
毛乌素沙地不同品种沙棘光合特性研究
2022-08-02 14:22白慧慧史建国贺一鸣段义忠
榆林学院学报 2022年4期
白慧慧,鲁 客,史建国,贺一鸣,折 倩,段义忠
(榆林学院 生命科学学院,陕西 榆林 719000)
沙棘(Hippophae rhamnoides L.)是一种落叶性灌木,属胡颓子科沙棘属,具有抗寒抗旱等特点,且生命力顽强[1]。沙棘根、茎、叶、花、果,特别是沙棘果实含有丰富的营养物质,以其研发的产品既可食用,又可药用。沙棘多见于中国西北部,常被用于绿化沙漠,是治理沙漠化的重要植物[2],同时沙棘还是西北干旱区重要的经济灌木[3]。在高乙萍[4]与李丽霞[5]等研究进展中可知,沙棘在经济、社会及生态效益上具有重要的战略意义,因此沙棘成为治理毛乌素沙地的优选材料[6]。
光是植物生长和发育的动力来源[7],植物生长发育过程中光合作用为植物最主要的生理活动之一,作物生物产量的90%~95%都来源于光合产物[8],另外的5%~10%来源于根系吸收的营养物质,故光合作用可为作物产量的积累提供大量物质基础,研究光合参数的变化规律可在一定程度上探究光合特性与产量的相关性[8]。梁振娟等[9]认为光合作用对马铃薯产量有决定性作用;谢文钢等[10]研究结果表明,对不同品种茶树的光合特性进行比较,可进一步明晰该物种的光合性能与生产潜力;李雪琴等[11]对多种蕨类植物光合特性探究时发现,蕨类植物的光合特性既具有相似性又存在物种特异性;申慧芳等[12]研究发现,光合特性对选育优良种质资源具有一定的参考性;殷东生等[3]研究显示沙棘作为喜光物种,应选择在光照充足的地区进行栽培。杨紫玲等[13]在探究种子萌发进程时得知在强光下种子萌发更快,且沙棘种子萌发温幅较宽;桑林等[14]通过净光合速率(Pn)明确了矾根在昆明的适应性;文军等[15]研究表明蒸腾速率(Tr)能表征植物对环境的响应;史梦霞等[16]基于作物水分利用效率(WUE)和降水耦合度的综合评估对调整作物种植结构有重大意义。
目前对沙棘的研究主要集中在分析沙棘根瘤内生菌的多样性[17]、探究沙棘生理特性[18]、沙棘的抗逆性及生态适应机制[19]及沙棘的形态解剖特征[20]等方面,而基于光合特性选育高光效沙棘品系、挖掘优质的沙棘种质资源的相关研究较少。本研究对毛乌素沙地不同沙棘品种结合光合因子进行综合分析,通过研究不同沙棘品种的光合属性,不仅明晰了沙棘的生理特征,而且为毛乌素沙地优良沙棘品种的选育及栽培、沙棘产业化发展提供理论依据。
1 材料与方法
1.1 试验地概况
试验地布设于陕西省榆林市榆阳区马合镇,该镇位于陕西省榆林市榆阳区西北部,毛乌素沙地南缘,地势平坦,属温带半干旱大陆性季风气候,气温四季明显,冷暖有序,春温高于秋温,春季升温快而不稳定,秋季降温迅速,冬季受北方冷气团控制,气压高、天气晴朗,多高云。马合镇年平均气温7.9~11.39 ℃,日照时数为2 593.5~2 914.2 h;无霜期134~169 d;年均降水量400 mm左右,年蒸发量在1 152.2~1 289.7 mm;降水集中在夏季,6~9月降水量占年均降水量的74.4%,约为281.9 mm。
1.2 试验方法
1.2.1 试验设计
试验材料为山西引进的6个沙棘品种,分别为‘实优1号’‘深秋红’‘北欧1号’‘中国沙棘’‘无刺大果’以及‘小果雌株’。于2021年9月上旬,随机选择各品种沙棘3株,利用Li-6400便携式光合作用测定仪,在8∶00~18∶00期间,每间隔2 h测定其靠顶端的完整叶片,最后通过计算平均值作为该时次的测量结果,同时使用土壤温湿度记录仪(TMS)对表层(0~5 cm)土壤含水量进行多次测定得到平均值21%。
1.2.2 测定指标及方法
