
不同浓度亚硝酸盐亚急性胁迫对凡纳滨对虾生长与免疫功能的影响*
2022-09-05 02:13黎兰诗梁震宇成良峰戴习林
渔业科学进展 2022年4期
方 成 黎兰诗 梁震宇 成良峰 戴习林
不同浓度亚硝酸盐亚急性胁迫对凡纳滨对虾生长与免疫功能的影响*
方 成 黎兰诗 梁震宇 成良峰 戴习林①
(上海海洋大学 水产科学国家级试验教学示范中心 农业农村部淡水水产种质资源重点实验室 水产动物遗传育种中心上海市协同创新中心 上海 201306)
为阐明盐度为5条件下不同浓度亚硝酸盐亚急性胁迫对凡纳滨对虾()生长与免疫功能的影响,本研究设置5个亚硝酸盐浓度组(0.50、0.90、1.70、3.20和6.00 mg/L)和对照组(0.05 mg/L),检测分析了亚硝酸盐胁迫40 d后凡纳滨对虾免疫相关酶活性、丙二醛(MDA)含量以及免疫和生长相关基因表达的变化。结果显示,凡纳滨对虾死亡率随亚硝酸盐浓度的增加而升高,6.00 mg/L浓度组体质量增长率(WGR)和体长增长率(LGR)均显著低于对照组(<0.05)。部分浓度组亚硝酸盐对凡纳滨对虾肝胰腺和血清中的免疫相关酶活性具有一定的诱导作用。其中,当亚硝酸盐浓度高于0.50 mg/L时,肝胰腺超氧化物歧化酶(SOD)活性显著高于对照组(<0.05);0.50、0.90和1.70 mg/L浓度组的过氧化氢酶(CAT)活性显著高于对照组(<0.05);血清中CAT和SOD活性随亚硝酸盐浓度的增加均呈先降低后升高再降低的趋势;0.90 mg/L浓度组的肝胰腺和血清中酸性磷酸酶(ACP)和碱性磷酸酶(AKP)活性均显著高于对照组(<0.05)。MDA含量变化无明显规律。此外,血清中谷丙转氨酶(GPT)活性显著升高(<0.05)。实时荧光定量PCR结果显示,除0.50 mg/L浓度组外,其他浓度组的和基因表达量显著升高(<0.05);各浓度组的和基因表达量显著低于对照组(<0.05)。经亚硝酸盐胁迫40 d后,各浓度组凡纳滨对虾的生长和免疫功能均受到明显的阻遏作用。在盐度为5条件下,为确保凡纳滨对虾的健康养殖,亚硝酸盐浓度应控制在0.50 mg/L以内。
凡纳滨对虾;亚硝酸盐;酶活性;环境胁迫
凡纳滨对虾()是我国非常重要的水产经济动物,自引入我国以来,养殖规模不断扩大,创造了巨大的经济价值。但养殖过程中出现的亚硝酸盐成为了制约凡纳滨对虾产业发展的重要因素。亚硝酸盐是养殖水体中常见的化学物质。氨氮在亚硝化作用下变为亚硝酸盐,之后通过硝化作用转变为硝酸盐。但硝化作用与亚硝化作用一般无法达到平衡,因此,亚硝酸盐容易在水体中积蓄。亚硝酸盐可导致虾类生长和发育异常,病菌易感性增加(Tseng, 2004;臧维玲等, 1996; Mallasen,2006),并会造成一定程度的组织损伤。一定浓度的亚硝酸盐可使血淋巴与氧结合能力降低(Chen,1995),最终导致虾窒息死亡。彭自然等(2004)研究发现,凡纳滨对虾在含有亚硝酸盐的水体中养殖14 d后,其生长和发育受到了明显抑制。胡义波等(2005)研究发现,凡纳滨对虾经亚硝酸盐胁迫后体内血细胞减少,细胞超微结构受到明显影响。吴中华等(1999)研究发现,亚硝酸盐慢性胁迫条件下,中国对虾()各个组织均受到不同程度的影响,其中肝胰腺组织病变较为严重。Cheng等(2000)研究发现,斑节对虾()经亚硝酸盐胁迫后,肝胰腺和中肠中亚硝酸盐积累较多。Zhang等(2015)研究发现,在经氨氮和亚硝酸盐联合胁迫24 h后,罗氏沼虾()血淋巴中活性氧升高,抗氧化酶活性受到影响。葛红星等(2014)研究发现,凡纳滨对虾在不同浓度亚硝酸盐水体中暴露20 d后,对副溶血弧菌()的易感性提高。总体而言,一定浓度的亚硝酸盐可对凡纳滨对虾的免疫功能和生长造成较明显的阻遏作用。近年来,关于亚硝酸盐的研究主要集中于亚硝酸盐对虾类的急性毒性影响,主要研究短期亚硝酸盐胁迫后虾体内抗氧化酶活性、氧化损伤以及对病菌易感性的变化,而较少有针对亚硝酸盐对凡纳滨对虾亚急性毒性的研究。本实验研究凡纳滨对虾在5个不同亚硝酸盐浓度的水体中养殖40 d后体内部分免疫相关酶活性、免疫和生长相关基因表达变化情况,以期为亚硝酸盐亚急性胁迫对凡纳滨对虾的影响提供参考依据。
1 材料与方法
1.1 实验材料
实验用虾取自上海申漕特种水产有限公司,挑选健康、体长为(7.45±0.21) cm、体质量为(4.75±0.51) g的凡纳滨对虾暂养于水泥池中。
1.2 亚硝酸盐亚急性胁迫实验
凡纳滨对虾暂养1周后开始实验。通过预实验得知,凡纳滨对虾96 h半致死浓度(96h-LC50)约为30 mg/L,取该浓度的20%为最高浓度并按等对数间距设置5个亚硝酸盐浓度,分别为0.50、0.90、1.70、3.20和6.00 mg/L。每个处理设3组平行,每个平行放30尾虾。对照组亚硝酸盐浓度约为0.05 mg/L。实验在卤虫池中进行,水体体积为100 L,每日换水1/2。每日利用分光光度计测得水体中亚硝酸盐浓度,并使用10 g/L NaNO2溶液控制水体中的亚硝酸盐浓度。
实验期间,温度控制在26℃~27℃,盐度为5,总氨氮≤0.5 mg/L,非离子氨≤0.02 mg/L,pH控制在7.8±0.2,24 h曝气。每日投喂3次人工饲料(鱼粉23%、酵母粉4%、大豆磷脂4%、豆粕20%、花生粕7.4%等),每日投喂量为虾体质量的11%~16%。每日记录虾的死亡状况。养殖实验进行40 d后(取样前停饲24 h),每个平行取3尾虾的肝胰腺放入离心管中,存放于–80℃冰箱,用于酶活测定和RNA提取。另外,使用1 mL注射器在第2腹肢基部抽取凡纳滨对虾的血淋巴,并放入离心管中。血淋巴存于4℃冰箱,放置24 h后离心取上清液,放置于–80℃冰箱,用于酶活测定。
1.3 生长指标的测定
通过以下公式计算得到不同浓度感染组的死亡率(MR)、体质量增长率(WGR)和体长增长率(LGR)。
死亡率(MR, %)=T/0×100% (1)
体质量增长率(WGR, %)=(T–0)/0×100% (2)
体长增长率(LGR, %)=(T–0)/0×100% (3)
