
高、低温胁迫对草地贪夜蛾生长发育及抗寒性的影响
2022-07-25 08:10母银林黄秀东王玉川喻会平陈祥盛
山地农业生物学报 2022年4期
汪 洁,母银林,杨 灿,黄秀东,王玉川,喻会平,陈祥盛*
(1.贵州大学 贵州山地农业病虫害省级重点实验室,贵州 贵阳 550025;2.贵州大学 昆虫研究所,贵州 贵阳 550025;3.贵州大学 贵州省昆虫资源开发利用特色重点实验室,贵州 贵阳 550025;4.贵州省烟草公司 毕节市公司技术中心,贵州 毕节 551700)
草地贪夜蛾隶属于鳞翅目Lepidoptera夜蛾科Noctuidae,是原产于美洲热带和亚热带的一种重要农业害虫。自2019年1月11日发现草地贪夜蛾侵入我国云南省江城县,同年10月已扩散至全国26个省(区、市)的1518个县发生为害,足见其扩散速度快和适应力强。草地贪夜蛾的食性广,可取食76科353种植物,尤喜食玉米、稻米、燕麦、高粱、小米、苜蓿、甘蔗、蔬菜和棉花等80多种作物,能导致粮食严重减产。草地贪夜蛾幼虫共六龄,各龄均可造成为害,其中1~3龄幼虫群集为害,取食植物叶片叶肉造成“窗孔”状透明薄膜,严重时叶片出现孔洞、缺刻等,4~6龄幼虫可取食玉米心叶、叶鞘、穗轴及籽粒等。
昆虫对温度胁迫的响应和耐受性可间接反映出种群动态、发生规律以及分布和扩散能力的信息。对短期、突发性刺激产生的反应通常要比长期、固定性因素更加强烈。如禾谷缢管蚜受38 ℃短时高温处理后,当代成蚜寿命显著缩短,产蚜量降低,且禾谷缢管蚜F代种群增长明显受到抑制;较短时间的低温驯化可有效提高广聚萤叶甲成虫在致死低温下的存活率;苹淡褐卷蛾幼虫经冷驯化处理后,过冷却点和死亡率均明显降低;短时高、低温处理对斜纹夜蛾的生长发育和存活会产生不利影响,但一定程度上可提高其耐寒性。目前,有关草地贪夜蛾对温度响应的研究已有报道。Schlemmer报道其最佳发育温度为26~32 ℃,最佳产卵温度在18~26 ℃。当温度为35 ℃时,草地贪夜蛾卵孵化率和蛹存活率均较低,且成虫不能产卵;15 ℃下的低温环境不能正常繁殖,表现出生长速率下降和死亡率上升。罗举等研究表明,草地贪夜蛾3龄幼虫过冷却点为-7.65 ℃;张红梅等报道草地贪夜蛾卵和蛹经短时高温处理后对其生长发育的影响显著。
草地贪夜蛾世代周期较短,一年内可发生多代,研究表明,短时温度变化可对世代周期短的昆虫生长发育、地理分布及生殖等产生影响。因此,研究短时高、低温对草地贪夜蛾选择入侵地及种群发生有重要的参考意义。由于草地贪夜蛾3龄幼虫开始进入暴食阶段且抗药性显著增强,故本试验选择3龄幼虫分别在高温36 ℃、低温13 ℃下进行短时胁迫,观察不同处理对其后期生长发育、死亡率及耐寒性的影响。以期为极端温度频发下草地贪夜蛾的种群发生规律、预测预报及防治提供参考依据。
1 材料与方法
1.1 材料
..供试虫源
草地贪夜蛾幼虫于2019年月6月采自贵州省六盘水市盘州市鸡场坪镇滑石乡(N 25°54′,E 104°36′,平均海拔1800 m)的玉米地,在人工气候箱中用新鲜玉米苗饲养,控制温度(27±1) ℃、相对湿度(65±5)%、光周期L∶D=16 h∶8 h,光照强度5000 lx,提供灭菌蛭石供其化蛹,成虫置于养虫笼中,用10%蜂蜜水喂养。供试虫源为室内继代饲养的第7代3龄幼虫。
