使块茎能够在不利条件下作为营养生殖器官保存,较长的休眠期有利于马铃薯的贮藏保鲜、加工利用,但同时也会造成马铃薯产量降低[1]。在马铃薯块茎的休眠期,芽的生长会受到明显抑制[2]。休眠时间的长短受到环境、生理和激素等影响[3]。而芽的萌发则标志着休眠期被打破,可用于相应的种薯生产等环节。如何做到人为控制马铃薯块茎休眠及芽萌发对马铃薯产业具有重要的经济价值,也是目前面临的一大挑战。
1 马铃薯块茎休眠及芽萌发的特征
马铃薯块茎达到生理成熟后,会进入很长一段时间的深度休眠状态。在此时种植马铃薯,即使是在理想的萌发条件下芽也不会萌发[4]。从生理学的角度来说,马铃薯块茎休眠期是指从匍匐茎顶端膨大开始到块茎发芽生长的一段时期,但人们习惯将块茎开始收获至块茎顶芽萌发结束的时期作为马铃薯块茎的休眠期[3]。处于休眠过程中的块茎仍保持着生命活动,维持着最基础的生理功能,只有经过一定的贮藏后休眠才会被解除,块茎才能进一步发芽生长[5]。休眠时期一般被划分为相对休眠、生理休眠和环境休眠。因各阶段间不存在明显的生理标志而导致休眠期不易被区分。对休眠时期的划分一般根据休眠是否可以被逆转来判断,相对休眠和环境休眠可受外界因子的影响被打破,而生理休眠则不会或很难被逆转[6]。
2 外界环境对马铃薯块茎休眠与萌发的影响
休眠时期的长短主要与品种的特性有关,同时也会受到环境、栽培措施和贮藏条件等因素的影响[3]。不同品种马铃薯的休眠期存在一定的差异,晚熟品种一般具有较短的休眠期,而早熟品种则相反。除此之外,休眠期时间还取决于马铃薯生长的土壤和气候等条件、块茎成熟度、贮藏条件以及块茎的完整度等。一般来说,栽培过程中适宜温度(15~21℃)范围内的高温、足够发育的水分和良好的土壤肥力会缩短块茎的生理休眠期;未成熟比成熟的块茎有着更长的休眠期[3];在低温、潮湿条件下贮藏则有利于延长块茎的休眠期[4]。
2.1 温度对马铃薯块茎休眠和萌发的影响一系列研究表明,马铃薯在收获后,块茎的休眠时间与贮藏温度有关。通常情况下,在持续高温条件下,休眠期会明显地缩短[4]。而随着学者们不断深入地研究,进一步发现不同的温度处理对马铃薯休眠期有显著的影响。Wurr 等[7]通过分析3 个马铃薯品种(Desiree、Vanessa、Home Guard)在不同贮藏温度(2~8℃、0~10℃、6~15℃)条件下休眠期的表现发现,马铃薯块茎在2~8℃条件下连续贮藏14d,块茎的萌发会受到明显抑制,而恢复到6~15℃时,休眠被打破,与对照组相比,休眠期缩短14d;而一直在10℃条件下贮藏的块茎则没有芽生长现象,同时揭示了经冷处理、常规温度条件下贮藏后,发芽的数量与品种密切相关。Van Ittersum 等[8]对不同品种(Desiree、Diamant、Draga、Jaerla)的马铃薯进行热预处理,即在28℃条件下连续贮藏20d 后恢复到18℃条件贮藏,与持续贮藏在18℃条件下相比,经热预处理的马铃薯块茎休眠期分别缩短了14d、29d、18d 和15d;进一步分析发现,持续贮藏在28℃条件下也可以缩短马铃薯块茎的休眠期,分别缩短了49d、13d、48d 和51d,与热预处理相比,Desiree、Draga、Jaerla 品种块茎的休眠期明显缩短。