式中,0为初始数量;T为实验结束时死亡数量;0和T分别为初始平均体质量和实验结束时平均体质量;0和T分别为初始平均体长和实验结束时平均体长。
1.4 抗氧化酶、代谢酶活性和MDA含量的测定
采用超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、碱性磷酸酶(AKP)、酸性磷酸酶(ACP)、丙二醛(MDA)、谷丙转氨酶(GPT)和谷草转氨酶(GOT)试剂盒(南京建成生物工程有限公司)测定凡纳滨对虾肝胰腺和血清的生理生化指标。
1.5 总RNA的提取
使用RNA Easy Fast动物组织/细胞总RNA提取试剂盒(DP451)(北京天根生化科技有限公司)提取肝胰腺组织RNA。采用核酸蛋白测定仪测定的RNA样品的OD260 nm和OD280 nm值,确定其浓度,并计算二者比值以确定RNA的纯度。0.8%琼脂糖凝胶电泳检测RNA质量。
1.6 反转录及引物设计
按照PrimeScript™1st Strand cDNA Synthesis Kit说明书(TaKaRa)将之前所提取的总RNA进行反转录,得到的cDNA存放于–20℃冰箱中。根据NCBI数据库中凡纳滨对虾(DQ005531)、(AY518322)、(AY645906)等基因的保守区域和基因的序列设计特异性引物(表1)。
表1 实验中所用引物序列

Tab.1 Sequences of the primers in this study
1.7 荧光定量PCR
使用ABI PRISM®7900 Sequence Detection System仪器,根据SYBR®Green Real-time PCR Master Mix试剂盒说明书,对各组实验样品的目的基因和基因cDNA进行定量测定。ABI PRISM®7900 Sequence Detection System的具体反应程序设置为:94℃预变性30 s,94℃变性5 s,60℃退火25 s,72℃延伸30 s,共40个循环。最后根据熔解曲线分析PCR产物质量。采用2–∆∆Ct计算法对各组样品测得的基因数据进行定量分析。
1.8 数据统计与分析
所有数据使用平均值±标准差(Mean±SD)表示。采用SPSS 17.0和Excel 2010分析数据,使用Duncan´s multiple range test检验统计差异,<0.05为差异显著。
2 结果
2.1 不同浓度亚硝酸盐对凡纳滨对虾死亡率和生长的影响
由表2可知,凡纳滨对虾死亡率随亚硝酸盐浓度的增加而升高。各浓度组死亡率均显著高于对照组(<0.05),6.00 mg/L浓度组死亡率为最高(<0.05)。
凡纳滨对虾体质量增长率和体长增长率总体上随亚硝酸盐浓度的增加呈降低的趋势。6.00 mg/L浓度组体质量增长率和体长增长率为最低,并显著低于对照组(<0.05),其他浓度组体长增长率和体质量增长率组间无显著差异(>0.05)。
表2 不同浓度亚硝酸盐对凡纳滨对虾死亡率和生长的影响
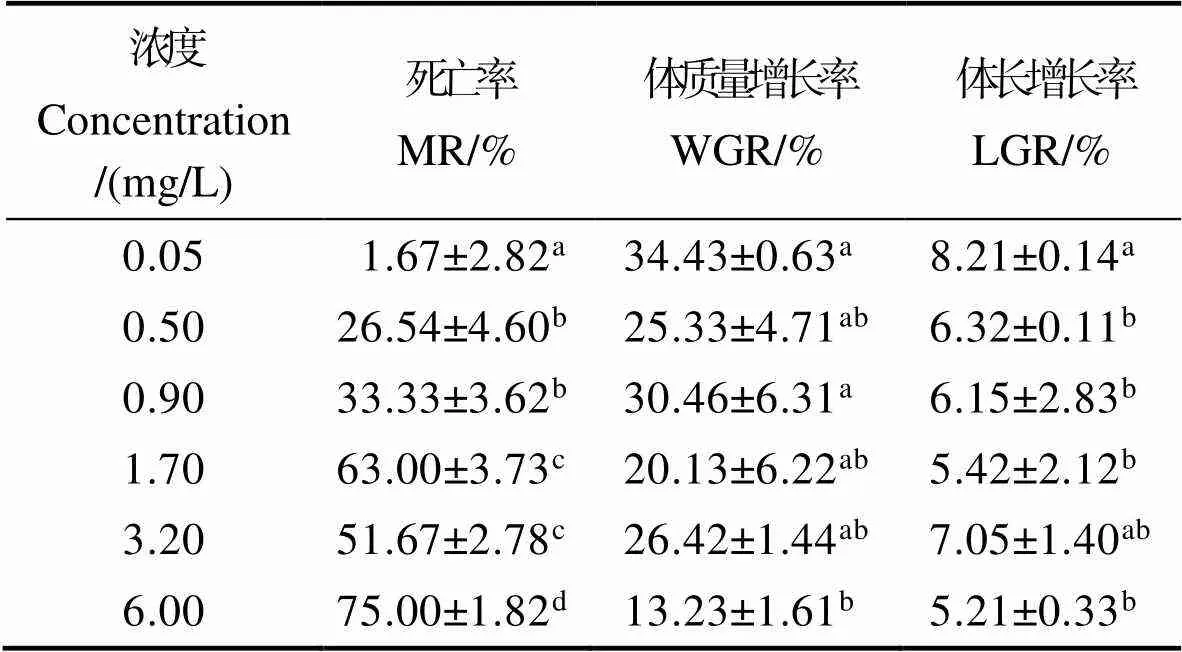
Tab.2 Effect of different concentrations of nitrite on growth performance and mortality rate of L. vannamei
2.2 不同浓度亚硝酸盐对凡纳滨对虾抗氧化酶活性和MDA含量的影响
除0.50 mg/L浓度组外,其他浓度组血清中SOD和CAT活性均高于对照组,其中,SOD和CAT活性分别在3.20和0.90 mg/L浓度组达到最高并显著高于对照组(<0.05)(图1a、图1b)。除0.50 mg/L浓度组外,其他浓度组肝胰腺SOD活性均高于对照组,0.90 mg/L浓度组SOD活性为最高(<0.05)(图1c)。如图1d所示,0.50、0.90和1.70 mg/L浓度组肝胰腺CAT活性显著高于对照组(<0.05)。3.20和6.00 mg/L浓度组CAT活性显著低于对照组,且各组间存在显著差异(<0.05)(图1d)。
血清中AKP和ACP活性随亚硝酸盐浓度的增加总体呈升高的趋势,而在肝胰腺中则呈波动变化。3.20 mg/L浓度组血清AKP和ACP活性达到最高(<0.05)(图1e、图1f)。而肝胰腺中ACP和AKP活性在0.90 mg/L浓度组为最高(<0.05)(图1g、图1h)。
MDA含量总体呈波动变化,其中,3.20 mg/L浓度组血清MDA含量显著高于对照组(<0.05),其他浓度组与对照组无显著差异(>0.05)(图1i)。除0.50 mg/L浓度组,其他浓度组肝胰腺MDA含量均高于对照组,其中,0.90、1.70和6.00 mg/L浓度组显著高于对照组(<0.05) (图1j)。

图1 不同浓度亚硝酸盐对凡纳滨对虾抗氧化酶活性和MDA含量的影响
不同字母代表存在显著差异(<0.05),下同。
Different letters indicate significant difference (<0.05). The same as below.
2.3 不同浓度亚硝酸盐对凡纳滨对虾代谢酶活性的影响
除0.50 mg/L浓度组外,其他浓度组GPT活性均显著高于对照组(<0.05)。各浓度组GPT活性无显著差异(0.05)(图2a)。3.20和6.00 mg/L浓度组GOT活性显著高于其他浓度组(0.05),其他浓度组间无显著差异(0.05)(图2b)。
2.4 不同浓度亚硝酸盐对凡纳滨对虾肝胰腺中免疫相关基因表达的影响
除0.50 mg/L浓度组外,其他浓度组和基因表达量显著高于对照组(<0.05)。和基因表达量分别在6.00和3.20 mg/L浓度组达到最高(<0.05)(图3a、图3b)。、和基因均随亚硝酸盐浓度的增加呈降低的趋势,各浓度组基因表达量均显著低于对照组(<0.05)(图3c~图3e)。
2.5 不同浓度亚硝酸盐对凡纳滨对虾肝胰腺中生长相关基因表达的影响