..主要仪器
试验使用的主要食品为RGL-P系列人工气候培养箱(0~50 ℃)(合肥达斯卡特生物科技有限公司)、METTLER TOLEDO万分之一天平(梅特勒利托多上海有限公司)、低温冰柜(Haier biomedical DW-40W100)。
1.2 方法
..高、低温处理对幼虫生长发育的影响
取大小一致的3龄幼虫,置于体积750 mL的养虫盒内,每盒10头,每个处理10头试虫,重复9次。在36 ℃和13 ℃人工气候箱中分别处理8、16、24、32 h后于正常饲养条件下继续饲养,以27 ℃下正常饲养的为对照(下同)。4龄后为了避免自相残杀,转移至体积25 mL的透明塑料杯中单头饲养,处理期间进入4龄的也及时转移到相同规格塑料杯中继续处理。每天8∶00和20∶00分别观察1次并根据情况更换食物,记录幼虫各龄期蜕皮、发育与死亡情况,直至幼虫全部化蛹。
..高、低温处理对预蛹和蛹的影响
幼虫进入预蛹后取出放入300 mL装有灭菌蛭石的养虫杯中,与幼虫期观察时间相同,观察记录预蛹期和化蛹率,待化蛹颜色完全呈棕红色后用电子天平称量蛹重(单头)。
..高、低温处理对幼虫耐寒性的影响
试虫与..处理相同,经高、低温胁迫后,选取存活试验试虫及对照试虫均置于-7 ℃冰柜中1 h,然后转入正常饲养条件下饲养24 h后观察统计存活情况。每处理10头试虫,重复9次。
成虫期:指初羽化的成虫一直到其死亡的时期;
死亡率(%)=(此发育阶段死亡的虫量/进入该阶段的虫量)×100%;
化蛹率(%)=(蛹数/老熟幼虫数量)×100%。
1.3 数据分析
数据采用SPSS 26软件先进行正态性检验和方差性检验,对通过正态性检验和方差齐性检验的数据进行单因素方差分析,用Duncan氏新复极差法进行多重比较,包括各虫态的发育历期、化蛹率、蛹重和抗寒性;各虫态死亡率数据不符合正态分布,采用Kruskal-Wallis法进行差异性检验,如存在差异则采用Bonferroni校正法进行多重比较,<0.05有统计学意义。利用Sgmaplot 14和GraphPad Prism 8作图,并用回归分析拟合生长发育与其处理时间之间的关系。
2 结果与分析
2.1 高、低温处理对草地贪夜蛾发育历期的影响
..高温对各龄幼虫发育历期的影响
36 ℃高温处理8 h后的3龄幼虫发育历期与对照组差异不显著(图1),其余处理组的3龄幼虫发育历期均极显著延长(=12.035,<0.01)。处理24 h的3龄幼虫,其4龄幼虫发育历期与对照组差异不显著,处理8 h、16 h和32 h的发育历期分别显著延长至3.242 d、3.084 d、3.126 d(=3.533,=0.015)。3龄幼虫经高温处理8 h后,其5龄幼虫发育历期与对照组差异不显著,分别为1.433 d和1.408 d,其余的处理组比对照组显著延长(=68.010,<0.01),3龄幼虫高温处理时长与5龄幼虫发育历期呈显著线性关系(=0.271+1.272,=0.817,=191.906,<0.01)。