
苹果砧木T337不同组织内生菌群落及其功能预测
2022-07-29 07:03张东华张俊忠
江苏农业科学 2022年14期
程 欢, 张东华, 张俊忠, 刘 丽
(西南林业大学保护生物学学院/云南省森林灾害预警与控制重点实验室,云南昆明 650224)
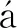
1 材料与方法
1.1 样品采集
苹果砧木M9T337(以下简称为T337)为二年生苗木,于2019年10月取自云南省昆明市西南林业大学树木园。苹果园的肥、水、修剪等管理方法参照常规措施。园中随机挖取无明显病虫害的植株4株,去除腐叶后,分别将根、枝、叶放入不同的无菌样品袋中混合均匀,再随机选取样品进行表面消毒处理,根(G)、枝(Z)、叶(Y)样品分别命名为G1~G4、Z1~Z4、Y1~Y4。
1.2 表面消毒
用无菌水将苹果根、枝、叶冲洗干净,用无菌纸吸干水分后,用75%乙醇浸泡30 s,将根、枝条和叶片分别用0.1%氯化汞振荡浸泡60、60、30 s,再用无菌水冲洗4次,最后用无菌滤纸吸干表面的水分。取最后1次清洗苹果组织的100 μL无菌水涂布在营养琼脂(NA)固体培养基上,于28 ℃培养72 h,用于检测灭菌效果。
1.3 总DNA的提取及扩增
对样品进行表面消毒后,加入液氮充分研磨,取适量转入灭菌EP管中,参照OMEGA试剂盒[Omega Mag-bind soil DNA kit(M5635-02)]说明书进行总DNA的提取。用1%琼脂糖凝胶电泳检测所提取的DNA纯度和浓度。
1.4 扩增子测序
通过Illumina MiSeq平台测序靶标DNA为16S rRNA基因V3~V4可变区(引物为343F、798R)和内转录间隔区1(ITS1)(引物为ITS5、ITS2)。将目标条带用凝胶纯化后构建文库,在根、枝、叶引物序列前插入6个碱基组成的标签序列,以区分根、枝、叶样品。由上海派森诺生物科技有限公司进行测序。
1.5 生物信息学分析
用FLASH、QIIME软件分别对不同样品的读长序列进行拼接、过滤。用VSEARCH软件检测并去除嵌合体序列,然后用UPARSE算法对优化序列在97%的相似性水平上进行操作分类单元(OTUs)聚类。用Greengene等数据库进行物种注释,去除注释为叶绿体、线粒体及非细菌界和真菌的OTUs。分别对细菌、真菌的OTUs进行α-多样性、β-多样性分析,同时对物种注释在各个分类水平上的群落结构进行统计。最后用PICRUSt软件进行功能和代谢途径的预测,并参照MetaCyc数据库(Encyclopedia of Metabolic Pathway)进行功能注释。
2 结果与分析
2.1 苹果砧木T337不同部位内生菌的OTUs分布及其多样性分析
