
碱度驯化对大鳞鲃幼鱼血液生理生化及肝脏抗氧化系统的影响*
2022-09-05 01:51李海涛赵志刚李明帅刘金雨耿龙武杜汝君丛艳峰
渔业科学进展 2022年4期
李海涛 徐 伟 赵志刚 罗 亮 李明帅,3 刘金雨 王 雨 张 瑞 郭 坤 耿龙武 杜汝君 丛艳峰
碱度驯化对大鳞鲃幼鱼血液生理生化及肝脏抗氧化系统的影响*
李海涛1,2徐 伟2①赵志刚2①罗 亮2李明帅2,3刘金雨1,2王 雨1,2张 瑞2郭 坤2耿龙武2杜汝君4丛艳峰4
(1. 上海海洋大学水产与生命学院 上海 201306;2. 中国水产科学研究院黑龙江水产研究所 黑龙江省冷水性鱼类种质资源及增养殖重点开放实验室 黑龙江 哈尔滨 150070;3. 东北农业大学 黑龙江 哈尔滨 150030;4. 杜尔伯特蒙古族自治县水产总站 黑龙江 大庆 166200)
为了从血液生理生化、肝脏抗氧化应激等方面研究大鳞鲃()对碱度驯化的生理适应性变化,选择体重为(13.66±1.26) g的大鳞鲃幼鱼开展NaHCO3碱度适应性驯化实验,空白组一直处于淡水中养殖,驯化组经20 mmol/L的碱度适应性驯养7 d后再放入40 mmol/L的碱度水体中,未驯化组直接放入40 mmol/L的碱度水体中,测定并比较了鱼体放入40 mmol/L碱度水体中第0 h、6 h、12 h、24 h、48 h、96 h、7 d幼鱼血液生理生化指标和肝组织抗氧化系统相关指标变化。结果显示,驯化组和未驯化组鱼体的血液渗透压、白细胞、淋巴细胞、中性粒细胞、单核细胞、血红蛋白、红细胞、血小板、尿素、白蛋白含量和血小板压积等生理生化指标以及肝组织抗氧化系统中的超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GSH-Px)和丙二醛(MDA)在0 h~7 d的碱度胁迫过程中,均表现为先升高后降低的变化趋势(<0.05),且驯化组峰值大小均显著性低于未驯化组(<0.05),空白组在此期间均未表现出显著性变化(>0.05)。驯化组鱼的血常规指标参数和肝脏组织的SOD、CAT出现峰值的时间均晚于未驯化组。驯化组鱼体在高碱度胁迫第7天时,其血液中的尿素浓度、淋巴细胞、单核细胞、血红蛋白、红细胞、血小板、白蛋白含量、谷草转氨酶(AST)、谷丙转氨酶(ALT)以及肝组织中的SOD、CAT、GSH-Px、MDA参数均显著性低于未驯化组(<0.05)。研究表明,大鳞鲃幼鱼经过一定程度的碱度驯化后,在遭受更高碱度的水环境胁迫时,从生理层面反映出机体具有更强的适应性。
大鳞鲃;碱度;生理生化;抗氧化酶
我国内陆地区约有0.64亿hm2的低洼盐碱水域资源,约占全国湖泊总面积的55% (武鹏飞等, 2017),盐碱水具有高盐、高碱、高pH、离子组成复杂等特点。大多数淡水鱼类无法在这样的水环境中生存或生长(李生荣, 2009),为充分开发利用盐碱水资源,科研人员陆续移植和引进了大鳞鲃()、咸海卡拉白()、雅罗鱼()和罗非鱼()等耐盐碱鱼类开展养殖(常玉梅等, 2021)。耐盐碱鱼类在养殖早期,尤其在鱼苗培育阶段,由于其身体结构和生理机能尚未发育完全,对盐碱水环境的适应性较差,易因应激反应导致鱼体的成活率较低(付占斐等, 2020)。王妤等(2011)研究表明,在盐碱水域养殖过程中,经过适当驯化可以有效提高鱼体的成活率,同时,其生长、抗氧化和免疫性能等方面也均有所提高。杨 静雯等(2021)研究表明,可通过每天2 g/L的升盐驯化方式使虹鳟()逐渐适应海水,驯化后的超氧化物歧化酶(SOD)活性、谷胱甘肽酶(GSH-Px)活性和丙二醛(MDA)含量与对照组相比无显著差异。成智丽等(2021)研究表明,每7 d降低1 g/L的慢性盐度驯化过程中,许氏平鲉()的谷草转氨酶(AST)、谷丙转氨酶(ALT)升高,而血液渗透压、尿素浓度、白蛋白浓度降低。 秦少宗等(2021)研究表明,以每天1 g/L的盐度速率对中华鲟()幼鱼进行海水盐度驯化过程中,随着盐度的升高,海水组中华鲟幼鱼血清渗透压和主要离子(Na+和Cl–)均出现先升高后保持稳定的现象。但关于碱度驯化的相关研究还未见报道。
大鳞鲃隶属于鲤科(Cyprinidae)、鲃亚科(Barbinae)、鲃属,主要分布于西亚的里海和咸海水域,成鱼体重可达5~10 kg,是当地名贵的大型经济鱼类。该鱼具有盐碱耐受性强、食性广、生长速度快、肉质鲜美、抗逆性强等优良养殖特性(徐伟等, 2012a)。2003年从乌兹别克斯坦引入至今,我国已在大鳞鲃的人工繁育技术(徐伟等, 2011; 耿龙武等, 2010)、养殖生物学(蔺玉华等, 2009)、养殖技术(徐伟等, 2012b)等方面开展了相关研究。党云飞等(2013)研究表明,当碱度达到39.80 mmol/L时对幼鱼生长有抑制作用,大鳞鲃幼鱼的总摄食量和饵料转化效率开始急剧下降,且幼鱼最适生长碱度为20.57 mmol/L。大鳞鲃在盐碱胁迫初期,肝、肾和鳃中的抗氧化酶活力、MDA含量及血清皮质醇含量均与盐度呈显著正相关性(张宇婷等, 2021)。但关于大鳞鲃在盐碱方面的研究都是单纯的直接放入盐碱水体中以急性胁迫的方式来研究机体适应机制。本研究则是从血液生理生化、抗氧化应激等方面研究大鳞鲃幼鱼经过碱度驯化后与未驯化的生理适应性变化,旨在探究大鳞鲃幼鱼对碱度环境变化的适应性机制,并为大鳞鲃在盐碱水域养殖技术的建立提供理论依据。