测定过程中及时记录净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)、水分利用效率(WUE=Pn/Tr)等参数。
1.3 统计分析
运用Excel 2019统计各项指标的平均值与标准误,采用SPSS 19.0统计分析软件进行单因素方差分析,采用Duncan’s新复极差法对同一时间段不同品种的沙棘进行显著性分析(P=0.05),以Pearson相关性分析光合生理指标间的相关关系,以ward(沃尔德)法进行聚类分析[21],用Sigmaplot 12.5作图。
2 结果与分析
2.1 光合生理参数日变化特征
2.1.1 净光合速率日变化
沙棘各品种净光合速率(Pn)日变化有所不同(图1a),‘实优1号’‘深秋红’‘北欧1号’‘中国沙棘’与‘小果雌株’的净光合速率均在8∶00~10∶00出现了最高值,分别为10.44 μmol/(m2·s)、6.66 μmol/(m2·s)、5.18 μmol/(m2·s)、3.32 μmol/(m2·s)及3.22 μmol/(m2·s);‘实优1号’‘北欧1号’、‘无刺大果’的谷值出现点均相同,在14:00~16:00,分别为1.62 μmol/(m2·s)、0.60 μmol/(m2·s)及1.35 μmol/(m2·s)。其中‘实优1号’与‘北欧1号’的净光合速率变化较为特别,基本呈现为早晚高、中午低的“V”形趋势;而‘深秋红’、中国沙棘、‘小果雌株’均呈现先降低再升高最后降低的较一致变化趋势。与其他品种不同的是‘无刺大果’在10∶00~12∶00出现了峰值,为2.65 μmol/(m2·s)。净光合速率日变化最高值由大到小顺序为‘实优1号’‘深秋红’‘北欧1号’‘中国沙棘’‘小果雌株’‘无刺大果’。沙棘各个品种净光合速率在8∶00~10∶00存在显著性差异(F=5.104, P=0.01),净光合速率日均值由大到小的顺序为‘实优1号’‘深秋红’‘北欧1号’‘小果雌株’‘中国沙棘’‘无刺大果’。
2.1.2 蒸腾速率日变化
各品种沙棘叶片的蒸腾速率(Tr)随光强的变化在不同时间段表现出一定的差异(图1b)。‘实优1号’‘深秋红’‘北欧1号’‘中国沙棘’‘无刺大果’及‘小果雌株’的蒸腾速率日变化趋势均呈现单峰型。‘北欧1号’‘无刺大果’及‘小果雌株’峰值出现较早,均在10∶00~12∶00,分别为1.64 mmol/(m2·s)、1.44 mmol/(m2·s)、2.29 mmol/(m2·s),其中‘北欧1号’与其他5个品种相比,其蒸腾速率变化较小;而‘小果雌株’蒸腾变化趋势较大;‘实生1号’与‘深秋’红的峰值出现于12∶00~14∶00分别为1.68 mmol/(m2·s)、1.98 mmol/(m2·s);而中国沙棘的峰值出现较晚,在14∶00~16∶00,为1.82 mmol/(m2·s)。蒸腾速率日变化最高值由大到小顺序为‘小果雌株’‘ 深秋红’‘中国沙棘’‘实优1号’‘北欧1号’‘无刺大果’。沙棘各个品种的蒸腾速率在16∶00~18∶00存在显著差异(F=3.422, P=0.038),蒸腾速率日均值由大到小的顺序为‘小果雌株’‘北欧1号’‘深秋红’‘实优1号’‘无刺大果’‘中国沙棘’。
2.1.3 气孔导度日变化
6个沙棘品种的气孔导度(Gs)日变化与其蒸腾速率日变化趋势在一定程度上具有相似性(图1c)。‘实优1号’‘深秋红’‘中国沙棘’‘无刺大果’在8∶00~10∶00出现了最高值,而‘北欧1号’与‘小果雌株’的峰值出现于10∶00~12∶00。气孔导度日变化最高值由大到小顺序为‘深秋红’‘小果雌株’‘无刺大果’‘实优1号’‘北欧1号’中国沙棘。沙棘各个品种的蒸腾速率在12∶00~14∶00存在显著差异(F=3.276, P=0.043),蒸腾速率日均值由大到小的顺序为‘深秋红’‘北欧1号’‘小果雌株’‘实优1号’‘中国沙棘’‘无刺大果’。