苹果砧木T337不同部位内生细菌的OTUs共有11 386个。其中根、枝、叶共有134个(占3.53%)OTUs,特有OTUs数分别为9 175个(占80.51%)、500个(占4.39%)、749个(占6.58%)(图1)。由此可见,不同部位的内生细菌种类组成差异很大,且根部细菌的种类组成特异性高于叶、枝。基于OTUs数量及相对丰度,对不同部位苹果树内生细菌进行α-多样性分析,结果表明,Chao1指数、香农(Shannon)指数、辛普森(Simpson)指数及物种丰富度(observed species)指数均表现为根部显著高于枝部、叶部,而枝部和叶部间无显著差异(表1),可见根部内生细菌的多样性、物种丰富度显著高于枝部、叶部。
苹果砧木T337不同部位内生真菌的OTUs共有2 718个,其中根、 枝、 叶部共有221个(占8.13%)OTUs,特有OTUs分别为1 187个(占43.67%)、295个(占10.85%)、603个(占22.19%)(图2),说明不同部位内生真菌的种类组成差异较大。基于OTUs数量及相对丰度分析内生真菌α-多样性可知,Simpson指数表现为根部、叶部均显著高于枝部,但根部与叶部间的差异不显著。Chao1指数、Shannon指数及Observed species指数均表现为在根、枝、叶3个部位差异显著,且根>叶>枝。总体趋势都是根部内生真菌多样性、物种丰富度最高,枝部最低(表2)。综合表1、表2的结果可知,苹果砧木T337根部内生细菌Chao1指数明显高于内生真菌,由此推测其根部内生细菌的物种丰富度高于内生真菌。
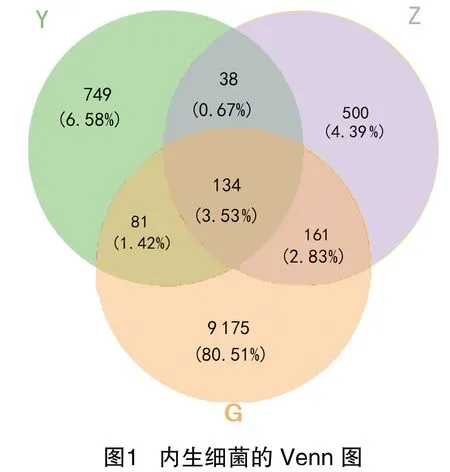
表1 内生细菌的α-多样性指数分析结果

表2 内生真菌的α-多样性指数分析结果
基于UniFrac距离的主坐标分析(PCoA)结果显示,对于苹果砧木T337组织样品而言,无论是内生细菌还是内生真菌都依据植物的部位在空间排布上呈现明显的聚类(图3)。枝与叶的内生细菌群落通过主成分1聚类,枝与根间通过主成分2聚类,其中主成分1的差异贡献率为27.45%,主成分2的差异贡献率为18.36%。
基于Jaccard距离的主坐标分析(PCoA)结果(图3-b)显示,不同部位内生真菌主坐标分析的主成分1、主成分2对样品的差异贡献率分别为17.5%、15.9%,其中主成分1是差异的主要来源,通过主成分1即可将叶与枝内生真菌群落聚类,叶、根内生真菌群落通过主成分2聚类。此外,无论是内生细菌还是真菌,其枝部组内样品间群落结构都具有更高的相似性。
2.2 苹果砧木T337不同部位内生菌在门水平上的组成和相对丰度
种群归类结果显示,苹果砧木T337根部所得内生细菌OTUs序列分属21个门、52个纲、75个目、154个科、319个属,枝部所得内生细菌OTUs序列分属12个门、24个纲、40个目、82个科、117个属,叶部所得内生细菌OTUs序列分属13个门、23个纲、36个目、69个科、93个属;根部所得内生真菌OTUs序列分属8个门、32个纲、88个目、195个科、332个属,枝部所得内生真菌OTUs序列分属4个门、24个纲、85个目、123个科、163个属,叶部所得内生真菌OTUs序列分属3个门、23个纲、79个目、139个科、213个属。