1 材料与方法
1.1 实验材料
大鳞鲃幼鱼取自中国水产科学研究院黑龙江水产研究所呼兰实验基地,体长为(15.09±3.54) cm,体重为(13.66±1.26) g。实验开始前,鱼体在室内循环水玻璃缸内暂养7 d,暂养用水为曝气自来水。暂养期间每天换水1/3,暂养过程中溶解氧(DO)保持在5 mg/L以上,pH为7.4,水温控制在(22.0±1.5)℃。每天07:30左右饱食投喂1次,2 h后将残饵和粪便吸出。不同碱度的实验用水采用曝气2 d的自来水和NaHCO3(国产分析纯)进行配制,并用酸碱滴定法进行测定和校准。
1.2 实验方法
实验分别设置驯化组、未驯化组和空白组,每组设置3个平行,每个平行挑选规格一致的80尾健康大鳞鲃幼鱼。驯化组:先将暂养后的大鳞鲃置于20 mmol/L碱度水体中驯化7 d后,然后将水体碱度增加到40 mmol/L;未驯化组:大鳞鲃在淡水中养殖7 d后,直接将水体碱度调整到40 mmol/L;空白组:大鳞鲃一直淡水中养殖。驯化组和未驯化组分别将鱼体置于40 mmol/L碱度水体后的0、6、12、24、48、96 h以及7 d时的7个时间点,随机收集10尾鱼,经过40 mg/L MS-222麻醉后,对其进行血液和肝脏组织样品的采集。采集的肝脏组织样品立刻用液氮保存。采用2 mL规格的注射器,并用肝素钠抗凝在尾静脉采集血液进行多尾混样处理。空白组在对应的时间内也进行相同操作的采样过程。各组在驯化期间的DO、水温投喂频率、换水情况和暂养期间一致,并于采样前24 h停止投喂,且采样期间不进行换水处理。
1.3 样品测定
将抽出每个平行组的血液分为2份,1份新鲜血液采用动物血液细胞分析仪(BC-2800Vet, Mindray)测定其红细胞、白细胞、血小板、淋巴细胞含量等血常规指标;另1份血液静置2 h后,以2500 r/min离心10 min,取上清液,用生化分析仪(BS-240Vet, Mindray)及其迈瑞生物医疗电子有限公司生产的天门冬氨酸氨基转移酶(AST)测定试剂盒(IFCC法)、丙氨酸氨基转移酶(ALT)测定试剂盒、尿素(UREA)测定试剂盒(紫外-谷氨酸脱氢酶法)、葡萄糖(Glu)测定试剂盒(葡萄糖氧化酶法)、白蛋白(ALB)测定试剂盒(溴甲酚绿法)对血浆进行测定。血浆渗透压采用冰点下降法,用冰点渗透压计(Osmomat)取50 μL血浆进行测定(mOsm/kg)。肝脏样品中的SOD、过氧化氢酶(CAT)、谷胱甘肽酶(GSH-Px)和MDA均采用南京建成所生产的试剂盒进行测定。
1.4 数据统计
采用SPSS 20.0进行单因素方差分析(one-way ANOVA),并对组间或组内不同时期的显著性差异进行Duncan多重比较,<0.05为差异显著,数据结果均以平均值±标准差(Mean±SD)表示。采用独立样本的非参数检验进行驯化组和未驯化组峰值大小的显著性差异比较(<0.05)。使用GraphPad PrisM 5.0软件进行作图。
2 结果
2.1 血常规分析
大鳞鲃驯化组和未驯化组血液中白细胞、淋巴细胞、中性粒细胞和单核细胞含量均呈先升高后降低的变化趋势(<0.05)(图1)。驯化组鱼体各项参数的峰值出现在48 h,未驯化组的峰值出现时间在24 h,在24 h时,驯化组的各项指标均显著低于未驯化组,但在48 h时则相反(<0.05)。在碱度胁迫第7天时,驯化组鱼体血液中的淋巴细胞和单核细胞含量均显著低于未驯化组,但仍显著高于空白组(<0.05)。
在7 d的碱度胁迫过程中,驯化组和未驯化组鱼体的血小板、红细胞、血红蛋白含量和血小板压积均表现为先升高后降低的变化趋势(<0.05)(图2)。驯化组和未驯化组各参数的峰值出现在24 h和12 h,驯化组各参数的峰值大小均显著低于未驯化组(<0.05)。驯化组鱼体各项参数在胁迫0 h时均显著高于未驯化组(<0.05),而在胁迫第7天时则显著低于未驯化组(<0.05)。
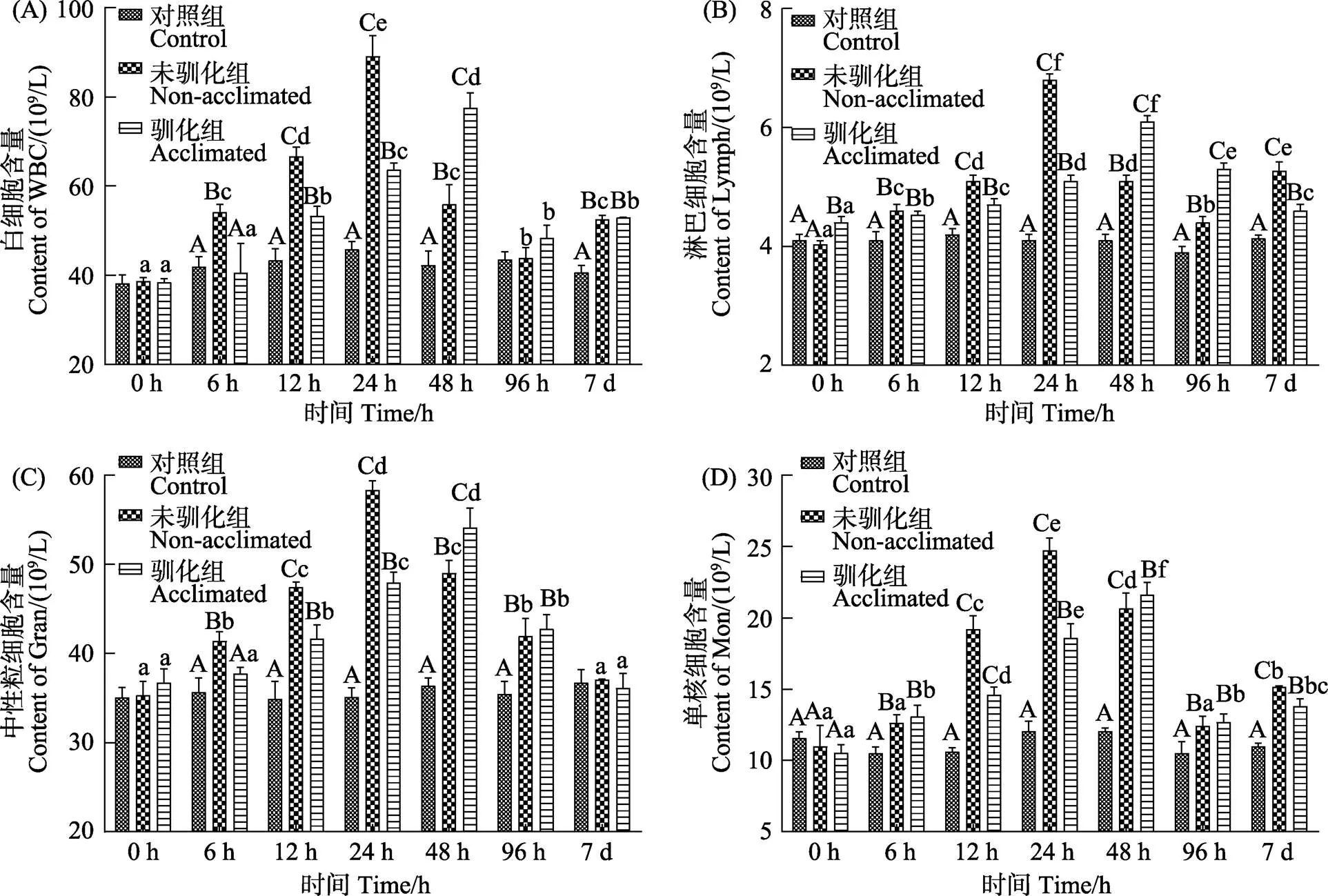
图1 碱度驯化对大鳞鲃幼鱼血液中的白细胞、淋巴细胞、中性粒细胞和单核细胞含量的影响
不同大写字母表示同时期不同处理组的显著差异(<0.05),不同小写字母代表同一处理组不同时期的显著差异(<0.05)。下同。
Different capital letters represent significant differences in different treatment groups at the same period (<0.05), and different small letters represent significant differences in different periods of the same treatment group (<0.05). The same as below.