2.2 湿度与光照对马铃薯块茎休眠和萌发的影响湿度在一定程度上会影响马铃薯块茎的休眠,关于湿度与马铃薯块茎休眠间的研究相对较少,主要是因为环境范围内湿度的变化对马铃薯块茎水分的影响不显著。然而,这种影响对块茎质量有重要的影响,尤其是新鲜的马铃薯[9]。同时高湿度有利于马铃薯块茎伤口的快速愈合,从而影响马铃薯的完整度,促进块茎芽的萌发,有利于打破休眠;相对湿度处于90%以下,可防止块茎重量下降过多;相对湿度处于38%~66%之间,块茎则开始发芽[10]。
有研究表明,短日照条件栽培的马铃薯比长日照条件栽培的马铃薯具有更短的休眠期[11]。在田间条件下,光周期对马铃薯块茎休眠的影响很难确定,所观察到的影响可能更多的与块茎形成时间或成熟度有关,而与休眠本身无关,通过对马铃薯(Desiree)处于休眠状态下的块茎进行不同的光照处理发现,光强对块茎休眠具有一定的影响。短日照条件下,与较低光强(5.3μmol/m2·s PPF)的处理相比,最高光强(106μmol/m2·s PPF)的处理则可以显著缩短马铃薯块茎的休眠期。同时发现块茎休眠期与光周期有关,与完全暴露在光照8h/d 相比,完全黑暗处理条件下的马铃薯块茎休眠期明显缩短[6]。
2.3 大气条件对马铃薯块茎休眠和萌发的影响块茎休眠的打破和芽的萌发过程中伴随着高浓度的CO2和O2等条件,但两者之间具体的生理机制尚不明确[9]。Burton[12]研究发现马铃薯块茎处于1%~2%的CO2含量下有利于打破休眠。后续的研究表明,随着块茎的老化,打破休眠和维持芽生长所需的O2含量会显著增加(从2%增加到23%),同时提出了“最适厌氧可逆代谢”的生长抑制机制。Thornton[13]研究表明,马铃薯块茎在25℃条件下连续施用40%~60%的CO2和20%的O23~7d,可有效打破休眠,同时还发现高浓度(20%~80%)的O2能增强这种效果。Coleman 等[14]发现对马铃薯块茎进行特定的CO2/O2气体混合物处理可缩短休眠期。其中有两种处理对所有马铃薯品种均适用,分别为低O2含量(<10%)/10%~60% CO2含量以及高CO2含量(60%)/O2含量(40%)。而最有效的方法则是对马铃薯的块茎进行20% CO2与40% O2或60%CO2与18%~20% O2处理,在此基础上引入乙烯可以进一步缩短马铃薯块茎的休眠期。
3 植物激素对马铃薯块茎休眠和萌发的影响
植物激素是影响马铃薯块茎休眠和芽萌发的重要因素,目前主要通过使用植物激素对块茎进行化学处理来提前解除休眠,但具体作用机制仍需进一步研究。前人研究表明,只有当一些特定的激素处于相对平衡状态,才可以促进或维持某一生理状态。在马铃薯块茎休眠过程中,植物激素会有明显的变化(表1),块茎休眠主要受生长促进剂和抑制剂的相对浓度调控,并提出了“抑制因子/促进因子平衡调节休眠”的学说来解释[3]。
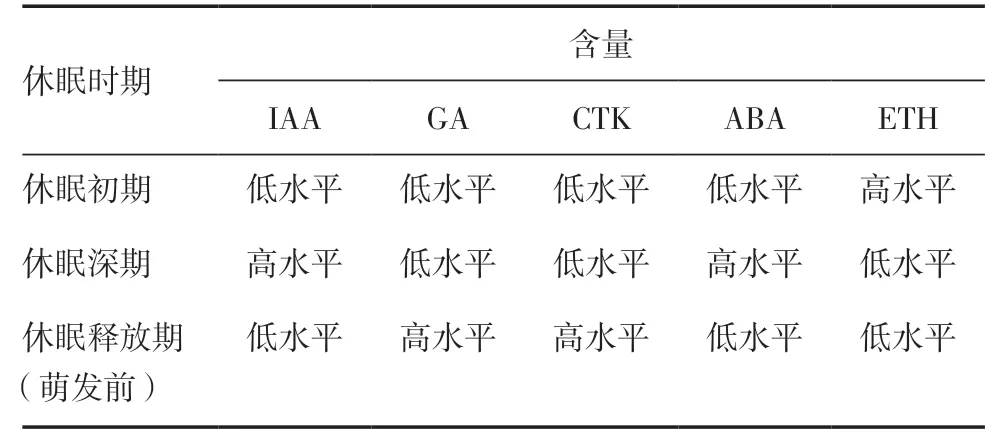
表1 马铃薯块茎休眠过程中主要内源激素的含量变化
3.1 生长素对马铃薯块茎休眠和萌发的影响植物生长素(IAA)是影响马铃薯块茎休眠的主要激素之一。马铃薯块茎中内源IAA 的含量会随着休眠程度的不同而产生相应的变化。在块茎形成过程的初期,内源IAA 含量升高;随着块茎进一步发育,IAA 含量会持续下降[15]。目前,IAA 控制马铃薯块茎休眠和芽萌发的作用机制尚不明确。Hemberg[16]对马铃薯块茎进行生物测定发现,块茎休眠期内源IAA 含量较低,伴随着早期芽的生长IAA 含量明显增加。Sukhova 等[17]进一步发现,马铃薯块茎中自由态IAA 含量从休眠初期到休眠结束后没有显著变化,但总的IAA 含量在减少,推测是结合态IAA 含量降低所导致。