和基因表达量随亚硝酸盐浓度的增加总体呈先降低后升高的趋势。各浓度组的基因表达量均显著低于对照组(<0.05),且各浓度组无显著差异(>0.05)。6.00 mg/L浓度组的基因表达量升高,并显著高于其他浓度组(<0.05)(图4a、图4b)。
基因表达量随亚硝酸盐浓度的增加呈波动变化,0.50 mg/L浓度组达到最低并显著低于对照组(<0.05)。0.90和6.00 mg/L浓度组显著高于对照组(<0.05)。1.70和3.20 mg/L浓度组与对照组无显著差异(>0.05)(图4c)。

图2 不同浓度亚硝酸盐对凡纳滨对虾GOT和GPT酶活性的影响
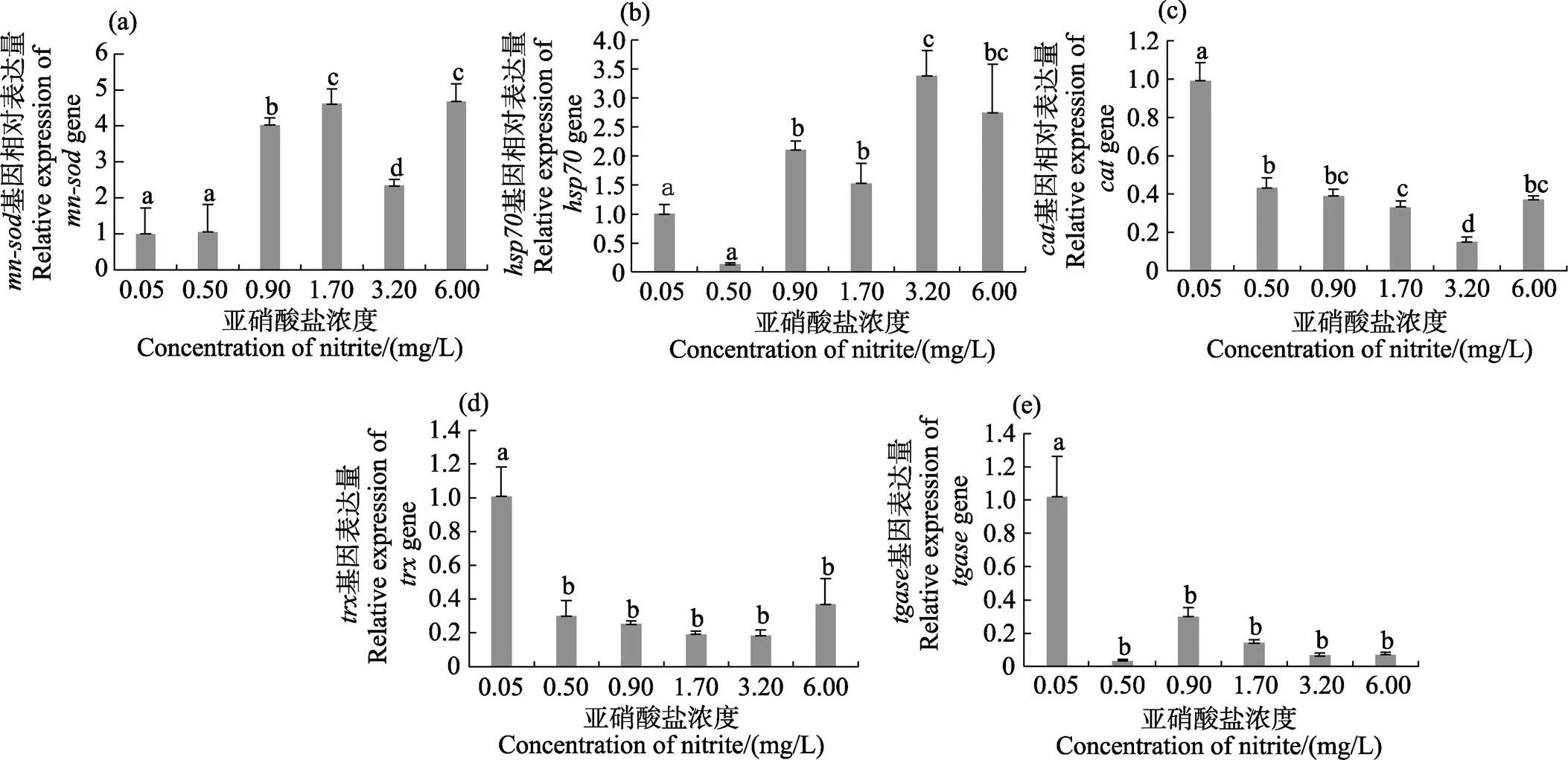
图3 不同浓度亚硝酸盐对凡纳滨对虾肝胰腺中免疫相关基因表达的影响

图4 不同浓度亚硝酸盐对凡纳滨对虾肝胰腺中生长相关基因表达的影响
3 讨论
甲壳动物在受到pH、重金属等环境因子胁迫时,会发生生理功能紊乱和免疫功能下降(汪蕾等, 2016; Lyu,2014; 周萌等, 2015),异物入侵会造成虾的氧化损伤(Kultz, 2005)并最终导致细胞功能出现障碍,而抗氧化酶可帮助机体消除多余的自由基,从而使机体免受氧化损伤。SOD和CAT是凡纳滨对虾体内2种重要的抗氧化酶,可协同清除自由基(洪美玲等, 2007)。Zhang等(2015)对罗氏沼虾进行了氨氮和亚硝酸盐的联合急性胁迫实验,当氨氮浓度为0 mg/L时,随亚硝酸盐浓度的升高,罗氏沼虾血清中SOD活性先降低后升高。葛红星等(2014)研究了不同浓度亚硝酸盐(2、4、6和10 mg/L)急性胁迫对凡纳滨对虾免疫相关酶活性的影响,其中6和10 mg/L浓度组对各时间段的抗氧化酶活性有明显的抑制作用,而其他浓度组不明显。本研究中,除0.50 mg/L浓度组外,其他浓度组SOD活性均显著升高,其中0.90和1.70 mg/L浓度组CAT活性显著升高,较高浓度组(3.20和6.00 mg/L) CAT活性有一定程度的下降。这与上述研究结果不一致,原因可能是当亚硝酸盐浓度较低时出现“毒性兴奋效应”,而较高浓度的亚硝酸盐则超出抗氧化酶的正常调节范围。MDA是脂质氧化过程中的产物(Liu, 2011),其含量可反映机体所受的氧化损伤程度。本研究肝胰腺中各浓度组MDA含量升高时,SOD活性也相应升高,说明机体中的抗氧化酶活性会随MDA含量的变化发生改变,以便及时帮助机体清除自由基。ACP和AKP是2种水解酶,在去磷酸化过程中起重要作用,且可参与机体解毒过程(何海琪等, 1992)。沈敏等(2019)研究发现,高盐胁迫可使凡纳滨对虾体内的ACP活性显著升高,而当盐度高于45后,ACP活性降低。彭军辉等(2018)研究发现,ACP和AKP活性随氨氮胁迫时间的增加先升高后降低。本研究中,AKP与ACP活性总体呈升高的趋势,未受到明显抑制,原因可能是AKP和ACP仍在进行解毒作用且亚硝酸盐浓度没有超出ACP和AKP的正常调节范围。此外,GOT和GPT活性可反映机体肝组织健康状态和功能,肝胰腺功能与免疫、代谢等能力息息相关,当对虾处于健康状态时,血清中只有少量的转氨酶;当虾肝组织受损时,肝细胞会向血淋巴中释放大量转氨酶(许宏庆等, 2006)。本研究中,除0.50 mg/L浓度组外,其他浓度组中的转氨酶活性均显著高于对照组,说明肝胰腺功能可能受到明显影响。
肝胰腺是凡纳滨对虾重要的消化器官,具有代谢、消化和免疫等重要功能(蒋昊, 2009),因此,肝胰腺是对虾受到环境胁迫后的重要研究对象。Wang等(2009)研究发现,pH胁迫对凡纳滨对虾肝胰腺基因的表达有诱导作用。本研究中,除0.50 mg/L浓度组外,其他浓度组肝胰腺基因表达量均显著升高,与Wang等(2009)的结果相似,表明在亚硝酸盐胁迫40 d后,肝胰腺中的SOD可能仍在发挥作用,且随亚硝酸盐浓度的增加,基因表达量升高以增强抗氧化能力。Zheng等(2016)研究发现,日本对虾()肝胰腺中基因表达量在高浓度亚硝酸盐(20 mg/L)的影响下,短时间会明显升高,但随时间的增加,基因表达量逐渐降低。本研究中,基因表达量随亚硝酸盐浓度的增加显著降低,原因可能是长时间的亚硝酸盐胁迫会对基因表达产生抑制作用,从而减少CAT的产生。值得注意的是,CAT酶量减少,但部分浓度组活性仍较高,造成该结果的原因有待研究。本研究中,基因表达量明显升高,而基因则相反,表明凡纳滨对虾在SOD活性方面对亚硝酸盐胁迫有较强适应性。
硫氧还蛋白(TRX)是具有氧化还原活性的蛋白质,属于过氧化酶(PRX)家族。PRX广泛存在于各种哺乳动物、植物和无脊椎动物中,具有多种生理功能,具有抗氧化、抗高温等功能(Radyuk, 2003)。在无脊椎动物中,PRX通过消除机体内H2O2来参与体内抗氧化反应。郭慧等(2017)研究发现,亚硝酸盐胁迫能显著抑制凡纳滨对虾肝胰腺中基因的表达。本研究各浓度组的基因表达量均显著降低。说明在长时间的亚硝酸盐胁迫下,肝胰腺中的TRX在抗亚硝酸盐胁迫过程中被不断消耗,且基因表达受到抑制。
谷氨酰胺转移酶(TGASE)在对虾免疫功能中具有重要作用。Fagutao等(2012)研究发现,将日本对虾基因沉默后,虾体内的血细胞数量显著降低,同时抗菌肽和溶菌酶的基因表达显著降低,细菌总数升高;Yao等(2019)研究发现,凡纳滨对虾血蓝蛋白可与TGASE直接相互作用并调节其表达。以上研究表明,TGASE参与对虾凝血过程。虾类受到异物入侵时,血细胞会释放TGASE并形成血浆蛋白(Maningas,2008)。卢芷程(2018)则发现,溶藻弧菌()可在短期内诱导基因的表达,而随时间延长,基因表达量持续降低。说明长时间的环境胁迫会导致基因表达受到抑制。这与本研究结果基本一致,推测凡纳滨对虾的凝血功能可能受到一定程度的影响。
热休克蛋白(HSPs)作为一种分子伴侣,可对各种外界刺激造成的细胞损伤进行自我修复,并在虾类的应激反应和先天免疫系统中发挥重要作用(雷爱莹等, 2008)。当虾类机体内自由基达到一定数量并超出机体消除自由基的能力后,热休克蛋白基因会被诱导表达。董学兴(2019)研究发现,短期氨氮胁迫可诱导罗氏沼虾基因大量表达。本研究中,除0.50 mg/L浓度组外,其他浓度组基因表达量均显著高于对照组,表明凡纳滨对虾体内自由基与抗氧化酶系统可能失衡,热休克蛋白基因被诱导表达并参与机体修复。综上所述,凡纳滨对虾总体上生理和免疫功能均受到抑制,但在SOD活性方面表现出较强的适应性。