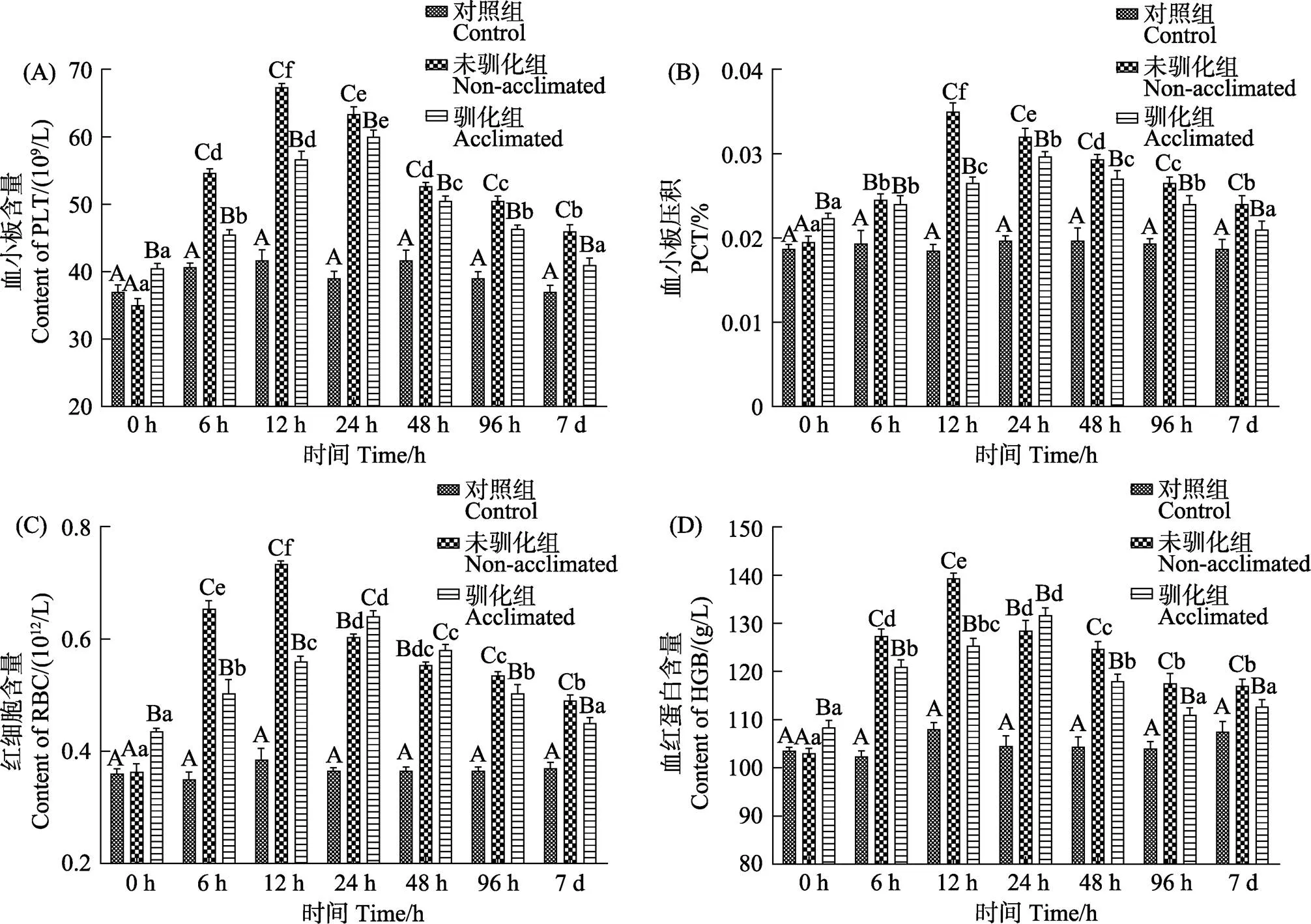
图2 碱度驯化对大鳞鲃幼鱼血液中的血红蛋白、红细胞、血小板含量和血小板压积的影响
2.2 血浆生理生化指标
驯化组和未驯化组鱼体血浆中的白蛋白含量、血糖浓度、尿素浓度、血浆渗透压均呈现先升高后降低的变化趋势(<0.05)(图3)。在6 h~7 d的各个取样时间点,驯化组鱼体的白蛋白含量均显著小于未驯化组(<0.05)。驯化组和未驯化组鱼体血浆白蛋白和血糖含量均在48 h时出现峰值,但驯化组的峰值显著小于未驯化组(<0.05)。驯化组在24 h时的血浆渗透压和尿素浓度峰值均显著小于未驯化组(24 h) (<0.05)。驯化组鱼体血浆渗透压在48 h后开始趋于稳定,而未驯化组在96 h后才开始逐渐降低。第7天时,驯化组鱼体的血浆白蛋白含量显著小于未驯化组(<0.05),且与空白组相比无显著性差异(>0.05)。在胁迫第7天时,驯化组鱼体的血浆尿素浓度显著小于未驯化组(<0.05),而其血浆渗透压在3组之间无显著差异(>0.05)。
在0 h~7 d内,驯化组和未驯化组鱼体的AST和ALT均呈现先升高后降低的变化趋势(<0.05),除0 h外,驯化组的血浆AST和ALT值均显著小于同时期未驯化组(<0.05)(图4),但仍显著高于空白组(<0.05)。驯化组鱼体在胁迫0 h和7 d时,血浆AST和ALT值均无显著性差异(>0.05)。
2.3 肝脏抗氧化酶系统
驯化组和未驯化组鱼体肝脏的SOD、CAT、GSH-Px、MDA在7 d的碱度胁迫范围内均呈先升高后降低的变化趋势(<0.05),空白组则无显著性变化(>0.05)(图5)。驯化组鱼体在胁迫24 h时的SOD和CAT峰值均显著小于未驯化组在12 h出现的峰值(<0.05)。胁迫第7天时,驯化组的SOD和CAT值均显著低于未驯化组(<0.05),但仍显著高于空白组(<0.05)。驯化组鱼体在96 h出现的GSH-Px峰值和48 h出现的MDA峰值均显著小于未驯化组(<0.05),胁迫7 d时,驯化组GSH-Px和MDA值显著低于未驯化组(<0.05),与空白组相比,驯化组的GSH-Px未表现出显著性差异(>0.05)。
3 讨论
3.1 碱度驯化对大鳞鲃血液生理的影响
当鱼体处于应激状态时,某些血液指标的变化可在一定程度上反映机体的基本状况(张宇婷, 2020)。本研究结果显示,驯化组和未驯化组的大鳞鲃幼鱼血液中白细胞、淋巴细胞、中性粒细胞和单核细胞的含量随着碱度胁迫时间的延长呈先升高后降低的变化趋势。Ashaf-Ud-Doulah等(2019)认为,当鱼体遭受环境因子胁迫时可通过增加自身的免疫反应来应对环境变化。本实验驯化组的大鳞鲃血液白细胞、淋巴细胞、中性粒细胞和单核细胞含量的上升幅度均显著小于未驯化组,表明大鳞鲃在经过低碱度水体驯化后,再遭遇较高碱度水体时,机体的应激反应较小。且在应激第7天时,虽然驯化组大鳞鲃血液中的淋巴细胞和单核细胞含量均显著大于空白组,但显著低于未驯化组,表明经驯化后的鱼体在遭遇较高碱度胁迫后,其机体生理恢复能力较强。
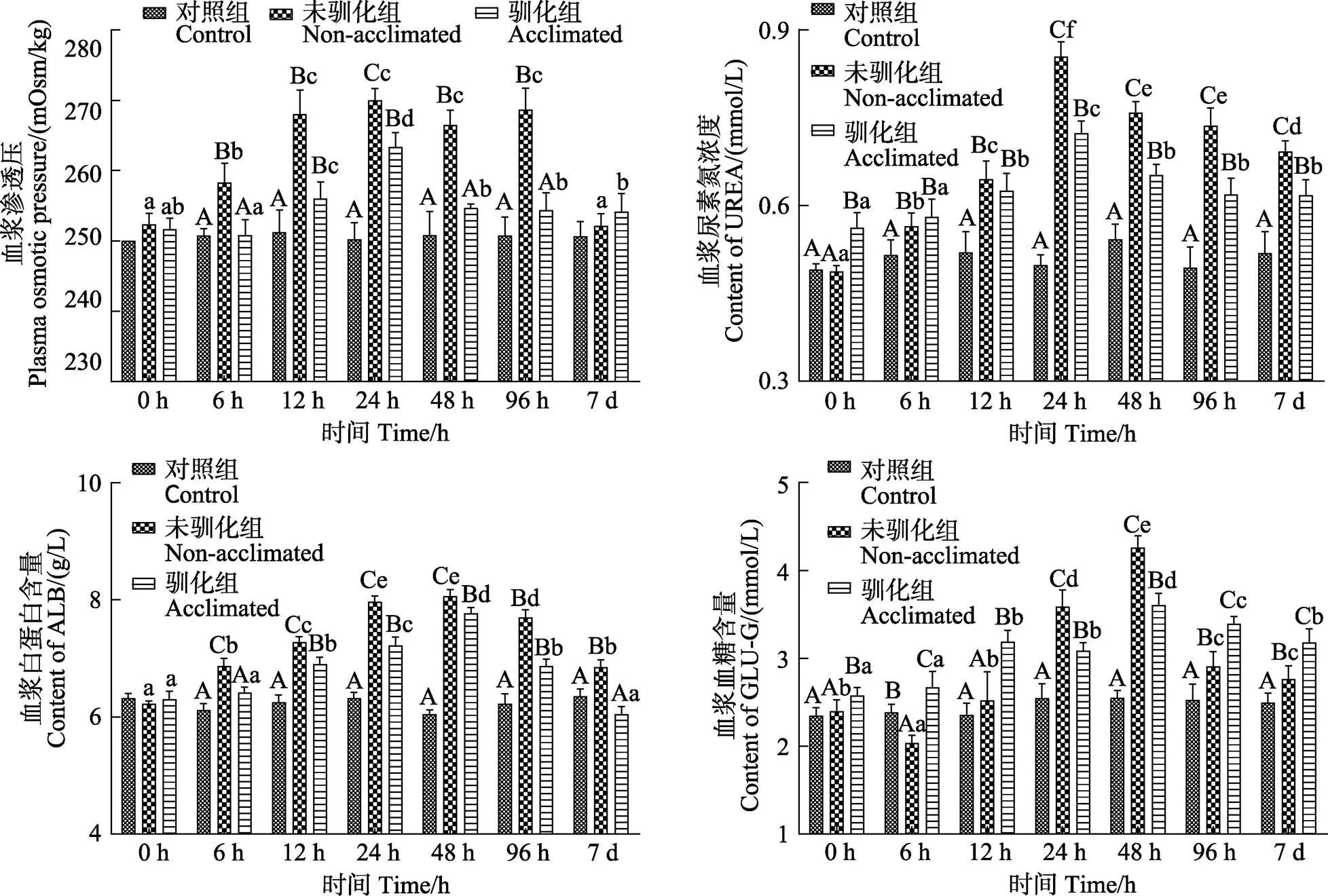
图3 碱度驯化对大鳞鲃幼鱼的血浆渗透压、尿素浓度、血糖浓度和血浆白蛋白含量的影响
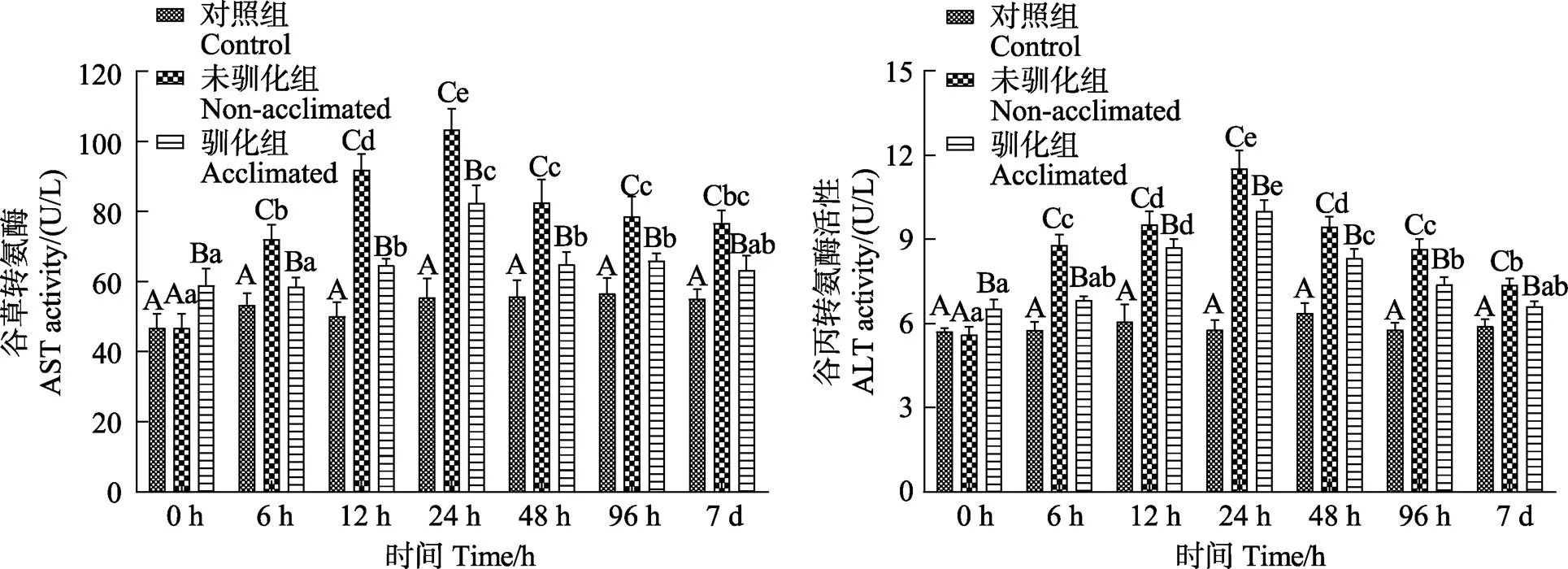
图4 碱度驯化对大鳞鲃幼鱼的血浆AST和ALT的影响
血小板在鱼体血液中起着凝血止血、修补破损血管的功能(王龙涛等, 2019)。本研究中,驯化组的大鳞鲃幼鱼急性碱度胁迫下血小板出现峰值时间(24 h)晚于非驯化组(12 h),且变化幅度小于非驯化组;同时,驯化组鱼体的血小板含量和血小板压积在胁迫第7天时显著性低于非驯化组,表明大鳞鲃在经低浓度碱度驯化后,机体应对高碱度环境时其组织的血管损伤度有所降低。机体血液红细胞中含有的血红蛋白具有运输氧的功能,血液中红细胞和血红蛋白数量的增加意味着血液对氧运输能力增强,鱼体的代谢能力提高(赵海涛等, 2006)。本研究中,虽然驯化组鱼体在应激0 h时的红细胞、血红蛋白含量均显著性高于未驯化组和空白组,这也有可能是前期驯化导致的,但从血红蛋白、红细胞含量的峰值大小和峰值出现时间来看,驯化组鱼体在经历较高浓度的碱度胁迫时表现出更小的应激反应。有研究表明,碱度胁迫下鱼体的耗氧率增加(姚娜等, 2016; 耿龙武等, 2017),驯化组鱼体在应激第7天时,其血液红细胞含量、血红蛋白含量均显著低于未驯化组,表明与未驯化的大鳞鲃相比,驯化后的鱼体在经历更高碱度胁迫时,需要消耗更少的能量来维持机体的代谢。
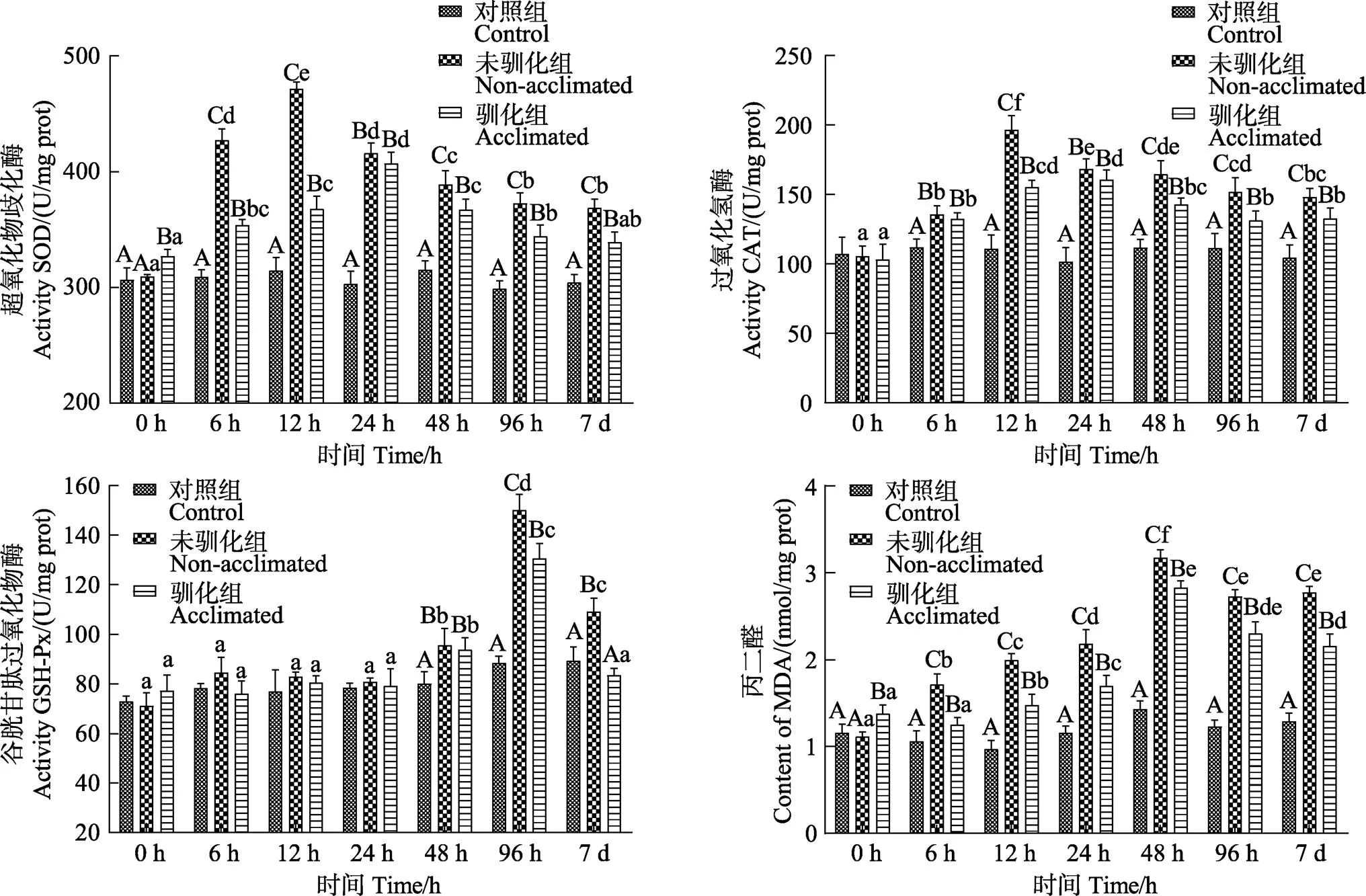
图5 碱度驯化对大鳞鲃幼鱼SOD、CAT、GSH-Px、MDA的影响
3.2 碱度驯化对大鳞鲃血浆生理生化的影响
鱼类生命活动最直接的能量来源于葡萄糖,鱼体血糖水平的变化与水环境胁迫因子在某种程度上有着密不可分的联系(Wagner, 2004)。洪磊等(2004)研究指出,环境胁迫会引起鱼类的内分泌变化,如儿茶酚胺和皮质类固醇的升高,随之血液中葡萄糖和乳酸含量增加。本研究中,驯化组和未驯化组鱼体的血糖浓度均高于空白组,这与王龙涛等(2019)对异育银鲫()的研究结果一致。王瑞芳等(2019)对瓦氏雅罗鱼()的研究中也同样发现,在盐碱湖泊中的鱼体糖原异生作用增强,且血糖浓度高于淡水瓦氏雅罗鱼。本研究中,驯化组大鳞鲃在应激第7天时的血糖浓度显著高于未驯化组,其原因可能是经碱度驯化后的大鳞鲃幼鱼耗氧率有所降低(耿龙武等, 2017),导致其能量代谢水平降低,血糖浓度维持在较高水平。机体耗氧率降低,在一定程度上有利于鱼体生长(徐钢春等, 2012)。驯化组在应激第7天时的红细胞含量和血红蛋白浓度显著低于未驯化组,也间接说明了经驯化后大鳞鲃的耗氧率有所降低。