由图4可以看出,在门水平上,对相对丰度≥1% 的内生细菌进行分析比较发现,根部相对丰度较高的菌门依次为放线菌门(Actinobacteria,45.227%)、变形菌门(Proteobacteria,40.618%)、拟杆菌门(Bacteroidetes,3.091%)、酸杆菌门(Acidobacteria,2.593%)和Candidatus_Saccharibacteria(尚无统一的中文名,1.178%);枝部相对丰度较高的菌门依次为变形菌门(42.063%)、放线菌门(12.329%)、拟杆菌门(4.503%);叶部相对丰度较高的菌门依次为变形菌门(37.168%)、放线菌门(7.338%)和拟杆菌门(4.913%)。疣微杆菌门(Verrucomicrobia,0.123%)、衣原体门(Chlamydiae,0.020%)、Parcubacteria(尚无统一的中文名,0.007%)、Microgenomates(尚无统一的中文名,0.003%)、硝化螺旋菌门(Nitrospirae,0.002%)、梭杆菌门(Fusobacteria,0.001%)、Spirochaetes(尚无统一的中文名,0.001%)是根部特有的菌门。由此可以看出,在门分类水平上,枝部、叶部内生细菌群落组成更加相似,根部的最优势菌门为放线菌门,枝部、叶部的最优势菌门为变形菌门,但变形菌门在根、枝、叶3个部位的相对丰度均大于35%。
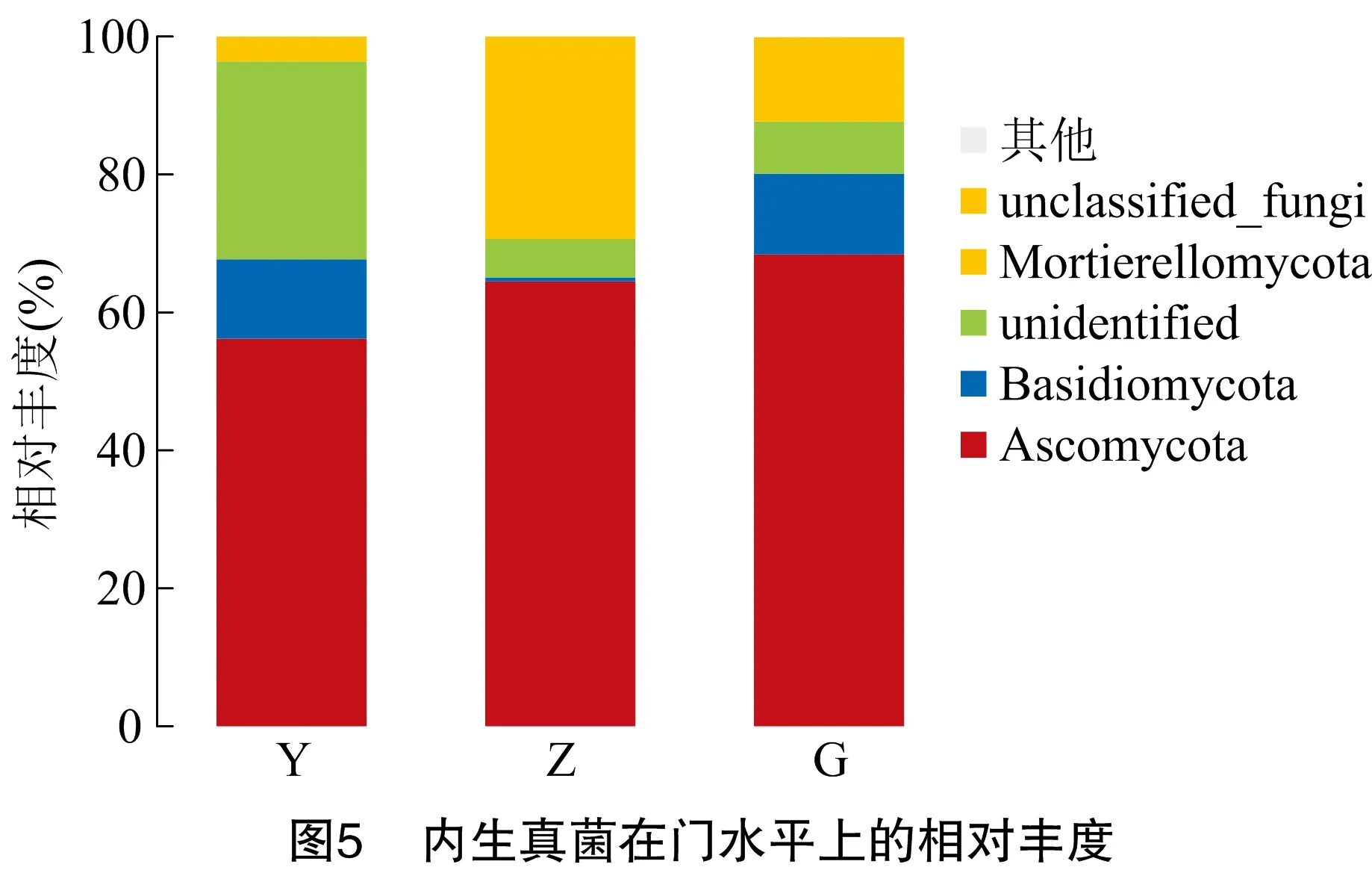
由图5可以看出,内生真菌在根部、枝部、叶部相对丰度≥1%的菌为子囊菌门(Ascomycota)和担子菌门(Basidomycota);根部、枝部、叶部最具优势的菌门均为子囊菌门(68.410%、64.500%、56.180%),次优势门为担子菌门(11.700%、0.581%、11.530%)。由此可见,枝部担子菌门内生真菌明显少于根部、叶部。Olpidiomycota(0.034%)、被孢霉门(Mortierellomycota)(0.020%)、 壶菌门(Chytridiomycota)(0.014%)和隐真菌门(Rozellomycota)(0.001%)是根部特有的菌门。球囊菌门(Glomeromycota)只存在于枝部、根部。

2.3 苹果砧木T337不同部位内生菌在属水平上的组成和相对丰度
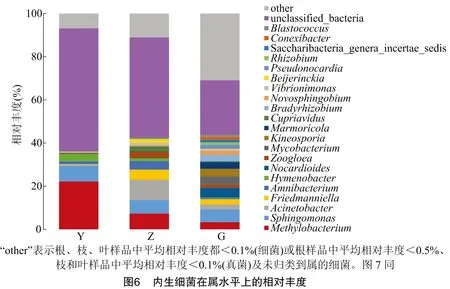
由图6可知,在属水平上,根部内生细菌相对丰度排名前十的优势属依次为鞘氨醇单胞菌(,5900)、类诺卡氏菌(,4.492%)、分直杆菌属(,4003)、动孢菌属(,3422)、甲基杆菌属(,3311)、大理石雕菌属(,3063)、慢生型大豆根瘤菌(,2870)、弗莱德门菌属(,2480)、不动杆菌属(,2303)、(1957);枝部内生细菌相对丰度排名前十的优势属依次为不动杆菌属(9431)、甲基杆菌属(7320)、鞘氨醇单胞菌(6221)、弗莱德门菌属(4711)、(3944)、(尚无统一的中文名,2845)、贪铜菌属(,2337