此外,本研究发现,亚硝酸盐会显著抑制凡纳滨对虾的生长。随着亚硝酸盐浓度的增加,体质量增长率和体长增长率均呈降低的趋势,与肖威等(2020)的结果一致。本研究检测的4种生长相关基因均与凡纳滨对虾的生长发育有非常密切的关系,其中,组织蛋白酶B (CTSB)在脊椎动物中研究较多,它参与多种生理功能,如食物消化、免疫和蜕皮等过程(Wang, 2008; Stephen, 2012)。几丁质酶又被称为壳多糖,是甲壳动物在蜕皮过程中最重要的酶之一。吕黎等(2011)报道,几丁质酶在免疫方面起到一定作用。胰蛋白酶则具有消化食物和水解蛋白质的功能。本研究中,各亚硝酸盐浓度组胰蛋白酶基因和几丁质酶基因均显著降低,表明凡纳滨对虾的消化功能受到了抑制。虾类在遭受外界病原体或异物侵袭时,体内的能量代谢会发生变化。由于外界环境的恶化,体内稳态失衡,细胞产生应激反应,包括DNA和蛋白质损伤修复、细胞周期停滞或凋亡,并从细胞的生长状态转型至细胞的修复状态(Kultz, 2005)。这些过程会消耗大量能量,从而影响虾的生长。本研究中,各浓度组胰蛋白酶基因和几丁质酶基因表达量显著降低,可能与凡纳滨对虾体内大量能量参与免疫调控及维持渗透压平衡有关。李忠帅等(2021)研究发现,亚硝酸盐可在凡纳滨对虾体内迅速积累并加快其能量代谢,进一步说明亚硝酸盐抑制凡纳滨对虾生长的机制。郭慧等(2017)研究发现,亚硝酸盐可显著诱导基因的表达,之前郭慧等(2013)研究发现,CTSB可能参与细胞凋亡。本研究中,基因表达无明显规律,可能因为该基因功能较复杂,未来需要作进一步的研究。
4 结论
在亚硝酸盐浓度分别为0.50、0.90、1.70、3.20和6.00 mg/L,盐度为5的水体中养殖40 d后,凡纳滨对虾各浓度组死亡率显著升高,生长性能和免疫功能受到阻遏,但在SOD活性方面表现出较强的适应性。各浓度组生长相关酶基因表达量显著降低,凡纳滨对虾消化功能受到抑制。此外,当亚硝酸盐浓度大于0.9 mg/L时,凡纳滨对虾肝胰腺功能可能受到明显影响。在盐度为5条件下,为保证凡纳滨对虾的健康养殖,亚硝酸盐浓度应控制在0.50 mg/L以下。
CHEN J C, CHENG S Y. Hemolymph oxygen content, oxyhemocyanin, protein levels and ammonia excretion in the shrimpexposed to ambient nitrite. Journal of Comparative Physiology B Biochemical Systems and Environmental Physiology, 1995, 164(7): 530–535
CHENG S Y, CHEN J C. Accumulation of nitrite in the tissues ofexposed to elevated ambient nitrite after different time periods. Archives of Environmental Contamination and Toxicology, 2000, 39(2): 183–192
DONG X X. Stress response ofto ammonia-N and its environmental effects in different culture patterns. Doctoral Dissertation of Shanghai Ocean University, 2019 [董学兴. 罗氏沼虾对氨氮的胁迫响应及其不同养殖模式的环境效应. 上海海洋大学博士研究生学位论文, 2019]
FAGUTAO F F, MANINGAS M B, KONDO H,. Transglutaminase regulates immune-related genes in shrimp. Fish and Shellfish Immunology, 2012, 32(5): 711–715
GE H X, LI J, CHEN P,. Susceptibility ofto: The influence of environmental nitrite nitrogen. Journal of Fishery Sciences of China, 2014, 21(3): 629–636 [葛红星, 李健, 陈萍, 等. 亚硝酸盐氮胁迫下凡纳滨对虾对副溶血弧菌的易感性. 中国水产科学, 2014, 21(3): 629–636]
GUO H, TAN C T, YOU L Y,. Effects of nitrite stress on gene expression of antioxidant enzymes, heat shock protein and cathepsin B in hepatopancreas of. Journal of Guangdong Ocean University, 2017, 37(3): 117–122 [郭慧, 谭翠婷, 游林玉, 等. 亚硝酸盐胁迫对凡纳滨对虾肝胰腺抗氧化酶、热休克蛋白和组织蛋白酶B基因表达量的影响. 广东海洋大学学报, 2017, 37(3): 117–122]
GUO H, XIAN J A, LI B,. Gene expression of apoptosis-related genes, stress protein and antioxidant enzymes in hemocytes of white shrimpunder nitrite stress. Comparative Biochemistry and Physiology Part C: Toxicology and Pharmacology, 2013, 157: 366–371
