
利用F-MSAP分析菜心表观遗传多样性
2022-09-07 08:53史卫东
广西植物 2022年8期
史卫东
( 广西壮族自治区农业科学院蔬菜研究所, 南宁 530007 )
菜心是起源于中国南方的十字花科蔬菜,复种指数高,种植面积大,品种繁多。菜心种质资源狭窄,开展种质创新及鉴定对品种选育很重要。目前,基因组分子标记广泛应用于菜心遗传多样性分析,ISSR、SCoT和AFLP分析表明菜心遗传多样性较低(孙雪梅等,2010;Shi et al., 2011;史卫东等,2015),AFLP和SCoT分析表明菜心遗传变异主要来源于种内(Shi et al., 2011;史卫东等,2015)。菜心分子标记分类与传统分类的一致性和差异性并存,SRAP多态性的聚类分析结果与基于表型特征的分类结果基本一致(李桂花等,2012),SCoT标记的分类结果与熟期分类结果比较一致(史卫东等,2015),但分子标记聚类分析与表型分类也存在不一致的情况(孙雪梅等,2010;李桂花等,2012),说明菜心除了具有DNA序列变化之外,还具有不依赖于DNA序列的表观遗传变化,这种表观遗传变化并不能被基因组标记检测出来,因此,有必要开展菜心表观遗传多样性的研究,以提高鉴定效率和准确性。
DNA甲基化不改变DNA序列的遗传修饰,在减数分裂和有丝分裂阶段可以稳定遗传(Kakutani et al., 1999),因而成为植物最重要的表观遗传标记。DNA甲基化可以发生在所有序列环境中,包括对称的CG和CHG序列以及非对称的CHH序列(Chan et al., 2005),检测CG序列的甲基化状态常用甲基化敏感扩增多态性(methylation-sensitive amplification polymorphism,MSAP),MSAP技术以其简单可靠及费用较低等优点,已广泛应用于拟南芥、甘蓝、芥蓝和油菜等十字花科植物的DNA甲基化分析(Cervera et al., 2002;陆光远等,2005;Salmon et al., 2008;史卫东等,2012;Zhang et al., 2013),基于荧光标记引物扩增的F-MSAP也已应用于辣椒、鸡和牡蛎等动植物的DNA甲基化分析(徐青等,2005;姜群等,2014;徐小万等,2021)。
本研究旨在通过检测49份菜心的DNA甲基化水平和模式变化,揭示菜心表观遗传多样性形成的机理,为进一步提高杂交育种鉴定准确性和效率提供了理论基础和技术支持。
1 材料与方法
1.1 材料
49份菜心为广西壮族自治区农业科学院蔬菜研究所收集和创制的品种资源和自交系,包括7份自交系亲本(1号、2号、3号、5号、6号、7号、14号),8份双自交系杂种(21号、24号、26号、31号、34号、36号、47号、48号),17份单自交系杂种(15号、18号、19号、20号、22号、23号、25号、29号、30号、32号、33号、35号、37号、39号、43号、44号、49号),7份商品种(4号、8号、9号、10号、11号、12号、13号),10份商品种杂种(16号、17号、27号、28号、38号、40号、41号、42号、45号、46号),共计14份品种和35份杂种。田间试验于2014年在广西壮族自治区农业科学院里建科学研究基地进行,每份资源小区面积5 m,采收期随机选取5株,调查记载形态指标,取嫩叶混合冻存于-20 ℃。