)、(尚无统一的中文名,1670)、薄层菌属(,1325)、微生物弧菌属(,1059);叶部内生细菌相对丰度排名前十的优势属依次为甲基杆菌属(22174)、鞘氨醇单胞菌(7175)、薄层菌属(,3430)、(尚无统一的中文名,1227)、不动杆菌属(0654)、微生物弧菌属(0326)、(尚无统一的中文名,0280)、弗莱德门菌属(0262)、贪铜菌属(0212)、根瘤菌属(,0149)。根部内生细菌的特有属最多,有187个,其中相对丰度≥1的是假诺卡氏菌属(,1913);枝部特有属有5个,分别为(尚无统一的中文名,0006)、罗氏菌属(,0006)、(尚无统一的中文名,0004)、(尚无统一的中文名,0004)、泛菌属(,0004);叶部特有属有6个,分别为(尚无统一的中文名,0076)、(尚无统一的中文名,0035)、丛毛单胞菌(,0005)、沙雷氏菌属(,0005)、(尚无统一的中文名,0003)、蜂窝囊菌属(,0002)。另外,在根、枝、叶中还存在许多未归类的细菌(相对丰度分别为35885、50185、59884)。
由图7可知,在属水平上,根部内生真菌相对丰度排名前十的优势属依次为亚隔孢壳属(,7177)、枝孢属(,5077)、(尚无统一的中文名,5001)、异茎点霉属(,3983)、(尚无统一的中文名,2888)、(尚无统一的中文名,2757)、(尚无统一的中文名,2171)、(尚无统一的中文名,2003)、(尚无统一的中文名,1849)、外瓶霉属(,1752);枝部内生真菌相对丰度排名前十的优势属依次为(11804)、横断孢属(,2635)、(尚无统一的中文名,0661)、链格孢属(,0316)、拉奇枝孢霉(,0238)、担孢酵母属(,0221)、(尚无统一的中文名,0210)、亚隔孢壳属(,0182)、枝孢属(,0181)、(尚无统一的中文名,0147);叶部内生真菌相对丰度排名前十的菌属依次为壳针孢属(,22783)、链格孢属(6777)、枝孢属(6109)、亚隔孢壳属(,6103)、(尚无统一的中文名,3571)、(尚无统一的中文名,2728)、(尚无统一的中文名,1432)、(尚无统一的中文名,1196)、(尚无统一的中文名,1170)、隐球菌属(,1137)。根部内生真菌特有属最多,有121个,其中相对丰度≥005的属依次为根念珠属(,0145)、卡普龙属(,0115)、(尚无统一的中文名,0106)、(尚无统一的中文名,0093)、(尚无统一的中文名,0081)、木霉属(,0064)、(尚无统一的中文名,0063)、(尚无统一的中文名,0062)、(尚无统一的中文名,0055)、(尚无统一的中文名,0052);枝部特有属有7个,其中相对丰度≥0001的依次为(尚无统一的中文名,0007)、高氏白粉菌属(,0001);叶部特有属有35个,其中相对丰度≥001的属依次为假尾孢属(,0025)、(尚无统一的中文名,0017)、(尚无统一的中文名,0013)。另外,在根、枝、叶中还存在许多未归类的真菌(相对丰度分别为41031、82036、39467)。