HE H Q, SUN F. Studies on the characteristics of acid and alkaline phosphatases in Chinese shrimp. Oceanologia et Limnologia Sinica, 1992, 23(5): 555–560 [何海琪, 孙凤. 中国对虾酸性和碱性磷酸酶的特性研究. 海洋与湖沼, 1992, 23(5): 555–560]
HONG M L, CHEN L Q, GU S Z,. Effects of ammonia exposure on immunity indicators of haemolymph and histological structure of hepatopancreas in Chinese mitten crab (). Journal of Fishery Sciences of China, 2007, 14(3): 412–418 [洪美玲, 陈立侨, 顾顺樟, 等. 氨氮胁迫对中华绒螯蟹免疫指标及肝胰腺组织结构的影响. 中国水产科学, 2007, 14(3): 412–418]
HU Y B, WANG Y, JIANG N C. Effects of ammonia-N and nitrite-N on the hemocyte count and ultrastructure of.Journal of Zhejiang University (Science Edition), 2005, 32(6): 691–697 [胡义波, 王玥, 姜乃澄. 氨态氮、亚硝态氮对罗氏沼虾血细胞及超微结构的影响. 浙江大学学报(理学版), 2005, 32(6): 691–697]
JIANG H. Comparative proteomic profiles of the hepatopancreas inresponse to stresses. Doctoral Dissertation of Chinese Academy of Sciences (Institute of Oceanology), 2009 [蒋昊. 中国明对虾在胁迫条件下肝胰脏的差异蛋白质组学研究. 中国科学院研究生院(海洋研究所)博士研究生学位论文, 2009]
KULTZ D. Molecular and evolutionary basis of the cellular stress response. Annual Review of Physiology, 2005, 67: 225–257
LEI A Y, ZENG D G. Effects of compound Chinese herbal on the expression of heat stress protein 70 gene in white shrimp (). Journal of Southern Agriculture, 2008, 39(6): 830–833 [雷爱莹, 曾地刚. 复方中草药对凡纳滨对虾热应激蛋白70基因表达的影响. 广西农业科学, 2008, 39(6): 830–833]
LI Z S, MA S, SHAN H W,. Accumulation of nitrite and responses of energy metabolism exposed to nitrite stress in. Journal of Fisheries of China, 2021, 45(11): 1825–1834 [李忠帅, 马甡, 单洪伟, 等. 亚硝态氮胁迫下凡纳滨对虾体内亚硝态氮的积累与能量代谢响应. 水产学报, 2021, 45(11): 1825–1834]
LIU L X, XI Q Y, YANG L,. The effect of dietarypolysaccharide extract on the immune responses in white shrimp,. Fish and Shellfish Immunology, 2011, 30(2): 495–500
LU Z C, XU L J, LU W Y,. Effects ofon hemocytes toxicity, apoptosis and immune-related genes in. Journal of Southern Agriculture, 2018, 49(12): 2559–2565 [卢芷程, 许銮佳, 卢文宇, 等. 溶藻弧菌对凡纳滨对虾血细胞毒性及细胞凋亡和免疫相关基因的影响. 南方农业学报, 2018, 49(12): 2559–2565]
LÜ L, NING Q J. Research progress on the structure and function of chitinase gene in crustacean. Progress in Physiological Sciences, 2011, 42(6): 457–459 [吕黎, 宁黔冀. 甲壳动物几丁质酶基因结构与功能的研究进展. 生理科学进展, 2011, 42(6): 457–459]
LYU K, ZHU X X, Chen R,. Molecular cloning of manganese superoxide dismutase gene in the cladoceran: Effects of microcystin, nitrite, and cadmium on gene expression profiles. Aquatic Toxicology, 2014, 148(1): 55–64
MALLASEN M, WAGNER C V. Effect of nitrite on larval development of giant river prawn. Aquaculture, 2006, 261(4): 1292–1298