1.2 方法
基因组DNA的提取:采用CTAB法提取基因组总DNA。F-MSAP:Taq DNA 聚合酶、缓冲液、dNTP和荧光引物均由北京鼎国昌盛生物技术有限责任公司提供。酶切连接一步法反应体系:DNA(50 ng·μL)4 μL,Adapter 1 μL,EcoR I/MspⅠ或 EcoRⅠ/HpaⅡ 2 μL,10 X Reaction buffer 2.5 μL,10 mmol·LATP 2.5 μL,T4 Ligase 1 μL,HO 7 μL。分别用EcoRⅠ/MspⅠ组合、EcoRⅠ/HpaⅡ组合对同一基因组DNA酶切,混匀后离心数秒,37 ℃保温5 h,8 ℃保温4 h,4 ℃过夜。预选扩增反应:反应体系25 μL为DNA 2 μL、预扩增引物1 μL、dNTPs 0.5 μL、10 X PCR buffer 2.5 μL、Taq 酶0.5 μL、ddHO 18.5 μL。离心数秒,反应条件:94 ℃ 2 min;94 ℃ 30 s,56 ℃ 30 s,30个循环;72 ℃ 80 s,72 ℃ 5 min;4 ℃。选择性扩增反应:预选扩增产物1∶20稀释后作为选扩增模板。反应体系25 μL:DNA 2 μL,10 X PCR buffer 2.5 μL,dNTP 0.5 μL,EcoR I 引物1 μL,HpaⅡ/ MspⅠ引物1 μL,Taq酶0.5 μL,ddHO 17.5 μL。反应条件:94 ℃ 30 s,65 ℃ 30 s,72 ℃ 80 s;每轮循环温度递减0.7 ℃,12个循环;94 ℃ 30 s,55 ℃ 30 s,72℃ 80 s,23个循环;72 ℃ 10 min,4 ℃。接头及引物序列见表1。
表 1 MSAP扩增接头和引物序列(HM: Hpa II/Msp I)Table 1 Adaptors and primer sequences used for MSAP amplications (HM: Hpa II/Msp I)
数据读取及统计分析:将选择性扩增产物进行聚丙烯酰胺凝胶电泳,使用ABI377测序仪检测,通过GENESCAN软件分析,按照“无带为0,有带为1”记录数据矩阵。利用软件Excel 2007计算甲基化多态性比例和甲基化类型,将DNA甲基化模式分为四种类型:Ⅰ型为HpaⅡ和MspⅠ双酶切的(1,1),表示CCGG位点未甲基化;Ⅱ型为HpaⅡ酶切,MspⅠ不能酶切的(1,0),表示CCGG位点C外侧半甲基化;Ⅲ型为HpaⅡ不能酶切,MspⅠ酶切的(0,1),表示CCGG位点内侧C全甲基化;Ⅳ型为HpaⅡ和MspⅠ都不能酶切的(0,0),表示抑制全甲基化CCGG位点酶切,也可能是突变位点。利用软件POPGENE 1.32进行表观遗传多样性分析,统计多态性位点百分率(P%)、Nei’s 基因多样性指数、Shannon 多样性指数、Nei’s 遗传距离、遗传相似系数和基因流等。利用软件MEGA 4.0按照UPGMA方法进行聚类分析。利用软件GenAlEx 6.41进行主成分分析和AMOVA分析。引物多态信息量 (polymorphism information content, PIC),公式为=1-∑,式中表示第个基因型频率。