鱼类通过机体的渗透压调节来适应水环境的变化,当机体渗透压超过一定限度后会影响鱼体的生长发育,严重时会导致鱼体死亡(沈双烨等, 2013)。本研究结果显示,经碱度驯化后的大鳞鲃幼鱼血浆渗透压在应对40 mmol/L的碱度胁迫时其应激反应较低,应激胁迫后渗透压恢复较快。主要原因是经过驯化后的大鳞鲃在应对更高碱度胁迫时,机体已经提前适应低碱度变化而进入主动调节阶段,当鱼体再次应对碱度变化时,相对于未驯化组并未表现出更大的应激反应(Partridge, 2002)。黄国强等(2013)对褐牙鲆()幼鱼的研究中也发现,每日的盐度变化幅度越小,幼鱼血清渗透压达到稳定所需时间越短。驯化组和未驯化组大鳞鲃渗透压在碱度胁迫第7天时,均能恢复至空白组水平,表明大鳞鲃幼鱼能够通过自身调节渗透压来应对40 mmol/L碱度的变化,这与耿龙武等(2011)的研究结果一致。耿龙武等(2011)研究表明,鱼体血液中尿素含量增加可以在碱性水境中作为其他离子的补充调节渗透压,可在一定程度上增强鱼体在高碱水域中的渗透调节能力,从而提高鱼体对高碱水环境的适应性。本研究驯化组鱼体血浆尿素含量的出现峰值早于未驯化组,且在应激第7天时,其血浆尿素含量显著低于未驯化组,这些结果均表明,驯化组鱼体因在一定程度上适应了碱度变化而导致其血液尿素含量变化较小。同时,驯化后的鱼体此时可能通过调节自身代谢降低氨的产生进而适应高碱环境。
正常情况下,鱼体肝脏组织中的转氨酶只有少量被释放到血浆中,当鱼体受到应激或组织病变时,会引起组织细胞膜通透性增加,细胞内的转氨酶就会大量释放出来进入血液,引起血清中AST和ALT活性的异常升高(黄晓荣等, 2007)。本研究中,驯化组和未驯化组大鳞鲃血浆AST和ALT活性在实验过程中均显著高于空白组,说明2组鱼体在此期间均受到了碱度胁迫。驯化组鱼体中2种酶的活性在各时间点均显著低于非驯化组,表明经驯化后的大鳞鲃一定程度上适应了碱度胁迫,避免了在高碱度环境下鱼体组织受到伤害。值得注意是,驯化组鱼体的AST和ALT在应激0 h和7 d的2个时间点均无显著性差异,表明驯化后的大鳞鲃幼鱼在面对40 mmol/L的NaHCO3碱度变化时,能够恢复至驯化后胁迫前的水平,相较于未驯化组机体的损伤程度较低。
3.3 碱度驯化对大鳞鲃肝脏抗氧化系统的影响
SOD、CAT和GSH-Px等抗氧化酶在生物体的抗氧化防御机制中有着清除活性氧分子、增强吞噬细胞防御能力和机体免疫能力的作用,而MDA含量的高低又间接反映了机体细胞受自由基攻击的严重程度(张宇婷等, 2021)。杨静雯等(2021)研究表明,当水体盐度变化较快时,虹鳟和硬头鳟()因长期处于氧化应激状态且不断产生自由基,抗氧化酶为清除活性氧危害始终处于较高水平状态。本实验结果显示,与空白组相比,驯化组和未驯化组鱼体肝脏的SOD、CAT、GSH-Px酶活性和MDA含量均显著较高,表明鱼体在面对高碱度变化时机体受到了较大程度的应激,肝组织在一定程度上受损,需要较高的SOD、CAT、GSH-Px酶活来清除体内过多的自由基。驯化组鱼体在应激第7天时,各种酶活性均显著小于未驯化组,且驯化组各种酶的峰值也均显著性小于未驯化组,表明经碱度驯化后的大鳞鲃在遭受更高碱度的胁迫时具有更强的适应性。
4 结论
本文研究了碱度适应性驯化对大鳞鲃幼鱼血液生理生化及肝脏抗氧化系统的影响,研究表明,大鳞鲃幼鱼经过一定程度的碱度驯化后,在遭受更高碱度的水环境胁迫时,从生理层面反映出机体具有更强的适应性。
ASHAF-UD-DOULAH M, SHAHJAHAN M, ISLAM S M M,Thermal stress causes nuclear and cellular abnormalities of peripheral erythrocytes in Indian major carp, rohu. Journal of Thermal Biology, 2019, 86: 102450
CHANG Y M, LIANG L Q.Advances of research of physiological and molecular mechanisms related to alkali-saline adaptation for fish species inhabiting alkali-saline water. Journal of Fisheries of China, 2021, 45(5): 798–812 [常玉梅, 梁利群. 耐盐碱鱼类的生理和分子机制研究进展. 水产学报, 2021, 45(5): 798–812]
CHENG Z L, SUN P F, LUO J,. Changes in serum bochemical profiles and osmotic pressure of Schlegel’s black rockfishacclimatized by various salinities. Fisheries Science, 2021, 40(3): 310–317 [成智丽, 孙鹏飞, 罗珺, 等. 盐度驯化下许氏平鲉血清生化指标及渗透压的变化. 水产科学, 2021, 40(3): 310–317]
DANG Y F, XU W, GENG L W,. Effects of NaCl and NaHCO3on growth and gill microstructure in juvenile. Journal of Fishery Sciences of China, 2013(3): 577–584 [党云飞, 徐伟, 耿龙武, 等. NaCl盐度和NaHCO3碱度对大鳞鲃幼鱼生长及鳃组织特征的影响. 中国水产科学, 2013, 20(3): 577–584]
FU Z F, WANG X, WANG F,Effect of acclimation at different salinities on survival and growth of rainbow trout and steelhead trout juveniles. Periodical of Ocean University of China (Natural Science), 2020, 50(7): 37–43 [付占斐, 王鑫, 王芳, 等. 不同盐度驯化方式对虹鳟和硬头鳟幼鱼存活和生长的影响. 中国海洋大学学报(自然科学版), 2020, 50(7): 37–43]