2.1.4 胞间CO2浓度日变化
各品种沙棘叶片胞间CO2浓度日变化基本与蒸腾速率和气孔导度变化规律一致(图1d)。‘实优1号’‘深秋红’‘北欧1号’及‘小果雌株’的胞间CO2浓度日变化趋势均呈现单峰型。‘实生1号’与‘深秋红’的峰值出现于12∶00~14∶00;‘北欧1号’的峰值出现于14∶00~16∶00;中国沙棘与‘无刺大果’的最高值出现于8∶00~10∶00,其中‘无刺大果’的变化趋势基本呈现为早晚较高、中午略低的“W”形趋势。胞间CO2浓度日变化最高值由大到小顺序为 ‘无刺大果’ ‘小果雌株’‘实优1号’‘深秋红’‘北欧1号’‘中国沙棘、。沙棘各个品种的胞间CO2浓度在12∶00~14∶00存在显著性差异(F=3.398, P=0.038),蒸腾速率日均值由大到小的顺序‘小果雌株’‘北欧1号’‘无刺大果’‘深秋红’‘实优1号’‘中国沙棘’。
2.1.5 水分利用效率日变化
各个品种沙棘的水分利用效率(WUE)与净光合速率的变化趋势较为一致(图1e)。除‘无刺大果’外,其余5个品种的最高值均出现于8:00~10:00,这与净光合速率日变化的最高值高度吻合;而‘无刺大果’的最高值时间点为12∶00~14∶00。水分利用效率日变化最高值由大到小顺序为‘实优1号’‘深秋红’‘北欧1号’‘中国沙棘’‘无刺大果’‘小果雌株’。沙棘各个品种水分利用效率在12∶00~14∶00存在显著差异(F=3.311, P=0.041),水分利用效率日均值由大到小的顺序为。‘实优1号’‘深秋红’‘中国沙棘’‘无刺大果’‘北欧1号’‘小果雌株’。

图1 不同沙棘品种光合指标日变化
2.2 光合生理参数日累积量的差异性
‘小果雌株’的Tr与Ci的日累积量均高于其他品种,除‘北欧1号’外,‘小果雌株’的Tr累积量与其他品种差异显著(P<0.05),‘小果雌株’的Ci累积量与5个品种间差异均显著(P<0.05)。‘实优1号’的Pn及WUE累积量显著大于其他品种(P<0.05),并与其余5个品种差异均显著,(P<0.05)(图2a与图2e);‘深秋红’的Gs累积量在6个品种中呈现出最大值,与其余品种间差异显著(P<0.05)(图2c)。
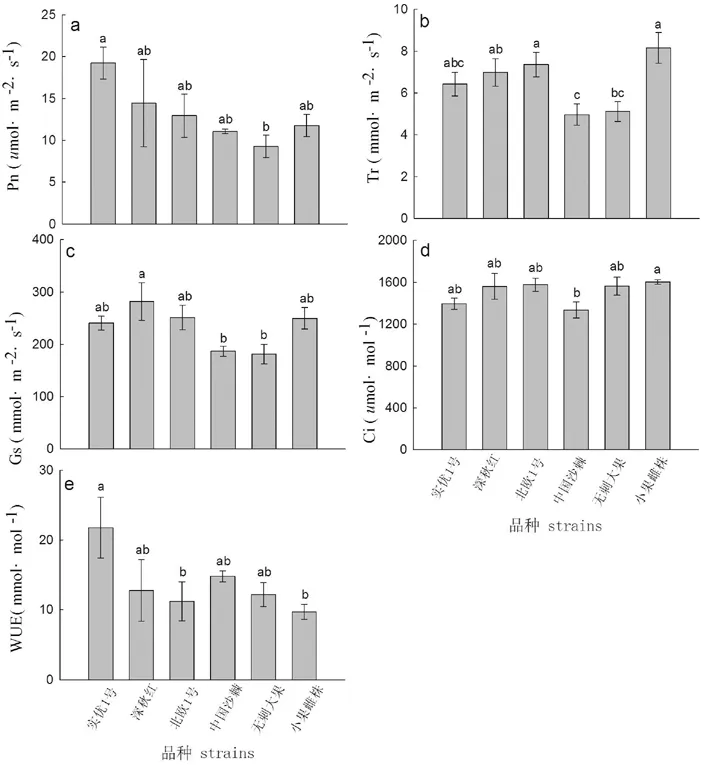
图2 不同品种沙棘光合生理参数日累积量的差异性
2.3 光合作用参数间相关性
Pn与Gs、WUE呈极显著正相关(分别为r=0.400;r=0.639)(P<0.01)但与Ci呈极显著负相关(r=-0.369,P<0.01)。Tr与Gs、Ci呈极显著正相关(分别为r=0.833;r=0.434)(P<0.01),而与WUE呈极显著负相关(r=-0.463,P<0.01)。Gs与Ci呈极显著正相关(r=0.403,P<0.01),与WUE呈显著负相关(r=-0.222,P<0.05)。Ci与WUE呈极显著负相关(r=-0.536,P<0.01)。
2.4 聚类分析
6种沙棘植物聚为两类(图3),其中‘实优1号’‘北欧1号’与‘小果雌株’这3个品种为一类,其光合生理较相近,且光合性能较优;而‘深秋红’‘无刺大果’及‘中国沙棘’聚为另一类,这三个品种的光合生理特性较相近,但在大田表现一般。
3 讨论
不同植物在相同生境下其结构和光合生理特征上常会出现一些差异[22],光合作用是植物感知外界环境变化的一项重要生理过程,众多学者已将光合作用作为评判植物是否适应环境的关键指标之一[23],同时植物的光合作用研究也可作为挖掘优良品种的重要依据[22]。因此,研究不同沙棘品种的光合作用属性不仅能判定沙棘的适生环境,而且有助于筛选优良的沙棘品种,并为沙棘系列产品的开发提供理论依据。