2 结果与分析
2.1 DNA甲基化多态性分析
利用8对条带清晰和多态性较好的引物进行49份菜心的F-MSAP扩增,一共扩增出1 728条带,其中多态性条带1 479条,多态性比例为86%。8对引物的多态性条带分别为196、186、200、173、188、200、200、178条,平均为190条,多态性比例为88%,PIC值分别为0.230 4、0.201 2、0.247 8、0.237 3、0.244 7、0.241 8、0.234 0、0.224 6,均值为0.232 7,多态性位点的PIC值均位于0~0.5 之间。按照总平均、自交系、品种和杂种进行分类分析,49份菜心、7份自交系亲本、8份双自交系杂种、17份单自交系杂种、7份商品种、10份商品种杂种的平均多态性分别为68.15%、65.33%、68.55%、67.25%、69.54%、70.09%,表明F-MSAP检测效率较高,菜心DNA甲基化多态性较高,商品种的DNA甲基化多态性比自交系及其杂种高,杂种的DNA甲基化多态性比亲本高,杂交可以提高DNA甲基化多态性。
2.2 表观遗传多样性分析
遗传多样性分析显示,49份菜心的平均表观观察等位基因数、有效等位基因数、Shannon多样性指数、期望杂合度分别为1.702 0、1.201 0、0.142 7、0.241 0,7份自交系亲本分别为1.668 0、1.190 0、0.135 4、0.228 9,8份双自交系杂种分别为1.682 9、1.188 9、0.135 4、0.230 1,17份单自交系杂种分别为1.707 5、1.212 0、0.148 7、0.249 1,7份商品种分别为1.729 5、1.202 0、0.143 5、0.243 4,10份商品种杂种分别为1.712 1、1.200 0、0.143 4、0.243 2。结果显示,双自交系杂种的Shannon多样性指数和期望杂合度分别等于和大于自交系亲本,单自交系杂种两者均大于自交系亲本,商品种与商品种杂种两者变化很小,表明菜心表观遗传多样性较低,自交系杂种的表观遗传多样性比亲本高,单自交系杂种的表观遗传多样性比双自交系杂种高,杂交能够增加自交系杂种的表观遗传差异。49份菜心、7份自交系亲本、8份双自交系杂种、17份单自交系杂种、7份商品种、10份商品种杂种的表观遗传距离分别为0.009 4、0.009 5、0.009 4、0.009 4、0.008 6和0.009 6,结果显示,自交系亲本之间的表观遗传距离大于自交系杂种和商品种,单、双自交系杂种之间相同,但均小于商品种,商品种杂种大于商品种,表明自交增加自交系亲本的表观遗传距离,杂交增加商品种杂种的表观遗传距离。AMOVA分析表明,表观遗传变异主要来源于种内(96%),种间较少(4%)(=0.036)。基因流为8.681 5,大于1,表明菜心均质化严重,遗传分化受到抑制,大部分遗传变异来源于种内,只有少量的遗传变异存在于种间。