2.4 功能预测分析
为了探究苹果砧木T337不同组织部位内生菌群的功能,本研究基于16S rRNA和ITS序列,采用PICRUSt2软件对内生细菌、内生真菌的功能进行预测。结果显示,从内生细菌中共预测到417个MetaCyc代谢通路,其中在植株不同部位间二级功能通路差异显著的有35个通路(<0.05);从内生真菌中共预测到73个MetaCyc代谢通路,其中在不同组织部位间二级功能通路差异显著的有19个通路(<0.05)。内生菌群的预测功能分别隶属于生物合成、降解/利用/同化、前体代谢产物和能量的产生、聚糖途径、其他代谢共5个方面(表3、表4)。
进一步分析内生细菌的功能发现,生物合成通路的相对丰度总体上高于其他通路,其中氨基酸合成通路的相对丰度最高,达到10%左右,其次为辅助因子、维生素合成通路的相对丰度,均达8%以上。内生细菌的氨基酸生物合成、芳香族化合物合成、核苷酸合成、次级代谢产物合成及细胞结构合成等通路的相对丰度均表现为叶部、枝部显著高于根部,而枝部和叶部间无显著差异。反之,上述生物大分子的分解反应对应的二级功能通路的相对丰度表现为根部显著高于叶部、枝部。在分解反应方面,无机盐、高分子化合物降解等二级功能通路的相对丰度表现为枝部、叶部显著高于根部。在碳水化合物生物合成和降解功能方面,相对丰度均表现为叶部、枝部显著高于根部。在能量代谢方面,发酵、糖酵解、无糖磷酸化、光合作用和呼吸作用等二级功能通路的相对丰度表现为枝部、叶部显著高于根部。异丙醇和甲基酮的合成、TCA循环、乙醛酸循环等二级功能通路的相对丰度则表现为根部显著高于枝部、叶部。嘧啶脱氧核糖核苷酸磷酸化的二级功能通路表现为枝部、叶部显著高于根部,其合成功能的相对丰度则是根部显著大于枝部、叶部(表3)。

表3 不同部位内生细菌预测的MetaCyc二级功能通路的相对丰度
对苹果砧木T337不同部位内生真菌群落功能的预测结果表明,大多数代谢通路的相对丰度在叶部和根部间存在显著差异。内生真菌核苷酸合成二级功能通路的相对丰度最高,达到44%左右,其次为脂肪酸和脂质的合成二级功能通路,其相对丰度达28%左右。在生物合成方面,内生真菌的碳水化合物生物合成、核苷酸合成等二级功能通路的相对丰度表现为叶部显著高于根部。在辅助因子和维生素的合成及脂肪酸、脂质的合成二级功能通路上,相对丰度则表现为根部、枝部显著高于叶部。但内生真菌相应的氨基酸降解、脂肪酸和脂质降解、核苷酸降解等分解反应二级功能通路的相对丰度也同样表现为叶部最高,这与内生细菌的功能预测规律不同。在能量代谢方面,甲基酮生物合成及呼吸作用等二级功能通路的相对丰度仍以叶部最高。在磷脂酶代谢及嘧啶脱氧核糖核苷酸磷酸化等二级功能通路上的相对丰度则是根部、枝部显著高于叶部(表4)。

表4 不同部位内生真菌预测的MetaCyc二级功能通路的相对丰度
3 讨论