MANINGAS M, KONDO H, HIRONO I,. Essential function of transglutaminase and clotting protein in shrimp immunity. Molecular Immunology, 2008, 45(5): 1269–1275
PENG J H, CHEN L Y, CHENG C H,Acute toxicity of ammonia nitrogen toand its influence on immune factors in serum. Progress in Fishery Sciences, 2018, 39(5): 114–121 [彭军辉, 陈丽英, 程长洪, 等. 氨氮对拟穴青蟹的急性毒性及对其血清免疫相关酶活力的影响. 渔业科学进展, 2018, 39(5): 114–121]
PENG Z R, ZANG W L, GAO Y,. Toxic effects of ammonia and nitrite onjuvenile. Journal of Shanghai Fisheries University, 2004, 13(3): 274–278 [彭自然, 臧维玲, 高杨, 等. 氨和亚硝酸盐对凡纳滨对虾幼虾的毒性影响. 上海水产大学学报, 2004, 13(3): 274–278]
RADYUK S N, SOHAL R S, ORR W C. Thioredoxin peroxidases can foster cytoprotection or cell death in response to different stressors: Over- and under- expression of thioredoxin peroxidase incells. Biochemical Journal, 2003, 371(3): 743–752
SHEN M, ZHAO Y C, LING T,Effects of high-salt abrupt on growth and related enzyme activities in. Oceanologia et Limnologia Sinica, 2019, 50(1): 204–209 [沈敏, 赵玉超, 凌涛, 等. 高盐突变对凡纳滨对虾()生长性能及相关酶活力的影响. 海洋与湖沼, 2019, 50(1): 204–209]
STEPHEN A, ROJO L, ARAUJO-BERNAL S,. Cathepsin B from the white shrimp: cDNA sequence analysis, tissues-specific expression and biological activity. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2012, 161(1): 32–40
TSENG I T, CHEN J C. The immune response of white shrimpand its susceptibility tounder nitrite stress. Fish and Shellfish Immunology, 2004, 17(4): 325–333
WANG L, ZHANG X X, ZHENG P H,.Effects of sulfide stress on expressions of antioxidant enzymes inhaemocytes. Sichuan Journal of Zoology, 2016, 35(6): 884–888 [汪蕾, 张秀霞, 郑佩华, 等. 硫化物胁迫对凡纳滨对虾血细胞抗氧化酶基因表达的影响. 四川动物, 2016, 35(6): 884–888]
WANG W N, ZHOU J, WANG P,. Oxidative stress, DNA damage and antioxidant enzyme gene expression in the pacific white shrimp,when exposed to acute pH stress. Comparative Biochemistry and Physiology Part C: Toxicology and Pharmacology, 2009, 150(4): 428–435
WANG X M, LIU B Z, WANG G D,. Molecular cloning and functional analysis of cathepsin B in nutrient metabolism during larval development in. Aquaculture, 2008, 282(1/2/3/4): 41–46
WU Z H, LIU C B, LIU C R,.Histopathological research on chronic poisoning ofby nitrite and ammonia. Journal of Central China Normal University (Natural Sciences), 1999, 33(1): 119–122 [吴中华, 刘昌彬,刘存仁, 等. 中国对虾慢性亚硝酸盐和氨中毒的组织病理学研究. 华中师范大学学报(自然科学版), 1999, 33(1): 119–122]
XIAO W, SHAN H W, MA S,. Effects of chronic nitrite stress on body composition and glucose metabolism of.Progress in Fishery Sciences, 2020, 41(6): 74–81 [肖威, 单洪伟, 马甡, 等. 亚硝态氮慢性胁迫对凡纳滨对虾体成分和糖代谢的影响. 渔业科学进展, 2020, 41(6): 74–81]