3龄幼虫经高温处理8 h和16 h后,其6龄发育历期分别显著延长至4.060 d和3.733 d(=4.108,=0.007),处理24 h和32 h后与对照组的差异不显著。高温处理极显著延长了3~6龄幼虫累计发育历期,处理32 h的延长至13.496 d (=45.601,<0.01),二者呈显著线性关系(=0.719+10.458,=0.807,=180.257,<0.01)。高温处理不同时长后的预蛹期均极显著缩短(=107.511,<0.01),二者呈显著线性关系(=0.258+2.125,=0.861,=265.689,<0.01)。3龄幼虫经过高温处理8 h后蛹期显著缩短为8.428 d,处理32 h的显著延长为9.313 d (=12.622,<0.01)。高温处理后成虫期差异不显著(=12.622,=0.647)。表明36 ℃高温处理的3龄幼虫在随后的发育过程中,3~6龄幼虫累计发育历期及预蛹期所受影响较大,其次是5龄幼虫的发育历期,成虫期受影响较小。
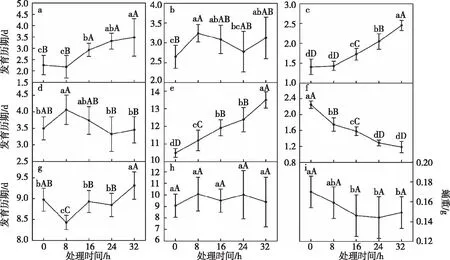
注:a~i依次为3龄幼虫、 4龄幼虫、5龄幼虫、6龄幼虫、3~6龄幼虫、 预蛹、蛹、成虫、蛹重。图中数据为平均数±标准差。不同大、小字母表示同一龄期不同时长处理间经Duncan氏新复极差法检验在P<0.01和P<0.05水平差异显著。图1 经36 ℃高温处理后草地贪夜蛾各龄幼虫的发育历期及蛹重Fig.1 Effects of high temperature 36 ℃ on larval duration of 3rd~6th instars and pupal weight of S. frugiperda
..低温对各龄幼虫发育历期的影响
3龄幼虫经13 ℃低温不同时长处理后,随后各龄及3~6龄幼虫累计发育历期整体呈延长趋势(图2)。其中处理8 h后3龄幼虫发育历期与对照组无显著差异,其余处理的3龄幼虫发育历期均显著长于对照组,其中处理32 h的延长至3.483 d,是对照组的1.543倍(=34.362,<0.01)。经低温处理8 h、16 h及24 h后4龄幼虫发育历期与对照组差异不显著,处理32 h的显著长于对照组(=3.330,=0.019)。低温处理后5龄幼虫发育历期均显著长于对照组(=108.116,<0.01),二者呈显著线性关系(=0.280+1.592,=0.814,=187.988,<0.01)。3龄幼虫经低温处理8 h后,其6龄幼虫发育历期与对照组无显著差异,处理16、24和32 h的显著长于对照组(=4.370,=0.005)。经过低温处理不同时长后,3~6龄幼虫累计发育历期极显著延长(=62.002,<0.01),两者呈显著线性关系(=0.544+10.406,=0.808,=180.829,<0.01)。预蛹期差异均不显著(=0.860,=0.496),蛹期均极显著短于对照组(=11.687,<0.01),成虫期差异不显著(=1.935,=0.123)。由结果可知草地贪夜蛾3龄幼虫经短时低温处理后,3龄幼虫发育历期受影响最大,其次是5龄幼虫及3~6龄幼虫的累计发育历期,预蛹期和成虫期受影响较小。