Sorce 等[18]分析了马铃薯块茎芽萌发前IAA 的含量,发现萌发芽中自由态的IAA 含量显著增加,进一步通过免疫组织化学方法分析块茎组织和芽中IAA 分布发现,生长素有利于打破休眠,在发芽初期可促进芽的分化和生长。同时也发现高浓度IAA 或合成的生长素对芽萌发有一定的抑制作用,通过施用不同IAA 浓度梯度观察芽的萌发情况,发现高浓度IAA(1000mg/L)完全抑制芽的萌发,100mg/L IAA 处理下抑制不明显,而低浓度IAA(<10mg/L)可刺激芽的萌发,且当IAA 浓度为1mg/L时刺激芽萌发的效果最佳[16]。但也有研究表明,IAA 会抑制马铃薯块茎腋芽的发生以及匍匐茎的伸长,IAA 在抑制匍匐茎伸长的同时会促进无柄块茎的形成和块茎的膨大[19]。但IAA 浓度高于5μmol/L时,会抑制块茎的形成,这可能是高浓度IAA 含量影响了乙烯(ETH)的形成[20]。综上所述,内源IAA控制马铃薯块茎休眠的假设并没有得到有力证据的支撑,却证实了IAA 和其他内源性生长素在马铃薯解除休眠后芽生长萌发过程中发挥作用。因此,一定浓度范围的IAA 是芽生长所必需的,但不是芽生长萌发的启动因子。
3.2 赤霉素对马铃薯块茎休眠和萌发的影响赤霉素(GA)是一类可以促进马铃薯块茎打破休眠及芽生长萌发的植物激素,在马铃薯块茎的生长发育过程中,GA 通过细胞分裂及细胞骨架、光周期诱导块茎形成时信号的传递和碳水化合物的合成等过程来调控马铃薯块茎的形成和发育[21]。
GA 对马铃薯块茎休眠的打破具有重要作用。低浓度GA 可加速休眠打破和块茎芽的萌发进程[4],而短日照条件下GA(浓度≥100mL/L)对马铃薯块茎的形成具有抑制或延迟块茎形成的作用[22]。Suttle[23]通过研究马铃薯块茎中GA1、GA19和GA20的含量发现,块茎中GA19和GA20的含量在发芽旺盛时期最高,且此时GA 总含量增加。同时有研究表明发现,GA19对非休眠期块茎的发芽生长有一定的促进作用。以上结果表明,内源性GA对块茎休眠的打破与芽的生长萌发具有一定的调控作用。Xu 等[24]通过对诱导条件(8%蔗糖+GA 0.01~0.10μmol/L)和非诱导条件(1%蔗糖;8%蔗糖+GA4/7)下马铃薯块茎或匍匐茎中GA 的含量进行比较发现,在匍匐茎膨大形成块茎的过程中,两者GA 的含量均显著下降,在块茎中维持较低水平。进一步分析发现,非诱导条件(1%蔗糖)下GA 的含量较诱导条件(8%蔗糖)下高,说明马铃薯块茎的形成需要GA 的含量低于一个临界水平(1μmol/L)。内源GA 类似化合物在块茎休眠期含量较低而在芽生长前增加,表明用GA 处理休眠期的块茎可打破其休眠状态[25]。尽管GA 在马铃薯块茎休眠调控中的作用一直存在争议,但通过外源施用、生物合成基因的异位表达以及人工增加块茎GA 含量等手段都能够促进块茎打破休眠。
3.3 细胞分裂素对马铃薯块茎休眠和萌发的影响细胞分裂素(CTK)是马铃薯块茎休眠及芽生长萌发过程中的调节因子,能够刺激植物细胞分裂、增强植物组织的吸收能力,进而促进块茎打破休眠和芽的生长萌发[26]。对CTK 活性和含量的相关研究表明,当块茎处于深度休眠期时,CTK 含量较低,而在发芽前和出苗期前含量会增加且达到最高值。块茎休眠解除后,块茎和芽中CTK 总含量进一步增加[17]。目前为止,细胞分裂素调控块茎休眠和发芽的作用机制仍不清楚。
