
沙漠腹地绿洲植物群落物种与系统发育多样性分布格局及驱动因子∗
2022-08-02 10:56史浩伯师庆东李孟泽安外尔阿卜杜热伊木
新疆大学学报(自然科学版)(中英文) 2022年4期
史浩伯,师庆东†,李孟泽,安外尔·阿卜杜热伊木,戴 岳
(1.新疆大学 生态与环境学院,新疆 乌鲁木齐 830017;2.绿洲生态教育部省部共建重点实验室,新疆 乌鲁木齐 830017;3.新疆大学 地理与遥感科学学院,新疆 乌鲁木齐 830017)
0 引言
生物多样性研究中,物种多样性与系统发育多样性均属于解析群落构建机制的基础指标之一[1−2].物种多样性研究指的是以物种为单元,以系统学为基础探讨物种的空间分布格局[3].而系统发育多样性则包含了物种之间的谱系关系与进化信息,在进化与保护生物学中占据着重要地位[4−5].随着生物学、生态学与进化学等学科的快速交叉与融合,以谱系研究为基础的系统发育研究方法在近些年被大量应用于群落生态学的相关研究之中[6−7].一般认为,系统发育多样性与物种多样性之间有着紧密的联系[8],系统发育多样性既能够以进化与生态的视角为物种多样性研究中的局限性提供补充信息,又能够在特定的时空范围内通过物种的系统发育进化史对该类群的综合生长状况进行评估,为群落构建过程中的物种共存与演化特征提供进一步的视角[9].除此之外,群落的系统发育多样性研究还有助于掌握某一演替阶段的系统发育结构,净谱系亲缘关系指数和净最近种间亲缘关系指数作为其中的主要指标[10],可以用作明确系统发育结构聚集与分散特征的重要方法,从而在一定程度上反映群落构建的内在机制[11−12],这对理解局域尺度上物种的分布特征具有关键意义.
绿洲是干旱半干旱地区的典型景观,是以大尺度荒漠背景为基质,具有一定生物群落基础,且以“岛屿”状存在于荒漠盆地或山前冲积平原的特有绿地[13].水是制约绿洲规模的关键因素,水资源空间分布格局限定了绿洲的格局,呈现出“有水则为绿洲,无水则为荒漠”的基本特征[14−16].达里雅布依绿洲位于中国塔里木盆地,是克里雅河深入到塔克拉玛干沙漠腹地形成的冲积绿洲.该绿洲除受地下水条件的制约外,每年丰水期还会受到地表水干扰造成的漫溢影响,二者的联合作用形成了干旱区荒漠湿地生态系统,但在此背景下,绿洲植物群落物种与系统发育多样性对环境的响应机制尚未明确.本文以达里雅布依绿洲为研究对象,通过设立地表水、地下水影响梯度为主要手段,分析不同梯度下物种多样性与系统发育多样性的格局特征,并探讨不同梯度下多样性格局的影响因素,旨在揭示不同环境因子在干旱区荒漠生态系统多样性维持中的作用及重要意义.
1 研究区概况
克里雅河是我国塔里木盆地南缘的第二大河流,地处新疆维吾尔自治区于田县境内,全长530 km.河流发源于昆仑山主峰,经于田县绿洲后继续蜿蜒向北,深入塔克拉玛干沙漠腹地,并在尾闾形成达里雅布依绿洲.该绿洲地理位置为38˚16′∼38˚37′N、81˚05′∼81˚46′E,海拔在1 100∼1 300 m之间,总面积约342 km2.达里雅布依绿洲所在区域为典型的暖温带干旱沙漠气候,年均降水量低于20 mm,潜在蒸发量高于2 000 mm,年均温12.1 ℃,昼夜温差大,气候极端干旱[17].绿洲内部具有错综复杂的网状河相沉积特征,每年存在丰水期与枯水期,受此影响导致绿洲在局域尺度上存在不同强度的地表水漫溢情况[18].绿洲群落主要由胡杨(Populus euphratica)、柽柳(Tamarix chinensis)、芦苇(Phragmites australis)等物种组成[19],本课题组于绿洲内修建了地下水监测井,为地表水、地下水梯度下的群落生态学研究提供了理想场地.