微生物与植物相互依存的状态被认为是植物在自然界的真实存在状态,植物内生菌在生态作用上对宿主植物十分有利,并在此过程中形成了地域特异性、宿主保守性及组织专一性等适应性特点。在植物不同组织间内生菌多样性的研究中,有研究认为, 内生菌在植物地下部分的丰度高于地上部分,而地上部分的种类要多于地下部分。也有研究认为,植物组织内生菌多样性在不同宿主植物间并不具有一致性。本研究利用高通量测序方法分析了苹果砧木T337不同部位内生菌的多样性,结果表明,苹果砧木T337根部内生细菌的丰富度、多样性都显著高于枝部、叶部,内生真菌的则表现出根>叶>枝。对巨菌草、牛至及水稻的研究也发现,根部内生细菌的多样性均高于其他组织。翅碱蓬根中内生真菌群落的多样性要远远高于枝和叶。上述研究结果表明,内生微生物在地下部分有更高的多样性和丰富度,这与植物内生菌的来源息息相关,植物内生菌大部分来源于根际土壤微生物,后经根系运输到植株其他部位,但大气中的微生物也参与了内生细菌种群的构建。
放线菌门、变形菌门和拟杆菌门是苹果砧木T337根部、枝部和叶部的优势细菌门,其中放线菌门在苹果砧木T337根部样品中占绝对优势。已有研究发现,大多数植物内生细菌的最优势菌门为变形菌门,如番茄、黑果枸杞、牡丹、玉米、小麦等植物内生细菌的优势门均为变形菌门。但放线菌门在龙胆草等多种植物根际土壤中为最优菌门。变形菌门是苹果砧木T337枝部、叶部的最优菌门,在根部该菌门虽然不是最优菌门,但其相对丰度并不亚于枝部、叶部。变形菌门细菌可被用来促进氮肥的利用。Lupatini等研究发现,放线菌门细菌不但与磷酸盐、磷酸盐代谢相关,还可以产生农用抗生素、维生素,这暗示着开发苹果氮、磷吸收内生菌的潜力部位为苹果根部。疣微杆菌门只存在于T337根部,同时也是巨菌草根部的特有菌门。
苹果砧木T337内生细菌属水平的分析结果显示,不同部位优势属的相对丰度存在显著差异。鞘氨醇单胞菌属为根部最优菌属,研究发现鞘氨醇单胞菌属也是玉米根部的最优属,在其他作物中虽然不是最优属,但是也会有益于植物生长。Khan等从非洲山毛豆叶片中分离的sp. LK11可以产生吲哚-3-乙酸(IAA),从而增加植物生物量,促进植物生长。不动杆菌属为枝部的最优属,且广泛分布于植物中。Shi等从甜菜根部分离出的约氏不动杆菌可以促进土壤中微量元素的吸收,还可以产生生长素、溶解磷酸盐,进而促进植物生长。且不动杆菌属在林木宿主中被鉴定到的频率高于农作物。甲基杆菌属是叶部的最优势属,且在内生细菌中占有绝对优势。甲基杆菌属也是林木杨树和柳树叶、枝条的优势属,且作为甲基细菌可以在光合作用中为植物提供碳源,并诱导更高的光合作用活性。甲基杆菌属为自生固氮微生物,能参与植物的固氮,叶片中内生细菌可以通过固定大气中的氮或者保护植物免受病原菌的感染等机制影响植物生产力,促进植物生长。假诺卡氏菌属为植物根部特有菌属,麻疯树根部也存在假诺卡氏菌属细菌,前人研究发现,假诺卡氏菌属微生物主要与纤维素降解、抗生素合成有关,纤维素酶可以降解细胞壁,使植物根部和根际的微生物发生交换,且根系分泌物可以调节土壤通透性,从而间接地调节土壤对水分、温度的需求,并促进植物生长,根系分泌物还可以产生一些重要的酶类和维生素,为植物提供营养。动孢菌属、只存在于根部、枝部,但丰度较小,具体生物学功能还有待研究。