注:a~i依次为3龄幼虫、4龄幼虫、5龄幼虫、6龄幼虫、3~6龄幼虫、 预蛹、 蛹、成虫、蛹重。图中数据为平均数±标准差。不同大、小字母表示同一龄期不同时长处理间经Duncan氏新复极差法检验在P<0.01和P<0.05水平差异显著。图2 经13 ℃低温处理后草地贪夜蛾各龄幼虫的发育历期及蛹重Fig.2 Effects of low temperature 13 ℃ on larval duration of 3rd~6th instars and pupal weight of S. frugiperda
2.2 高、低温处理对草地贪夜蛾死亡率的影响
..高温对各龄幼虫死亡率的影响
3龄幼虫经36 ℃高温不同时长处理后,随后各龄幼虫的死亡率与对照组差异显著(图3),高温处理后3龄幼虫死亡率均显著高于对照组(<0.05),其中处理32 h后平均死亡率达到60%~80%,显著高于对照组及其他处理组。高温处理16 h和24 h的4龄幼虫期死亡率显著高于对照组(<0.05),其中处理24 h的死亡率在0~60%,多集中在20%~50%之间。5龄幼虫期死亡率均显著低于对照组(<0.05),对照组死亡率在0~30%,多集中在0~15%之间。6龄幼虫期,处理8 h的死亡率在0~40%,处理16 h的有效死亡率在0~10%。3~6龄幼虫累计死亡率在高温处理24 h和32 h下均极显著高于对照组(<0.01)。结果表明36 ℃高温处理3龄幼虫不同时长后,在随后的发育过程中,死亡率受影响最大的是3龄、4龄幼虫,其次是3~6龄幼虫累计发育历期,5龄幼虫发育历期受影响最小。
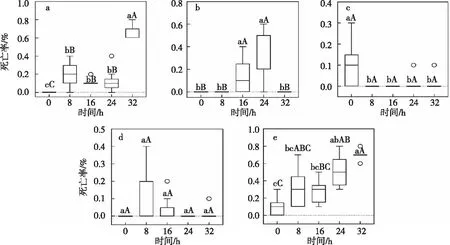
注:a~e依次为3龄幼虫、 4龄幼虫、 5龄幼虫、 6龄幼虫、3~6龄幼虫。利用克鲁斯卡尔—沃利斯单因素ANOVA检验(k个样本),并对多项检验结果进行Bonferroni校正法校正。图3 经36 ℃高温处理后草地贪夜蛾各龄幼虫死亡率的箱线图Fig.3 Box plot of mortality rate of each instar larvae of S. frugiperda treated at 36 ℃
..低温对各龄幼虫死亡率的影响
3龄幼虫经13 ℃低温不同时长处理后,对后续各个龄期死亡率的影响变化见图4,从检测数据上看,3龄幼虫死亡率各处理间差异均不显著(>0.05),处理16 h后的死亡率在0~30%,处理32 h的死亡率在0~20%。3龄幼虫低温处理8 h后4龄幼虫死亡率显著高于对照组(=0.027),在0~20%之间,其余处理组差异不显著(>0.05)。5龄幼虫发育历期所受影响趋于稳定,处理组间差异均不显著(>0.05)。6龄幼虫发育历期处理组的死亡率在一定空间内波动上升。处理组3~6龄幼虫累计发育历期死亡率中位数值高于对照组,但处理组间差异不显著(>0.05)。表明3龄幼虫在经过13 ℃低温处理后,各个发育过程的死亡率所受影响波动最大的是3、4龄幼虫发育历期幼虫,其次是3~6龄幼虫累计发育历期,5龄幼虫受影响最小。