GENG L W, JIANG H F, XU W,Effects of carbonate alkalinity on oxygen consumption and ammonia excretion in bulatmai barbel. Chinese Journal of Fisheries, 2017, 30(6): 30–33 [耿龙武, 姜海峰, 徐伟. 碳酸氢钠对大鳞鲃耗氧率和排氨率的影响. 水产学杂志, 2017, 30(6): 30–33]
GENG L W, XU W, LI C T,Influence of salinity and alkalinity on serum ion concentrations, osmosis and gill Na+/K+-ATPase activity of. Journal of Fishery Sciences of China, 2011, 18(2): 458–465 [耿龙武, 徐伟, 李池陶, 等. 盐碱对大鳞鲃血清渗透压、离子含量及鳃丝Na+/K+-ATP酶活力的影响. 中国水产科学, 2011, 18(2): 458–465]
GENG L W, XU W, LIN Y H,Technique of artificial reproduction of. Journal of Jilin Agricultural University, 2010, 32(2): 218–220 [耿龙武, 徐伟, 蔺玉华, 等. 大鳞鲃人工繁育技术初报. 吉林农业大学学报, 2010, 32(2): 218–220]
HONG L, ZHANG X M. Effects of environmental stress on physiological function of fish. Advances in Marine Science, 2004, 22(1): 114–121 [洪磊, 张秀梅. 环境胁迫对鱼类生理机能的影响. 海洋科学进展, 2004, 22(1): 114– 121]
HUANG G Q, ZHANG L Y, LI J,Effects of salinity acclimatization on the osmoregulation and energy metabolism of brown flounder (). Periodical of Ocean University of China (Natural Science), 2013, 43(5): 44–51 [黄国强, 张灵燕, 李洁, 等. 盐度驯化对褐牙鲆幼鱼渗透压调节和能量代谢的影响. 中国海洋大学学报(自然科学版), 2013, 43(5): 44–51]
HUANG X R, ZHANG L Z, ZHUANG P,Haematological biochemistry of Amur sturgeon,, at different domestication salinities. Journal of Shanghai Fisheries University, 2007, 16(2): 144–150 [黄晓荣, 章龙珍, 庄平, 等. 盐度驯化下施氏鲟的血液生化指标. 上海水产大学学报, 2007, 16(2): 144–150]
LI S R. The present situation and countermeasures of water resources in China—The shortage of water resources restricts the development of China's economy and society. Journal of Yan´an Vocational and Technical Institute, 2009, 23(6): 101–103, 109 [李生荣. 我国水资源的现状与对策—水资源短缺制约着我国经济社会的发展. 延安职业技术学院学报, 2009, 23(6): 101–103, 109]
LIN Y H, GENG L W, WANG X H,Some biological characteristics ofbred in ponds. Journal of Tianjin Normal University (Natural Science), 2009, 29(2): 72–75 [蔺玉华, 耿龙武, 王信海, 等. 池塘驯养大鳞鲃某些生物学特性研究. 天津师范大学学报(自然科学版), 2009, 29(2): 72–75]
PARTRIDGE G J, JENKINS G I. The effect of salinity on growth and survival of juvenile black bream (). Aquaculture, 2002, 210(1/2/3/4): 219–230
QIN S Z, LENG X Q, LUO J,. Adaptive adjustment of osmotic organ structure of juvenile Chinese sturgeon in seawater conditions. Progress in Fishery Sciences, 2021, 42(4): 82–89 [秦少宗, 冷小茜, 罗江, 等. 中华鲟幼鱼渗透调节器官组织结构在海水条件下的适应性调整. 渔业科学进展, 2021, 42(4): 82–89]
SHEN S Y, ZHANG S S, XIE H,Effects of salinity domestication on serum biochemistry and osmotic pressure of carp. Chinese Journal of Ecology, 2013, 32(12): 3261–3267 [沈双烨, 张赛赛, 谢唤, 等. 盐度驯化对鲤血清生化指标及渗透压的影响. 生态学杂志, 2013, 32(12): 3261–3267]