动孢菌属是雷公藤枝部的优势属,研究发现,在三七地上部分的丰度高于地下部分,但是在苹果砧木T337的地上部位叶片中不存在。在以往的研究中,研究者从玉米、甘蔗、铁皮石斛、莽椒组织中都会分离到假单胞菌属(),且为优势属,但在苹果T337中,却不是优势属。由此可见,内生菌群落组成不仅由组织部位决定,而且可以通过植物-微生物组共同进化来解释。根瘤菌属为叶部样品的优势属,它广泛存在于豆科植物中,可以与植物共生并且具有固氮功能。
子囊菌门和担子菌门是苹果砧木T337根部、枝部和叶部的优势真菌门,且Ascomycota在根部、枝部和叶部中占绝对优势。另外,担子菌门在根部、叶部的相对丰度也较高。现有研究结果表明,多数植物的最优势内生真菌为子囊菌门。Olpidiomycota、被孢霉门、隐真菌门和壶菌门是苹果砧木T337根部的特有菌门。Rozellomycota在许多根际土壤中已有报道,例如茶树根际土壤。由于植物-内微生物-土壤微生物的相互作用,大多数内生菌来源于土壤微生物,这可能是导致根部特有门多于枝部、叶部的原因。苹果砧木T337内生真菌优势属种类和丰度随部位不同差异较明显。内生真菌根部最优属为,该属真菌可以分离获得很多化合物,且对我国医疗研究的发展具有很重要的作用;枝部最优属为,也是具柄重楼的优势属,但不是最优属,叶部最优属为壳针孢属,且在苹果砧木337内生真菌中占有绝对优势,广泛存在于植物叶片中,在T337中是否存在生物学功能还有待研究。除了关注不同部位内生真菌的优势属外,特有的丰度较低的属也在整个微生物组中扮演着重要角色。Harrison等研究发现,稀有内生菌会影响宿主大小、叶面积、叶片氮含量等,因此微生物组中丰度较小的内生菌也是不能忽视的。苹果砧木T337根部、枝部、叶部存在大量丰度较小的特有属,其中木霉属是苹果砧木T337根部的特有属,该属同样在杜鹃兰根部被检测到。贺字典等研究发现,木霉属不但可以拮抗多种植物病原菌,且可以促进辣椒根际土壤难溶性磷转化成速效性磷,从而促进辣椒植株生长,提高产量和品质。T337根部、枝部、叶部样品中都存在一定比例的植物病原菌属,尤其是叶部,这些真菌可能以条件性潜伏状态存在于植物组织中,也可能是有其他生物学功能,有待进一步研究证明。值得注意的是,苹果内生细菌和真菌中存在大量不能归类的序列,其中叶部未归类的内生细菌最多,占比大于50%,枝部未归类的内生真菌最多,占比达到80%;暗示着相应部位可能存在大量新类群。这也说明植物内生菌在研究方法上尚有很大的改进空间,研究者需从多方面进行改进,如多重16S rDNA测序、内生微生物的富集等。
PICRUSt软件是由美国哈佛大学的Curtis Huttenhower课题组开发的菌群代谢功能预测工具,通过将现有的基因测序数据与代谢功能已知的微生物参考基因组数据库相对比,实现对微生物代谢功能的预测。近年来,升级后的PICRUSt2功能预测的结果相比于PICRUSt更接近宏基因组的结果,从而提升了功能预测的精确度。且目前PICRUSt2输出的主要高水平预测为MetaCyc数据库通路。