注:a~e依次为3龄幼虫、 4龄幼虫、 5龄幼虫、 6龄幼虫、3~6龄幼虫。利用克鲁斯卡尔-沃利斯单因素ANOVA检验(k个样本),并对多项检验结果进行Bonferroni校正法校正。图4 经13 ℃处理后草地贪夜蛾各龄幼虫死亡率的箱线图Fig.4 Box plot mortality rate of each instar larvae of S. frugiperda treated at 13 ℃
2.3 高、低温处理对草地贪夜蛾蛹的影响
..高温对草地贪夜蛾蛹的影响
3龄幼虫经过高温处理8 h的蛹重与对照组无显著差异,处理16 h、24 h和32 h后其蛹重显著低于对照组(=3.281,=0.020)(图1-i)。经高温处理8 h和处理16 h的3龄幼虫,其化蛹率与对照组差异不显著,而高温处理24 h和处理32 h的3龄幼虫,其化蛹率极显著降低(=17.781,<0.01),分别为49.78%、65.00%(图5-b)。
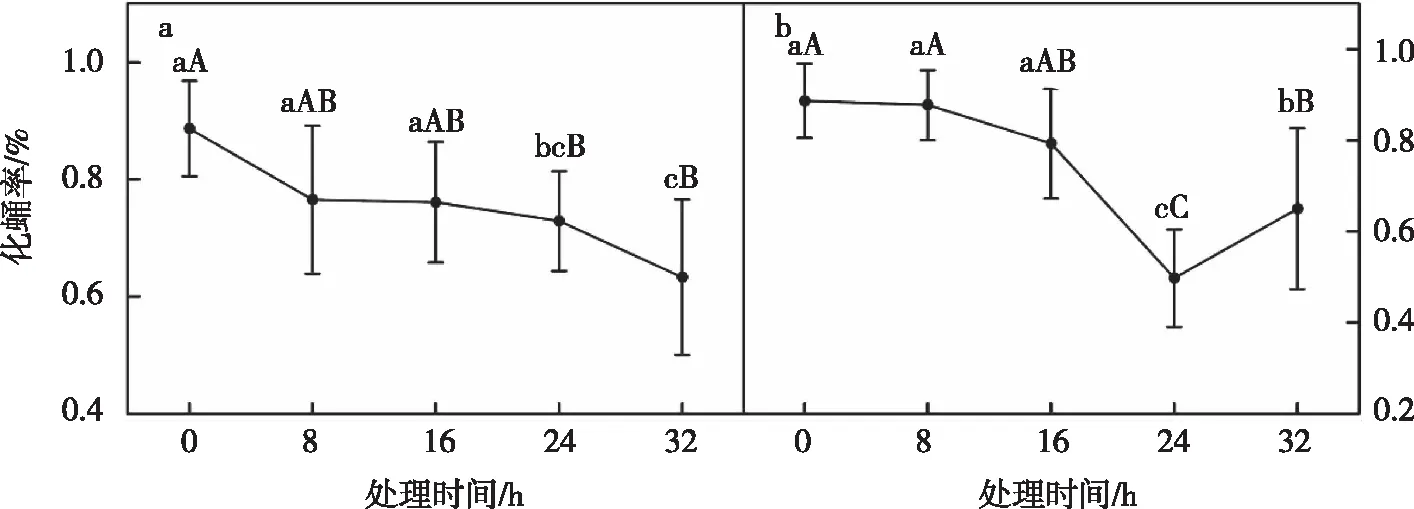
注:a为13 ℃化蛹率;b为36 ℃化蛹率。图中数据为平均数±标准差。不同大、小字母表示同一龄期不同时长处理间经Duncan氏新复极差法检验在P<0.01和P<0.05水平差异显著。图5 高、低温对草地贪夜蛾蛹的影响Fig.5 The effect of high and low temperature on the pupae of S. frugiperda
..低温对草地贪夜蛾蛹的影响
3龄幼虫经低温处理后,其蛹重极显著低于对照组(图2-i),其中处理24 h后蛹重降低至0.140 g(=7.463,<0.01)。经过低温处理后,幼虫化蛹率与对照组差异显著,其中处理8 h和16 h后化蛹率显著低于对照组(=6.384,<0.01),处理24 h和32 h的3龄幼虫的化蛹率分别为72.89%和63.33%,均极显著低于对照组的88.67%(图5-a)。
2.4 高、低温对草地贪夜蛾抗寒性的影响
3龄幼虫经高、低温处理不同时长后置于-7 ℃下胁迫后存活率由图6可知,36 ℃和13 ℃处理8 h后再胁迫的幼虫存活率均显著高于对照组(=5.515,=0.011);高、低温处理16 h后再胁迫的3龄幼虫存活率与对照组无显著差异(=2.161,=0.137),处理24 h和处理16 h的结论一致(=2.167,=0.136)。13 ℃处理32 h后胁迫的幼虫存活率与对照组差异不显著,36 ℃处理32 h后胁迫的幼虫存活率极显著低于对照组存活率(=7.579,=0.003)。基于数据检测可知,3龄幼虫经过36 ℃和13 ℃处理8 h后可以显著提高草地贪夜蛾的抗寒性,处理16 h、24 h无显著影响,高温处理32 h后再胁迫的抗寒性极显著下降,低温处理32 h后再胁迫对草地贪夜蛾抗寒性无显著差异。
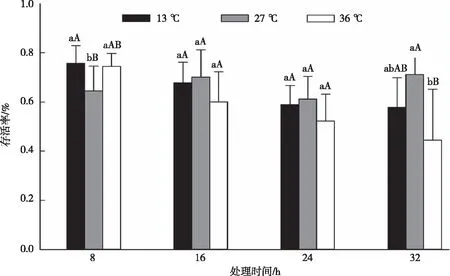