WAGNER T, CONGLETON J L. Blood chemistry correlates of nutritional condition, tissue damage, and stress in migrating juvenile Chinook salmon (). Canadian Journal of Fisheries and Aquatic Sciences, 2004, 61(7): 1066–1074
WANG L T, GE Z X, GAO C S,Effect of different concentrations of saline-alkali stress on the chronic injury of. Chinese Journal of Veterinary Medicine, 2019, 55(7): 84–87 [王龙涛, 葛晨霞, 高春山, 等. 不同浓度盐碱胁迫对异育银鲫的慢性损伤效应. 中国兽医杂志, 2019, 55(7): 84–87]
WANG R F, AN X P, QI J W,Energy metabolism and digestive enzyme activity regulation in Amur Ide () during spawning migration from Dali Nor Lake to Gongger River. Journal of Fishery Sciences of China, 2019, 26(4): 703–712 [王瑞芳, 安晓萍, 齐景伟, 等. 达里湖瓦氏雅罗鱼生殖洄游过程中能量代谢和消化酶活性的变化. 中国水产科学, 2019, 26(4): 703–712]
WANG Y, ZHUANG P, ZHANG L Z,. Effects of salinity on survival, growth and antioxidant defense system of. Journal of Fisheries of China, 2011, 35(1): 66–73 [王妤, 庄平, 章龙珍, 等. 盐度对点篮子鱼的存活、生长及抗氧化防御系统的影响. 水产学报, 2011, 35(1): 66–73]
WU P F, GENG L W, JIANG H F,Tolerance of three Cobitidae fish species to high salinity and alkalinity. Journal of Fishery Sciences of China, 2017, 24(2): 248–257 [武鹏飞, 耿龙武, 姜海峰, 等. 三种鳅科鱼对NaCl盐度和NaHCO3碱度的耐受能力. 中国水产科学, 2017, 24(2): 248–257]
XU G C, NIE Z J, BO Q K,Effects of water temperature on oxygen consumption rate, asphyxiant point, blood glucose content, and muscle and liver glycogen content of juvenile. Chinese Journal of Ecology, 2012, 31(12): 3116–3120 [徐钢春, 聂志娟, 薄其康, 等. 水温对刀鲚幼鱼耗氧率、窒息点、血糖及肌肝糖元指标的影响. 生态学杂志, 2012, 31(12): 3116–3120]
XU W, GENG L W, LI C T,The artificial propagation, embryonic development and saline-alkali tolerant experiment of. Journal of Fisheries of China, 2011, 35(2): 255–260 [徐伟, 耿龙武, 李池陶, 等. 大鳞鲃的人工繁殖、胚胎发育和耐盐碱测定. 水产学报, 2011, 35(2): 255–260]
XU W, GENG L W, LI C T,The growth and overwintering survival ofbred in pondin northern China. Freshwater Fisheries, 2012a, 42(1): 68–71 [徐伟, 耿龙武, 李池陶, 等. 北方寒地池养大鳞鲃生长和越冬成活. 淡水渔业, 2012a, 42(1): 68–71]
XU W, GENG L W, MIAO J S,. Current researches on saline-alkali tolerant fish of. Journal of Tianjin Agricultural University, 2012b, 19(3): 62–64 [徐伟, 耿龙武, 苗建生, 等. 耐盐碱鱼类大鳞鲃的研究现状. 天津农学院学报, 2012b, 19(3): 62–64]
YANG J W, YANG X G, HUANG M,. Effects of salinity change on antioxidant enzyme activities of rainbow and steelhead trout (). Periodical of Ocean University of China (Natural Science), 2021, 51(6): 26–33 [杨静雯, 杨小刚, 黄铭, 等. 盐度变化对虹鳟和硬头鳟抗氧化酶活性的影响. 中国海洋大学学报(自然科学版), 2021, 51(6): 26–33]