3.1 光合参数日变化特征
植物叶片生长是植物生长的基础,叶片还是光合作用的主要场所,植物在生长时叶片会呈现阶段性的特征变化,而这种变化会直接影响光合能力及光合效率[24]。净光合速率为评判光合作用的强弱的重要指标,净光合速率的大小受蒸腾速率、气孔导度、胞间CO2浓度、水分利用效率等多种因素影响,它们保障植物光合作用能够顺利进行[25],一般木本植物向阳生叶片的最大净光合速率为10~25 μmol/(m2·s)[3]。本研究中6个沙棘品种的净光合速率均较低,范围为0.9165 μmol/(m2·s)~10.437 μmol/(m2·s),与魏晓慧等[3]研究的沙棘的净光合速率较低一致,这可能受到沙棘叶片属性的影响。本研究中除‘无刺大果’在10∶00~12∶00出现峰值外,其余品种均在8∶00~10∶00出现了最高值,此时出现最高值的原因是早晨CO2浓度较高,空气中CO2在沙棘叶肉细胞扩散速率较快,同时此时的气孔导度值与各个时间段相比几乎为最大,气孔阻力也随之变小,而净光合速率达到最高值,在16∶00时又将至低谷,说明光强与温度不再是净光合速率的限制因素,而植物自身的呼吸速率起着关键作用,这与乔雨宁[26]等研究的黄土丘陵区5种乔灌木树种的光合生理特征一致,但沙棘净光合速率的变化趋势与陈海玲等[1]研究结果不太一致,可能是不同品种的光合反应及环境条件差异所致,而‘实优1号’与‘深秋红’在相同的生境下表现出较大的净光合速率,说明它们对光能的利用效率较高。
植物的蒸腾速率经常用来判断其水分散失情况[25],但蒸腾速率过高时植物极易水分失调,对其生长将造成阻碍[27];相反,在同样的环境下蒸腾速率越小说明植物抗旱能力越优,沙棘作为喜光植物,本身就具有极强的抗旱能力,在本研究中得知中国沙棘与‘无刺大果’的抗旱力优于其他品种,这与鲁客[28]研究的不同品种沙棘抗旱性评价相一致。‘北欧1号’、‘无刺大果’及‘小果雌株’均在10∶00~12∶00出现峰值,原因是正午温度高,植物因自身生理作用自动关闭气孔,从而蒸腾力降低;‘实生1号’与‘深秋红’的在12∶00~14∶00处于峰值,而中国沙棘的峰值在14∶00~16∶00,这是因为不同沙棘品种对当地环境适应有很大不同[28]。气孔导度可直接反映植物叶片的气体交换能力,同时它与蒸腾作用和胞间CO2浓度有密切关系[29],不仅如此经前人研究气孔导度是探究植物叶片—水汽热稳定与碳同化的生理指标[30]。同上气孔导度、胞间CO2浓度与蒸腾速率的日变化趋势相似,说明它们之间关系密切。CO2为植物光合作用的原料之一,胞间CO2浓度能够反映光合能力的强弱[31],而水分利用效率由植物的净光合速率和蒸腾速率来决定,一般将其作为评价植物在该区适生与否的重要指标[3]。在本研究中‘小果雌株’的蒸腾速率最大,而水分利用效率最低;中国沙棘的蒸腾速率最小,而水分利用效率最高;说明‘小果雌株’的抗旱能力差,一方面说明其传导组织发达,生理活动过旺,另外说明该品种在生长发育过程中需消耗更多的土壤水分,因此在旱季栽培时需频繁的灌溉以保证其生长状况可正常运行。
3.2 日累积量的差异性
分析光合生理日累积量的差异能够揭示沙棘在毛乌素沙地的生长潜力及应用推广价值[26]。本研究中‘实优1号’净光合速率日累积量远高于其他品种,桑林等[14]从净光合速率来决定矾根(Heuchera micrantha)的引种,净光合速率越强说明植物对高温和强光照的适应力更强。‘深秋红’的气孔导度日累积量在各品系间达到了最高值,说明其呼吸作用与蒸腾作用较优,而‘无刺大果’的气孔导度日累积量最小,因此呼吸及蒸腾作用较小。另外,‘实优1号’与‘中国沙棘’的水分利用效率日累积量较大,说明该品种与其他品种相比抗旱能力较差,在栽培过程中应适时灌溉,保证其正常的生命活动;而‘小果雌株’与‘北欧1号’抗旱性较佳,可在干旱地区进行指导栽培。