XV H Q, CUI Y H, LI J,. Effects of cadmium on some biochemical indexes in the serum of. Reservoir Fisheries, 2006, 26(5): 6–7, 20 [许宏庆, 崔勇华, 李晶, 等. 镉对克氏原螯虾血清中部分生化指标的影响. 水利渔业, 2006, 26(5): 6–7, 20]
YAO D, WANG Z, WEI M,. Analysis ofhemocyanin interacting proteins reveals its role in hemolymph clotting. Journal of Proteomics, 2019, 201: 57– 64
ZANG W L, JIANG M, ZHANG J D,. The toxic effects of NO2–-N and NH3-N onlarva. Journal of Shanghai Fisheries University, 1996, 5(1): 15–22 [臧维玲, 江敏, 张建达, 等. 亚硝酸盐和氨对罗氏沼虾幼体的毒性. 上海水产大学学报, 1996, 5(1): 15–22]
ZHANG Y F, YE C X, WANG A L,. Isolated and combined exposure to ammonia and nitrite in giant freshwater pawn (): Effects on the oxidative stress, antioxidant enzymatic activities and apoptosis in haemocytes. Ecotoxicology, 2015, 24: 1601–1610
ZHENG J B, MAO Y, SU Y Q,. Effects of nitrite stress on mRNA expression of antioxidant enzymes, immune-related genes and apoptosis-related proteins in. Fish and Shellfish Immunology, 2016, 58: 239– 252
ZHOU M, WU Z H, LIANG R S,. Biochemical and cellular immunological responses of Pacific white shrimp,to cold shock. Guangdong Agricultural Sciences, 2015, 24: 134–139 [周萌, 吴灶和, 梁日深, 等. 急性降温对凡纳滨对虾血液生化指标及细胞免疫指标的影响. 广东农业科学, 2015, 24: 134–139]
Changes in Immune System and Growth Performance ofafter 40-Day Challenge with Different Concentrations of Nitrite
FANG Cheng, LI Lanshi, LIANG Zhenyu, CHENG Liangfeng, DAI Xilin①
(Shanghai Ocean University, National Experimental Teaching Demonstration Center of Fisheries Science, Ministry of Agriculture and Rural Affairs, Key Laboratory of Freshwater Aquatic Germplasm Resources, Aquatic Animal Genetic Breeding Center Shanghai Collaborative Innovation Center, Shanghai 201306, China)
After 40 days of exposure to different concentrations of nitrate at a salinity of 5, the effect of nitrite on the immune system and growth performance ofwasexplored to provide a theoretical basis for constraining the effect of nitrite subacute stress on the speciesFive nitrite concentration groups (0.50, 0.90, 1.70, 3.20, and 6.00 mg/L) and a control group (0.05 mg/L) were used in this experiment. Furthermore, we detected the activity of immune-related enzymes, malondialdehyde (MDA) content, and relative expression of immune- and growth-related genes. Results showed that the mortality rate ofincreased with an increasing nitrite concentration, and the weight gain rate (WGR) and length gain rate (LGR) in the 6.00 mg/L group were significantly lower than those in the control group (<0.05). The activity