YAO N, CHEN S A, WANG S,Effects of salinity, alkali and weight on oxygen consumption and ammonia excretion rate of()(Day)Southwest China Journal of Agricultural Sciences, 2016, 29(2): 462–468 [姚娜, 陈生熬, 王帅, 等. 盐度、碱度和体质量对叶尔羌高原鳅耗氧率和排氮率的影响. 西南农业学报, 2016, 29(2): 462–468]
ZHANG Y T, YANG J, GENG L W,Effects of salinity stress on antioxidant enzymes and serum cortisol in. Progress in Fishery Sciences, 2021, 42(1): 56–62 [张宇婷, 杨建, 耿龙武, 等. 盐度胁迫对大鳞鲃抗氧化酶和血清皮质醇的影响. 渔业科学进展, 2021, 42(1): 56–62]
ZHANG Y T. Study of immunological reaction ofunder saline-alkali stress based on transcriptome sequencing technology. Master′s Thesis of Shanghai Ocean University, 2020 [张宇婷. 基于转录组技术的大鳞鲃盐碱胁迫免疫应激响应研究. 上海海洋大学硕士研究生学位论文, 2020]
ZHAO H T, ZHANG Q Z, ZHAO H P,Comparative study on hematological indices in larval and adult southern catfish. Chinese Journal of Zoology, 2006, 41(1): 94–99 [赵海涛, 张其中, 赵海鹏, 等. 南方鲇幼鱼和成鱼血液指标的比较. 动物学杂志, 2006, 41(1): 94–99]
Effects of Alkalinity Acclimation on Blood Physiology and Biochemistry and Liver Antioxidant System of Juvenile
LI Haitao1,2, XU Wei2① , ZHAO Zhigang2①, LUO Liang2, LI Mingshuai2,3, LIU Jinyu1,2, WANG Yu1,2, ZHANG Rui2, GUO Kun2, GENG Longwu2, DU Rujun4, CONG Yanfeng4
(1. Shanghai Ocean University, College of Fisheries and Life Science, Shanghai 201306, China;2. Heilongjiang Fisheries Research Institute, Chinese Academy of Fishery Sciences, Key Open Laboratory of Cold Water Fish Germplasm Resources and Breeding of Heilongjiang Province, Harbin, Heilongjiang 150070, China;3. Northeast Agricultural University, Harbin, Heilongjiang 150030, China;4. Aquatic Products Station of Dorbod Mongolian Autonomous County, Daqing, Heilongjiang 166200, China)
There are approximately 64 million hm2of low-lying saline-alkali water resources in inland China, accounting for 55% of the total lake area. The characteristics of saline-alkali water are: high salt content, high alkalinity (high pH), and complex ion composition. Most freshwater fish cannot survive or reproduce in these conditions. To utilize saline-alkali water resources for aquaculture, researchers have introduced salt-tolerant fish, such as,,, and. In the initial stage of aquaculture, fish fry are not yet adapted to saline-alkali conditions because their body structure and physiological functions are not underdeveloped. The survival rate of juvenile fish is low because of the stress response to saline-alkali water. Studies of saline-alkali aquaculture have shown that proper acclimation can effectively improve the survival rate, growth, antioxidant, and immune properties of fish.