2 研究方法
2.1 植被调查
为了充分考虑绿洲环境异质性对群落的影响,2018年底,根据与主河道之间距离的远近设置了7条由南向北分布的样带(图1).参考绿洲形状与面积的变化在每条样带内植被连续分布区域建造了数量不同的地下水监测井,以此进行长时间序列的地下水埋深监测.植被调查时间选在2021年夏季,在已建好的监测井附近共选取了19处50 m×50 m的样地,并划分为4个25 m×25 m的样方,共计76个.在每个样方内进行植被调查工作,记录了各物种的数量、高度、冠幅等指标.对于草本植物生长茂盛的区域,在样方内随机布设了5个1 m×1 m的草本小样方,以此调查草本植物的多度、高度、盖度等指标,在对取样面积与取样个体进行换算后得出常规样方中草本植物的生态特征;而对于草本植物稀疏的区域,则采用与乔灌木相同的每株检尺调查方法.研究过程中发现存在5个单物种样方,受物种与系统发育多样性在指数计算与统计上的限制影响将其剔除.调查结束后运用手持GPS记录各样地的经纬度信息.
2.2 土壤因子调查
每个样地中随机挖取3处深度为1 m的土壤剖面,分为6层(0∼5 cm、5∼20 cm、20∼40 cm、40∼60 cm、60∼80 cm、80∼100 cm),将不同剖面的同一深度土壤进行充分混合后,作为代表群落水平的土壤因子,并采用四分法获取室内实验所需的土壤分量.将获取的土壤样品放置在铝盒与塑封袋内保存,用于测量土壤理化性质,具体包括土壤含水率、pH值、电导率、有机质、全氮与全磷.
2.3 地表水、地下水梯度划分
对近5年(2017―2021年)的Landsant 8遥感影像的水体指数进行了提取,进行阈值分割后获取了地表水分布的频次特征,为量化地表水对群落的干扰作用,创建公式如下:
为充分考虑手持GPS在样地定位过程中存在的误差,S代表记录的样点所在1 ha范围内地表水的像元数量;wi代表第i个像元在监测时限内的频次;SWD代表该局域范围内综合考虑漫溢频次与漫溢水量后的地表水干扰程度.该值越高,表示群落尺度范围内造成的地表水干扰程度越大.结合地下水监测井建成以来生长季(4月至10月)的地下水埋深数据,通过K均值聚类将地表水、地下水对绿洲群落的干扰程度划分梯度(图2.A:低地表水干扰与深地下水埋深;B:低地表水干扰与浅地下水埋深;C:高地表水干扰与浅地下水埋深;Sp:样地编号).
2.4 物种多样性计算方法
在事先求得不同群落物种重要值的前提条件下,选取Shannon-Wiener多样性指数(H)、Simpson多样性指数(D)、Pielou均匀度指数(J)作为物种多样性计算的主要手段.其中:Shannon-Wiener指数来源于信息学,在同时考虑物种丰富度与均匀度的情况下用以描述物种出现的随机性与不确定性[20];Simpson指数来源于概率论,通过描述随机取样个体相同的可能性量化物种的多样性[21];Pielou指数表示物种在相应群落中分布的均匀程度[22].三者的具体公式如下:
式中:Pi为对应群落中第i个物种的重要值;S为总物种数.
2.5 系统发育结构与多样性研究方法
运用R 4.1.2中的“plantlist”包,将绿洲统计到的所有物种通过APG Ⅲ被子植物分类系统进行校对[23],在修正好正确的物种名录后运用“V.PhyloMaker”包进行系统发育树的构建[24],并导入MEGA 7.0中,对形成的系统发育树进行可视化操作(图3).