注:图中数据为平均数±标准差。不同大、小字母表示同一龄期不同时长处理间经Duncan氏新复极差法检验在P<0.01和P<0.05水平差异显著。图6 高、低温处理后经-7 ℃胁迫对草地贪夜蛾存活率的影响Fig.6 Effect of -7 ℃ stress on survival rate of S. frugiperda in control group and different temperature treatments
3 结论与讨论
温度是影响昆虫生长发育、繁殖、行为、生活史等生命活动的重要因素,当外界温度超过昆虫适生温度范围,昆虫生命活动就会受到一定程度的影响。如麦长管蚜、南亚实蝇、紫斑谷螟及沙葱萤叶甲经不同短时高低温处理后均不利于生长发育。本研究中草地贪夜蛾3龄幼虫经36 ℃高温胁迫后,发育历期受到不同程度的影响,其中3龄和4龄幼虫死亡率显著上升,预蛹期极显著缩短,3龄幼虫经高温处理24 h和32 h后化蛹率显著降低,随着高温处理时长的增加,蛹重显著下降;经13 ℃低温胁迫后,3龄及随后各龄发育历期整体呈延长趋势,4龄幼虫死亡率上升,低温处理后化蛹率和蛹重均显著降低。这与前人研究得到15 ℃时蛹存活率低;36 ℃下草地贪夜蛾卵孵化率下降,羽化过程死亡率高,37 ℃时蛹无法正常羽化;不同龄期幼虫经高温暴露对蛹重有显著影响等结论相似。表明在初春和夏季,草地贪夜蛾在遭受短时高低温胁迫后,对其各虫态生长发育与存活率均有不利影响。
昆虫的抗寒能力能够影响其来年的发生量,经过适宜的极端温度处理能提高昆虫的抗寒能力。本文在13 ℃下低温处理草地贪夜蛾3龄幼虫8 h,可提高其抗寒性,这与前人的研究结果一致。例如异色瓢虫的成虫经5 ℃胁迫一段时间后,再暴露于-5 ℃下,其存活率显著提高;东亚飞蝗的卵经0 ℃和5 ℃驯化后,均能显著提高其存活率。目前,关于高温胁迫对昆虫抗寒性影响的研究较少,本研究得出36 ℃高温处理草地贪夜蛾3龄幼虫可提高其幼虫存活率,这与斜纹夜蛾、黑腹果蝇的研究结果一致,均表明适宜的高温处理可提高部分昆虫的抗寒性。
草地贪夜蛾原产地属于热带和亚热带地区,高低温胁迫对其适应入侵地及种群繁衍有重要意义,本研究结果反映出短时高低温胁迫草地贪夜蛾对其生长发育存在不利的影响,但对其存活率有一定的提高作用。因此,在草地贪夜蛾发生的地区,当早春和夏季出现短时高低温后,应对草地贪夜蛾种群采取适当的控制措施。研究结论对进一步了解草地贪夜蛾生长发育、分布及迁移规律提供科学依据,对短时高低温频发条件下预测预报及防治有一定的参考价值。
猜你喜欢
天津农业科学(2022年6期)2022-07-17
汽车实用技术(2022年11期)2022-06-20
植物保护(2022年1期)2022-02-10
小天使·四年级语数英综合(2019年10期)2019-11-09
植物保护(2018年3期)2018-05-14
农村农业农民·B版(2016年7期)2016-10-21
中国机械(2015年3期)2015-10-21
祝您健康(2008年2期)2008-02-20
杂文选刊(2007年16期)2007-05-14
中国青年(1984年2期)1984-08-20