belongs to the Cyprinidae family (subfamily Barbinae). It occurs mainly in the Caspian Sea and the Aral Sea of Western Asia.is an economically important fish, with adults weighing 5~ 10 kg. The species has excellent breeding characteristics, high saline-alkali tolerance, varied food habits, fast growth, delicious meat, and strong stress resistance. In 2003, it was introduced from Uzbekistan to China. To date, research has reported onartificial breeding technology and breeding biology but has not yet considered salt-alkali domestication.
To study the physiological adaptability ofto alkalinity with regards to blood physiology, biochemistry, and antioxidative stress, we used juvenile(13.66±1.26) g to test NaHCO3alkalinity acclimation. The control group was cultivated in freshwater. The acclimation group was subjected to 20 mmol/L alkalinity for 7 d and then placed in 40 mmol/L alkaline water. The non-acclimation group was placed in 40 mmol/L alkaline water. The experimental media were prepared with tap water and NaHCO3(domestic, analytically pure) after aeration for 2 d, and the alkalinity was measured and calibrated by acid-base titration. The fish were fed once a day and fasted for 24 h before sampling.
Blood physiological and biochemical indices (water osmotic pressure, white blood cells, lymphocytes, neutrophils, monocytes, hemoglobin, red blood cells, platelets, platelet hematocrit, urea, and albumin), as well as indices related to the liver antioxidant system of juvenile fish in 40 mmol/L alkalinity water, were measured at 0 h, 6 h, 12 h, 24 h, 48 h, 96 h, and 7 d. In the control group, the same sampling was conducted at the corresponding moments. The physiological and biochemical indices for both domesticated and non-domesticated fish initially increased and then decreased across the sampling period (<0.05). In both the domesticated and non-domesticated groups, the antioxidant indices of fish liver tissue, superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GSH-Px), and malondialdehyde (MDA), increased initially and then decreased across the sampling period (<0.05). The peak values of each parameter in the domesticated group were significantly lower than those of the non-domesticated group (<0.05). No significant changes were observed in the control group during this period (>0.05). The peak values of leukocytes, lymphocytes, neutrophils, and monocytes appeared at 48 h in the acclimated group and at 24 h in the non-acclimated group; the blood hemoglobin content, red blood cell content, platelet content, hematocrit, SOD, and CAT peaked in both the acclimated and non-acclimated groups at 24 h. After 7 d, the urea, lymphocyte, monocyte, platelet, red blood cell, hemoglobin, and albumin content of the blood and aspartate aminotransferase (AST), alanine aminotransferase (ALT), SOD, CAT, GSH-PX, and MDA content of the liver tissue were significantly lower in the acclimation group than in the non-acclimation group (<0.05), but the plasma albumin content and glutathione catalase in the acclimated group returned to the level of the control group (>0.05). Studies have shown that after alkalinity acclimation, juvenileare less stressed and the body tissue shows less damage and greater recovery under high alkalinity. At the physiological level, the body has higher adaptability. This study provides a theoretical basis for the establishment of saline-alkali aquaculture technology for the cultivation ofthat is based on the adaptability of the species to gradual changes in the alkalinity of the environment.
; Alkalinity; Physiological and biochemical; Antioxidant enzymes
XU Wei, E-mail: xwsc23@163.com; ZHAO Zhigang, E-mail: zhaozhigang@hrfri.ac.cn
10.19663/j.issn2095-9869.20210712001
S965.199
A
2095-9869(2022)04-0012-10
*国家重点研发计划资助项目(2020YFD0900402; 2019YFD0900404)、中国水产科学研究院基本科研业务费项目(2020TD56)和中央公益性科研院所基本科研业务费专项(HSY201803M; HSY201809M)共同资助 [This work was supported by National Key Research and Development Project (2020YFD0900402; 2019YFD0900404), Basic Project of Chinese Academy of Fishery Science (2020TD56), and Central Level Public Welfare Scientific Research Institute Basic Scientific Research Business Expense Special (HSY201803M; HSY201809M)]. 李海涛,E-mail: 1834376935@qq.com
徐 伟,研究员,E-mail: xwsc23@163.com;赵志刚,副研究员,E-mail: zhaozhigang@hrfri.ac.cn
2021-07-12,
2021-11-01
http://www.yykxjz.cn/
李海涛, 徐伟, 赵志刚, 罗亮, 李明帅, 刘金雨, 王雨, 张瑞, 郭坤, 耿龙武, 杜汝君, 丛艳峰.碱度驯化对大鳞鲃幼鱼血液生理生化及肝脏抗氧化系统的影响.渔业科学进展, 2022, 43(4): 12–21
LI H T, XU W, ZHAO Z G, LUO L, LI M S, LIU J Y, WANG Y, ZHANG R, GUO K, GENG L W, DU R J, CONG Y F. Effects of alkalinity acclimation on blood physiology and biochemistry and liver antioxidant system of juvenileProgress in Fishery Sciences, 2022, 43(4): 12–21
(编辑 陈 辉)
猜你喜欢
求学·理科版(2022年4期)2022-04-03
福建基础教育研究(2020年2期)2020-05-28
饮食与健康·下旬刊(2019年6期)2019-03-09
恋爱婚姻家庭·养生版(2018年11期)2018-11-26
恋爱婚姻家庭(2018年33期)2018-07-22
养生保健指南(2018年4期)2018-04-11
建筑科技与经济(2017年7期)2017-07-20
医学信息(2017年9期)2017-04-22
养生保健指南(2016年12期)2017-01-06
现代农业科技(2009年19期)2009-03-20