3.3 光合参数间相关性
本研究中,沙棘的净光合速率主要受蒸腾速率、气孔导度、胞间CO2浓度、水分利用效率的影响,其中净光合速率与胞间CO2浓度呈极显著负相关,说明净光合速率会受到高CO2浓度的抑制;蒸腾速率、气孔导度、胞间CO2浓度都呈极显著正相关,说明气孔导度越大,参与光合作用的CO2越多,胞间CO2浓度增大,净光合速率也增大。有学者研究沙棘光合参数与光合有效辐射间关系时发现沙棘气孔导度下降会引起其光合速率与蒸腾速率下降,但当蒸腾速率下降速度大于光合速率时,水分利用效率会升高[32],这与本研究相一致,但因各学者所取试验地、实验材料以及研究指标兼有不同,因此还得后续进一步完善研究。此外,王宇等[33]发现蒸腾速率由光合有效辐射、大气温度(Ta)、大气湿度(RH)、叶面温度(Tleaf)、胞间CO2浓度、气孔阻力、水蒸气压亏缺(VPD)等多种因素影响,本研究针对蒸腾速率与另外4种光合参数的响应与此研究结果一致。
3.4 聚类分析
本研究中,‘实优1号’‘北欧1号’‘小果雌株’的光合性能优于‘深秋红’‘无刺大果’‘中国沙棘’,结合鲁客等[28]研究表明两年生‘深秋红’‘实优1号’‘北欧1号’在榆林地区适生性更强,但在其研究中无‘小果雌株’品种,因此本文与其研究结果较为一致,因此可以加大‘实优1号’‘北欧1号’‘小果雌株’这3个品种的种植面积。在5个沙棘苗木的光合生理指标中,高Pn对于评价沙棘苗木的光合生理特性起着决定性作用;但6个沙棘植株品种的光合生理特性是否相近除了这5个光合生理特性协同决定外,另外还受水蒸气压亏缺、光能利用效率(LUE)、气孔限制值(Ls)等光合指标以及大气湿度、土壤温度(Ts)与大气温度等多种环境因素影响,并不是仅仅由其中一两个光合指标来决定。
4 结论
6个品种的的Tr日变化趋势均呈现单峰型。‘实优1号’‘深秋红’‘北欧1号’‘中国沙棘’与‘小果雌株’的Pn均在8∶00~10∶00出现了最高值,而‘无刺大果’在10∶00~12∶00出现了峰值。‘实优1号’‘深秋红’‘中国沙棘’‘无刺大果’的Gs在8∶00~10∶00出现了最高值,而‘北欧1号’与‘小果雌株’在10∶00~12∶00出现峰值。‘实优1号’‘深秋红’‘北欧1号’及‘小果雌株’的Ci日变化趋势均呈现单峰型,中国沙棘与‘无刺大果’的最高值出现于8∶00~10∶00。除‘无刺大果’外,其余5个品种的最高值均出现于8∶00~10∶00;‘实优1号’的Pn与WUE日累积量高于其它品种;‘深秋红’的Gs呈现最大值;‘小果雌株’的Tr与Ci的日累积量高于其他品种; Pn与Gs、WUE;Tr与Gs、Ci;Gs与Ci呈极显著正相关;Pn与Ci;Tr与WUE;Ci与WUE呈极显著负相关;Gs与WUE呈显著负相关。6个一年生沙棘品种被分为两类,结合光合属性表明,‘实优1号’‘北欧1号’‘小果雌株’为较优品种,可作为主要的沙棘品种进行培育;而‘深秋红’‘无刺大果’‘中国沙棘’表现一般。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
花卉(2022年17期)2022-09-06
食品安全导刊(2021年21期)2021-08-30
花火B(2021年3期)2021-06-25
落叶果树(2021年6期)2021-02-12
启迪与智慧·下旬刊(2019年2期)2019-09-10
上海农业学报(2017年3期)2017-04-10
现代农村科技(2017年6期)2017-02-02
现代农业科技(2009年7期)2009-07-29
现代农业科技(2009年20期)2009-03-20