细胞分裂素存在于马铃薯块茎的所有部位,在休眠时期,细胞分裂素均匀分布在顶端、外侧和节间组织。在块茎打破休眠的过程中,CTK 能够刺激块茎进行细胞分裂,解除细胞周期G1/S 期的阻滞,最终导致休眠被打破,相关的研究表明,D 型细胞分裂素参与细胞周期G1/S 期的阻滞的去除[27],一定程度上印证了CTK 对块茎的解除休眠作用。据报道,块茎休眠的打破往往伴随着根尖芽以及相邻组织中游离细胞分裂素水平的迅速增加,且细胞分裂素在根尖组织顶端优势存在的情况下就会维持高水平的表达;而当顶端优势被克服,侧芽及邻近组织的细胞分裂素水平与根尖区域的细胞分裂素表达水平相当,表明细胞分裂素苷被运输到块茎的分生区,并被水解成自由碱基,在块茎的分生区和芽中明显地转化为贮藏形式(特别是玉米素糖苷)[28]。研究证明,马铃薯块茎中同时含有反式玉米素家族的细胞分裂素和顺式玉米素衍生物,外源细胞分裂素(顺式和反式玉米素)不仅可以有效地打破块茎休眠,还能引起内源游离顺式玉米素含量的增加,而顺式玉米素被认为是马铃薯块茎休眠的潜在调控因子[29]。此外,马铃薯块茎对细胞分裂素的敏感性随着休眠程度的进一步加深而逐渐增加。尽管收获后或收获后早期的马铃薯块茎对外施的细胞分裂素不敏感,但其内源性细胞分裂素含量会随着细胞分裂素的外施而增加[27],认为是外源性的细胞分裂素被休眠的块茎迅速代谢从而导致休眠被打破[30]。此外,休眠块茎对细胞分裂素表达水平随着时间的推移而不断下降,同时细胞分裂素代谢水平并没有明显的变化,表明细胞分裂素的感知、信号传导途径等受休眠状态的影响[29]。这些研究结果进一步表明,内源性细胞分裂素是马铃薯块茎中天然的终止休眠因子。
3.4 乙烯对马铃薯块茎休眠和萌发的影响植物激素乙烯(ETH)是一种有效的生长调节剂,能够影响植物的生长、分化和衰老等进程[31]。一系列关于乙烯与马铃薯块茎发芽间的研究表明,乙烯有利于块茎休眠的启动和维持,因此有必要对乙烯影响休眠的作用机制进行相应的研究和阐述。研究发现,乙烯在维持休眠的第1 阶段表现最为活跃,内源乙烯的含量在离体培养的块茎进入休眠初期时达到最高,随后迅速下降。用乙烯拮抗剂、硝酸银和降冰片二烯等可以使块茎休眠被打破,表现为块茎提前发芽,而这个过程可以被乙烯处理逆转,且在乙烯的作用下,休眠初期具有一定的稳定性[6]。与施用乙烯相比,休眠块茎经乙烯发生剂2-氯乙基膦酸处理后,不仅提高了组织中乙烯的含量,而且促进了ABA 的生物合成,最终表现为马铃薯块茎休眠期的延长。因此,乙烯被认为对块茎休眠有调控作用[3]。但也有学者提出不同的观点,Suttle[32]利用乙烯合成抑制剂氨基乙氧基乙烯基甘氨酸(AVG)和NG 同时处理微型薯时,发现乙烯含量没有增加,但微型薯提前解除休眠,而外源乙烯或者乙烯合成激活剂氨基环丙烷羧酸(ACC)处理的微型薯并没有提前解除休眠,认为内源乙烯没有参与诱导块茎休眠解除。Rylska 等[33]用不同浓度的乙烯处理马铃薯块茎,发现休眠块茎在浓度为2μL/L 的乙烯处理72h,块茎发芽的效果最好,且环氧乙烷的处理效果比乙烯更好。用乙烯处理马铃薯休眠块茎超过72h,则会抑制块茎发芽;对于已解除休眠期的块茎,用乙烯长期处理,也会抑制芽的伸长。但在马铃薯块茎中检测到的乙烯浓度很低(小于0.1μL/L),认为内源乙烯并不是芽的抑制剂。Prange 等[34]用乙烯持续处理商品化贮藏的块茎,通过分析马铃薯块茎中还原糖的含量发现,虽然马铃薯块茎中还原糖含量有所增加,但芽的生长萌发仍被抑制;且内源乙烯的表达水平只在发芽期才略有增加,在此之前乙烯的含量达不到萌芽的条件,因此乙烯不能作为芽的内源抑制剂。