虽然PICRUSt2预测偏向于现有的参考基因,罕见的特定功能不太可能被识别出来,但是随着高质量可用基因数量的持续增加,这个限制对预测结果的影响也会逐渐减少。PICRUSt功能预测已经被应用在番茄和黑果枸杞等植物上,且可以采用KEGG数据库对内生微生物群落进行功能预测,古丽尼沙·沙依木等基于FAPROTAX数据库对不同品种苹果植株内生微生物功能进行了预测。本研究选用MetaCyc数据库注释结果,因为MetaCyc包含比KEGG更广泛的数据库属性集,包括从化合物到其所调节的酶的关系、自发反应的识别及预期的代谢途径分类范围等。
本研究通过PICRUSt2功能预测分析不同部位苹果砧木T337的功能差异,益生功能信息的合成功能表现为叶部、枝部的相对丰度显著高于根部,相应的生物大分子的分解功能则表现为根部的相对丰度显著高于枝部、叶部。前人研究发现,黑果枸杞根、茎、叶和果内生细菌基因中包含氨基酸类、脂类、维生素、多糖、萜类和酮类及其他次生代谢产物等与植物成分生物合成有关的通路。苹果T337也包含氨基酸、脂类、次级代谢产物等的合成通路。通过基因功能预测发现,莽椒、鲜广椒中内生细菌的碳水化合物运输与代谢功能相对较强,而苹果T337内生细菌的氨基酸代谢功能是最强的,其次是辅助因子、维生素的合成。Yang等研究发现,牡丹叶片中内生细菌光合作用丰度显著高于根组织,此结论与苹果T337的结果一致。植物的叶片主要是进行光合作用,相应的,叶部的内生细菌也是合成功能、光合作用功能的相对丰度更高。植物根为植物提供营养元素,根部内生细菌的分解功能的相对丰度较高。上述结论佐证了植物内生细菌与植物组织间相辅相成的功能关系,因此对苹果砧木T337不同部位内生微生物组的研究为今后内生微生物与植物的互作研究和植物健康有益微生物的筛选提供了参考。
4 结论
不同部位苹果砧木T337都具有丰富的内生菌资源。在根部、枝部、叶部3个组织部位中,根部内生细菌、内生真菌物种丰富度、多样性最高,枝部最低;在群落组成上,T337根部优势内生细菌门为放线菌门,枝部、叶部为变形菌门;优势内生真菌门在3个组织部位相同,均为子囊菌门;在属水平上,3个组织部位内生微生物都各有不同的优势菌属。无论是内生细菌还是内生真菌,特有菌属均为根部组织最多。在内生菌中,未归类的细菌叶部最多,未归类的真菌枝部最多。功能预测结果表明,内生细菌分解功能在根部样品中显著高于枝部、叶部,合成功能则相反;内生细菌更多地参与氨基酸的生物合成,内生真菌更多地参与核苷酸的生物合成。
猜你喜欢
农业工程学报(2022年6期)2022-06-27
中国烟草科学(2022年2期)2022-05-27
中国比较医学杂志(2020年4期)2020-05-26
环球时报(2019-10-24)2019-10-24
水生生物学报(2019年4期)2019-07-20
生物安全学报(2019年3期)2019-02-15
川北医学院学报(2019年6期)2019-02-10
湖北农业科学(2018年18期)2018-12-11
中国糖料(2016年1期)2016-12-01