of superoxide dismutase (SOD), catalase (CAT), acid phosphatase (ACP), and alkaline phosphatase (AKP) in the serum and hepatopancreas increased under the influence of nitrite in 0.90 and 3.20 mg/L groups. The activity of SOD in the hepatopancreas was significantly higher than those in the control group when the concentration was higher than 0.50 mg/L (<0.05). CAT activities in the 0.50, 0.90, and 1.70 mg/L groups were significantly higher than those in the control group. The activity of SOD and CAT in the serum first decreased, then increased, and then decreased again with an increasing concentration of nitrite. The activities of AKP and ACP in the serum and hepatopancreas in the 0.90 mg/L group were significantly higher than those in the control group (<0.05). There was no obvious change in MDA content across groups. In addition, serum glutamic-pyruvic transaminase (GPT) activity was significantly higher in the serum than in the control group (<0.05). The results of quantitative real-time PCR showed that the relative expression ofandgenes in the hepatopancreas was significantly higher than that in the control group, except in the 0.50 mg/L concentration group (<0.05). The relative expression of,,,, andgenes in the hepatopancreas was significantly lower than that in the control group (<0.05). After 40 days of challenge with different concentrations of nitrite, the growth and immune function ofclearly diminished. The concentration of nitrite should be controlled below 0.50 mg/L in the production process at a salinity of 5 to ensure the healthy farming of.
; Nitrite; Enzyme activity; Environmental stress
DAI Xilin, E-mail:xldai@shou.edu.cn
10.19663/j.issn2095-9869.20210421001
S917.4
A
2095-9869(2022)04-0180-10
*上海市虾类产业体系建设项目(沪农科[2014]第5号)资助[This work was supported by Shanghai Shrimp Industry System Construction Project (Shanghai Agricultural Science [2014] No.5)]. 方 成,E-mail: 2414100497@qq.com
戴习林,教授,E-mail: xldai@shou.edu.cn
2021-04-21,
2021-06-18
http://www.yykxjz.cn/
方成, 黎兰诗, 梁震宇, 成良峰, 戴习林. 不同浓度亚硝酸盐亚急性胁迫对凡纳滨对虾生长与免疫功能的影响. 渔业科学进展, 2022, 43(4): 180–189
FANG C, LI L S, LIANG Z Y, CHENG L F, DAI X L. Changes in immune system and growth performance ofafter 40-day challenge with different concentrations of nitrite. Progress in Fishery Sciences, 2022, 43(4): 180–189
(编辑 马璀艳)
猜你喜欢
当代水产(2022年8期)2022-09-20
当代水产(2022年5期)2022-06-05
保健与生活(2022年3期)2022-02-11
健康之家(2021年6期)2021-09-08
人人健康(2020年4期)2020-05-25
食品与生活(2018年7期)2018-09-19
家庭科学·新健康(2018年4期)2018-05-14
漫画月刊·哈版(2017年7期)2018-01-29
恋爱婚姻家庭·养生版(2016年11期)2016-11-03
恋爱婚姻家庭·青春(2016年7期)2016-07-05