3.5 脱落酸对马铃薯块茎休眠和萌发的影响脱落酸(ABA)是导致和维持块茎休眠的主要植物激素之一。ABA 首次在马铃薯块茎中β-抑制性复合物中被检测,发现能够抑制块茎茎尖生长,在块茎休眠时含量最高,随着休眠的解除,茎尖开始生长时,含量迅速下降,被认为可以调控块茎的休眠[35]。杨柏云等[1]研究发现,芽眼和块茎组织中ABA 含量在休眠开始时均增加,在深度休眠期最高,在休眠结束时急剧下降,块茎中ABA 含量在打破休眠后也随之降低,表明ABA 可以调控马铃薯块茎的休眠。
研究证实外源ABA 会对马铃薯块茎芽的生长产生暂时性、剂量依赖性的抑制作用[31]。Suttle 等[36]通过系统研究了内源ABA 以及ABA 生物合成抑制剂(氟醚酮)在块茎休眠中的作用后发现,马铃薯块茎经氟醚酮处理后ABA 的合成被有效抑制,表现为块茎提前发芽。而外源施用ABA 则使块茎中ABA 恢复到对照水平,相应地观察到早熟芽被抑制的现象。此外,对休眠的马铃薯块茎外源施用ABA 抑制剂也会观察到ABA 含量下降以及早芽率的增加。综上所述,马铃薯块茎休眠的诱导和维持都需要ABA 的持续生物合成。近年来,块茎在生理休眠期间ABA 含量的变化与某些ABA 生物合成和分解基因之间的时间相关性已有报道,然而由于长时间的自然休眠过程使得阐述这些现象变得复杂。
4 展望
如何做到人为调控马铃薯块茎的休眠期以及芽的萌发,从而发挥马铃薯最大的经济价值是国内外学者所面临的重大科学问题。在马铃薯块茎休眠的调控中,外界环境因素和内源植物激素的特性和作用已经取得了一部分的进展。然而,关于植物激素在马铃薯块茎生理和休眠等过程中的调控机制,还需要进一步的研究。目前通过控制光照、温度、湿度等外界环境因素虽然可以控制马铃薯块茎的休眠期,一定程度上提高马铃薯的产量和品质。在此基础上,对内源植物激素间互相调控机制的阐述必将使得马铃薯产业得到进一步的发展。目前通过基因工程已确定马铃薯休眠过程中一些与调控激素相关的基因和蛋白质等物质,为更好地解释块茎休眠和芽萌发调控机制奠定了基础。
已知的内源植物激素间的调控及互作机制仍需要进一步探索,而其他内源植物激素在块茎休眠过程中的作用也在不断地被认知,如油菜素内酯等。为解决马铃薯休眠机制阐述这个问题,深入了解各激素特别是赤霉素和乙烯在块茎休眠和萌发中的作用机制尤为必要;而各植物内源激素间的平衡及相互调控机制以及其他内源激素在休眠和萌发中产生的影响和调控途径的相关研究则可以进一步加速对马铃薯块茎休眠机制的认识。最后,可利用基因工程等对马铃薯休眠机制进行更深层次的阐述。相信在不久的将来,随着马铃薯休眠及芽萌发机制的不断清晰,马铃薯的经济价值可以得到充分的利用。
猜你喜欢
农业工程学报(2022年8期)2022-08-08
作物学报(2022年9期)2022-07-18
农村农业农民·B版(2018年3期)2018-04-09
新农村(2017年14期)2017-09-30
考试周刊(2016年82期)2016-11-01
中学教学参考·理科版(2016年2期)2016-04-05
安徽理工大学学报·社会科学版(2014年6期)2015-02-06
银行家(2014年1期)2014-02-14
现代农业研究(2009年3期)2009-04-14