选取系统发育多样性指数(PD)、净谱系亲缘关系指数(NRI)、净最近种间亲缘关系指数(NTI)进行系统发育多样性方面的研究[25],具体公式如下:
式中:Lb为系统发育树量化后的分支长度;MPDs和MNTDs分别为平均成对系统发育距离的观测值和最近相邻系统发育距离的平均观测值;MPDmds和MNTDmds为随机999次模拟出的平均成对系统发育距离和最近相邻系统发育距离的平均值;SD为标准差.若NRI与NTI的结果均大于0,则代表群落的系统发育结构趋于聚集;若NRI与NTI的结果均小于0,则代表群落的系统发育结构趋于分散;若NRI与NTI的结果等于0,则代表系统发育结构趋于随机.
2.6 驱动因子分析
在对地表水、地下水不同梯度下物种多样性与系统发育多样性的驱动因子进行分析前,采用R语言中的“vegan”程序包进行DCA检验,结果显示第一轴的长度均小于3,因此采用基于线性模型的RDA分析进行解释变量与响应变量之间的多元回归.为排除环境因子之间可能存在的共线性,由高到低依次筛选掉共线性诊断过程中VIF>10的方差膨胀因子,将筛选后的环境因子纳入解释变量,并通过层次分割(Hierarchical Partitioning)与999次的随机置换检验(Permutation Test)量化不同解释变量的贡献率与显著性[26].
3 结果与分析
3.1 物种与系统发育多样性在不同梯度下的格局特征
基于地表水与地下水形成的三种梯度,计算了不同梯度范围内物种Alpha多样性的具体特征(表1).结果表明:各梯度Shannon-Wiener指数与Simpson指数之间的差异特征趋于一致,均为低地表水干扰、浅地下水埋深梯度下(B)的多样性指数最高,且与其它两个梯度存在显著性差异.而在物种的Pielou均匀度方面,第一梯度(A)与第二梯度(B)之间的差异不大,二者均显著高于第三梯度(C).绿洲群落的系统发育多样性的PD值在第一梯度下(A)最低,其余两个梯度之间没有明显的差异;净谱系亲缘关系指数NRI与净最近种间亲缘关系指数NTI随着梯度的顺序逐渐降低,系统发育结构呈分散格局.
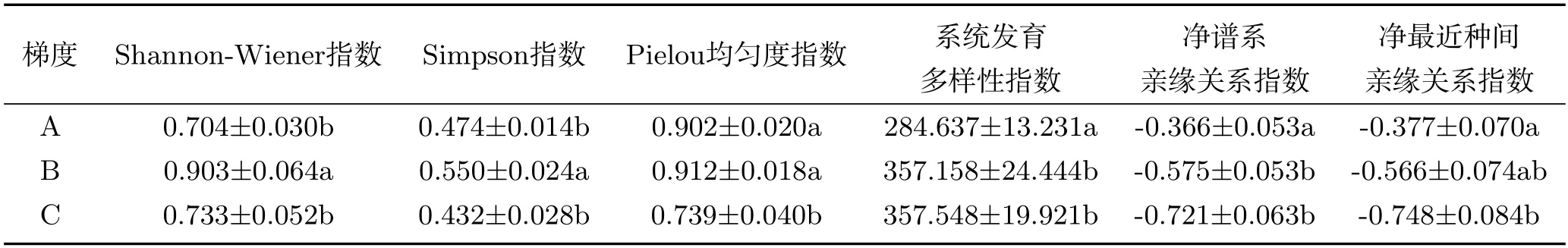
表1 绿洲群落物种多样性与系统发育多样性在不同地表水、地下水梯度下的变化
3.2 物种多样性与系统发育多样性的相关性分析
进行不同变量相关性分析之前,对观测数据的总体正态性进行了检验,结果显示不符合正态分布,因此选用Spearman相关性分析方法.相关性分析结果如图4所示.
物种多样性中,Shannon-Wiener指数与Simpson指数呈极显著的正相关关系(P <0.01),Simpson指数与Pielou均匀度指数呈极显著的正相关关系(P <0.01).系统发育多样性中,PD指数与NRI、NTI指数均呈极显著的负相关关系,而NRI与NTI指数之间呈极显著的正相关关系(P <0.01).物种多样性指数与系统发育多样性指数之间,PD指数与Shannon-Wiener指数、Simpson指数之间均呈极显著的正相关关系(P <0.01),与Pielou指数之间呈极显著的负相关关系(P <0.01),而NRI、NTI两个系统发育结构指数与Pielou指数之间也呈极显著的正相关关系(P <0.01).
3.3 物种多样性格局的驱动因子分析
在低地表水干扰与深地下水埋深梯度下(A),环境变量共解释了35.51%的物种多样性格局特征,其中土壤有机质的单独贡献率最高,为26.58%,经置换检验后显示该因子的层次分割结果显著,是该地表水、地下水梯度下物种多样性格局的主要驱动因子.在低地表水干扰与浅地下水埋深梯度下(B),环境变量共解释了44.76%的物种多样性格局特征,其中:土壤pH的单独贡献率最高,为23.02%;其次是全磷的12.46%,但土壤全磷的置换检验结果不显著,仅有土壤pH是该梯度下物种多样性格局的主导因子.在高地表水干扰与浅地下水埋深梯度下(C),环境变量共解释了47.73%的物种多样性格局特征,其中土壤电导率具有23.59%的单独贡献率,且层次分割后的置换检验结果显著,此外土壤全氮的贡献率也较高,为15.36%,但并不具备显著性,因此土壤电导率是该梯度下物种多样性格局特征的主要驱动因子(图5).
3.4 系统发育多样性格局的驱动因子分析
在低地表水干扰与深地下水埋深梯度下(A),环境变量共解释了28.86%的系统发育多样性格局特征.其中:土壤有机质的单独贡献率最高,为19.57%;其次是土壤含水率,具有6.13%的单独贡献率;土壤pH的贡献率则略低,为3.16%;土壤有机质在置换检验中呈现显著结果,系统发育多样性格局在此梯度下主要受该因子驱动.在低地表水干扰与浅地下水埋深梯度下(B),环境变量共解释了54.93%的系统发育多样性格局特征.其中:土壤pH具有25.20%的单独贡献率;其次是土壤全磷,具有16.46%的单独贡献率;二者的置换检验结果显著,是该梯度下系统发育多样性格局的主要驱动因子.在高地表水干扰与浅地下水埋深梯度下(C),环境变量共解释了34.73%的系统发育多样性格局特征.其中土壤电导率的单独贡献部分占比最高,为19.56%,且置换检验结果显著,是该梯度下驱动系统发育多样性格局的主要因子(图6).
4 讨论
4.1 地表水、地下水梯度变化下的物种多样性分布格局
通过对水资源丰富程度设立梯度,在此范围内同时考虑地表水与地下水过程,这是解析达里雅布依绿洲群落多样性格局的基础.物种多样性方面,低地表水干扰与浅地下水埋深这一梯度下(B)表现出了更适合多物种共存的特性.而在此基础上加大地表水干扰的情况下(C),物种的多样性并未继续增加,Shannon-Wiener指数与Simpson指数反而降为与第一梯度(A)同一水平,且均匀度指数降为三个梯度内的最低值.这是由于随着地表水干扰的增加,部分群落会以耐水淹、耐盐性的多年生草本植物为主[27],如本文中以芦苇为优势物种的顶级群落均出现在这一梯度内,这与艾克热木·阿布拉等对于塔里木河生态输水条件下台特玛湖植被格局的变化研究结果趋于一致[28].此外还有研究表明,即使同处地表水干扰造成的河水漫溢区域,物种的共存状况还与该区域是否为渍水区或冲刷区有关,长期渍水区域会使地表出现积盐,使得仅有耐盐能力突出的物种存活,而冲刷区则会令浅根植物在尚未成功定殖时即被冲走,这两种机制均可能成为导致物种多样性降低的主要原因[29].
4.2 系统发育结构变化及其成因
传统生态学中,生态位分化一直是群落构建的核心机制之一[30],在物种共存理论中占据着重要地位.在此基础上,关于群落的系统发育结构存在一个核心假设,即近缘种的共存主要受到环境过滤的影响,导致系统发育结构呈聚集格局;而远缘种的共存则主要受到竞争排除的影响,导致系统发育结构呈分散趋势[31].基于此,随着达里雅布依绿洲地下水与地表水的增多,满足了更多物种对可利用资源的要求,然而研究结果显示NRI与NTI的值逐渐降低,系统发育结构愈发分散,这反映了占据相似生态位的物种之间存在较为激烈的竞争关系,最终在这两个梯度下促进了远缘物种的共存,此条件下的竞争排除作用很可能占据了主导地位.但需要注意的是,竞争排除作用也极有可能会淘汰生态位相似但竞争能力较弱的远缘物种,理论上也会导致系统发育结构的聚集[32].在美国新泽西州一项利用50多年连续监测数据的群落谱系结构的研究中,研究者认为系统发育结构的发散是由远缘物种不断定居造成的,物种的灭绝并非由于竞争排除作用,而是受随机过程主导[33],这在一定程度上满足了中性理论的假设[34],也是对传统系统发育结构成因的挑战;该研究同时认为若群落在演替过程中出现了环境改良作用(Environmental Modification),即会出现阶段性的远缘物种定殖,系统发育结构也能够从聚集转为发散.不可否认的是,无论是基于已有资源的竞争排除还是环境改良导致的远缘种定殖,环境因素均具有重要的作用.因此,在今后的研究中,如何平衡确定性过程与随机性过程将是本研究区域的工作重点,进而科学且全面地揭示沙漠腹地绿洲群落构建的驱动机制.
5 结论
(1)克里雅河尾闾形成的达里雅布依绿洲中,物种多样性在低地表水干扰与浅地下水埋深梯度下呈膨胀格局.以此为基础,地表水干扰的增强与地下水埋深的增加均会降低物种的多样性.群落的系统发育多样性在低地表水干扰与深地下水埋深的梯度下最低,随着地下水埋深的变浅与地表水干扰的加剧,系统发育多样性有所提升且系统发育结构愈发分散.
(2)系统发育多样性指数(PD)与物种多样性各指数之间均具有显著的相关关系,而系统发育结构指数(NRI、NTI)仅与物种多样性指数中的Pielou均匀度存在显著的正相关关系.
(3)在低地表水干扰与深地下水埋深梯度下,驱动物种多样性与系统发育多样性格局的主导因子均为土壤有机质;在低地表水干扰与浅地下水埋深梯度下,驱动物种多样性格局的主导因子为土壤pH,驱动系统发育多样性格局的主导因子除土壤pH外还有全磷;在高地表水干扰与浅地下水埋深梯度下,驱动物种多样性与系统发育多样性格局的主导因子均为土壤电导率.
猜你喜欢
中国设备工程(2022年19期)2022-10-12
当代水产(2022年8期)2022-09-20
九江学院学报(自然科学版)(2022年2期)2022-07-02
昆明医科大学学报(2022年2期)2022-03-29
今日农业(2021年15期)2021-10-14
学苑创造·A版(2021年5期)2021-06-28
学生天地(2020年19期)2020-06-01
学校教育研究(2020年7期)2020-04-09
华东师范大学学报(自然科学版)(2019年3期)2019-06-24
北京航空航天大学学报(2017年12期)2017-04-23