2.3 DNA甲基化分析
由表2可知,DNA甲基化水平分析显示的49份菜心、7份自交系亲本、8份双自交系杂种、17份单自交系杂种、7份商品种、10份商品种杂种的甲基化率分别为68.14%、65.26%、68.99%、67.39%、69.86%、69.29%。DNA甲基化模式分析显示的未甲基化率分别为31.86%、34.74%、31.01%、32.61%、30.14%、30.71%,半甲基化率分别为33.18%、32.54%、37.80%、31.09%、33.01%、33.67%,全甲基化率分别为34.96%、32.72%、31.20%、36.30%、36.85%、35.62%。结果显示,49份菜心的DNA甲基化水平较高,全甲基化水平高于未甲基化和半甲基化,以全甲基化模式为主,自交系杂种的DNA甲基化水平比亲本高,其中7份自交系未甲基化水平升高导致甲基化水平降低,8份双自交系杂种去甲基化水平升高,17份单自交系杂种全甲基化升高造成甲基化水平升高,7份商品种和10份商品种杂种的DNA甲基化水平和模式变化很小,表明自交能够降低DNA甲基化水平,杂交通过DNA甲基化模式变化增加了自交系杂种的DNA甲基化水平。
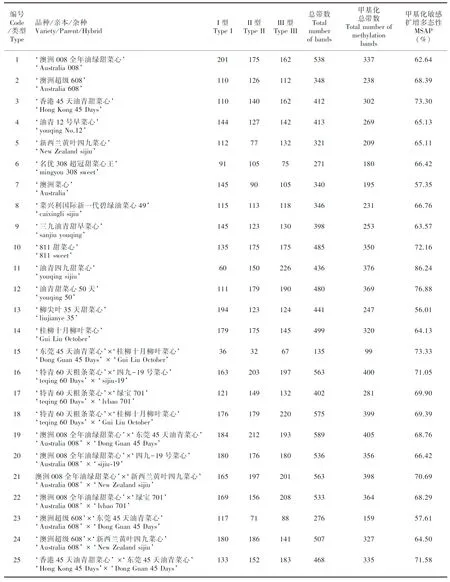
表 2 49份菜心的表观遗传多样性以及DNA甲基化模式和水平Table 2 Epigenetic diversity and DNA methylation pattern and level in 49 Chinese flowering cabbages
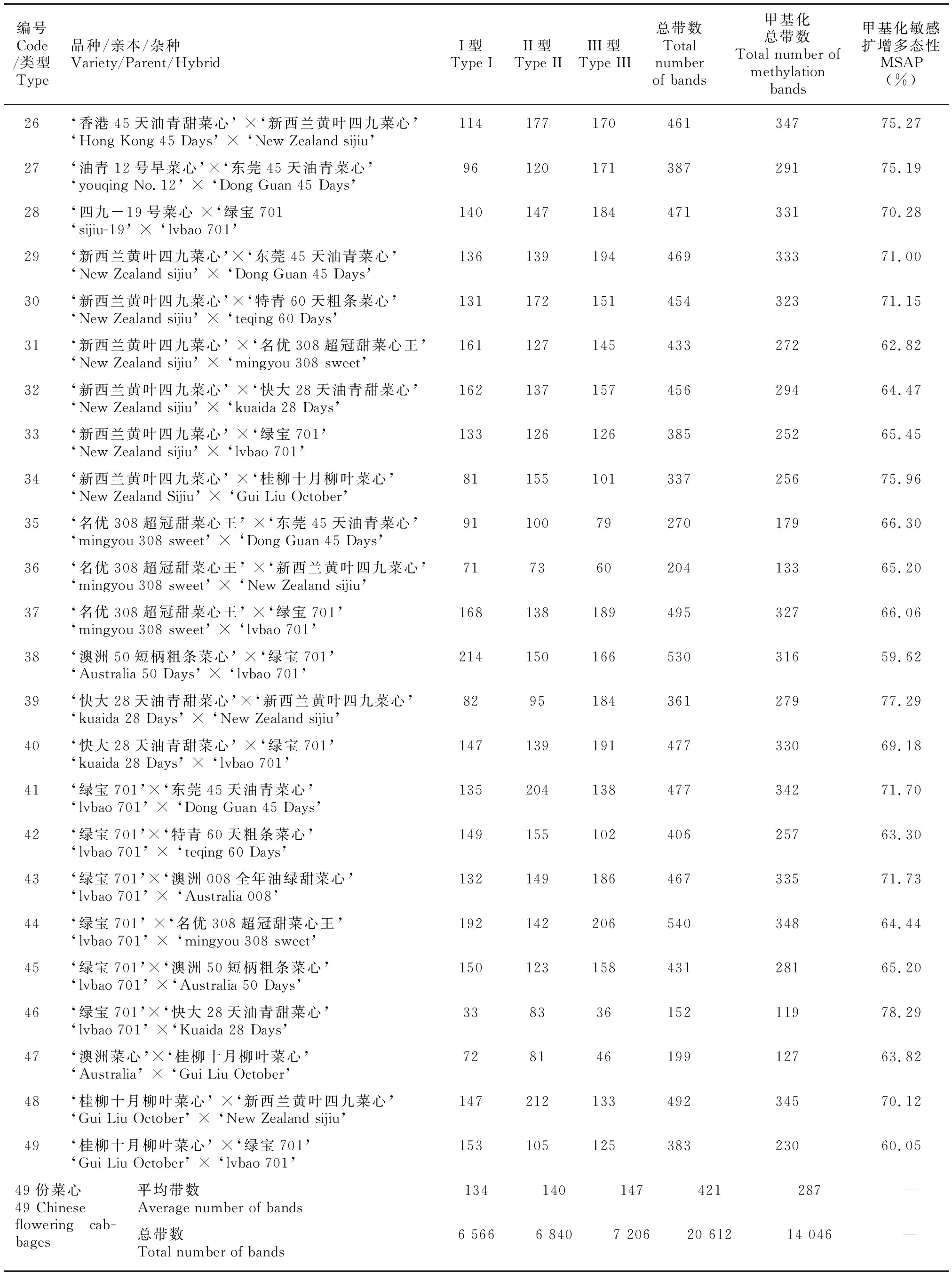
续表 2

续表 2
2.4 聚类分析
如图1所示,利用UPGMA聚类分析方法,约在Nei’s 遗传距离0.42处将49份菜心分成五类。第一类7份包括1号、16号、18号、19号、20号、21号和22号,其中1号是19号、20号、21号、22号的母本,‘特青60天粗条菜心’是16号、18号的母本。第二类2份包括38号和44号,‘绿宝701’分别是38号的父本和44号的母本。第三类14份包括2号、5号、6号、8号、9号、10号、11号、15号、23号、35号、36号、39号、46号和47号,其中2号是23号的母本,6号是35号、36号的母本,‘桂柳十月柳叶菜心’是15号和47号的父本。第四类11份包括12号、14号、17号、24号、37号、40号、41号、42号、43号、45号和48号,其中‘绿宝701’是41号、42号、43号和45号的母本,‘绿宝701’是17号、40号和37号的父本,14号是48号的母本。第五类15份包括3号、4号、7号、13号、25号、26号、27号、28号、29号、30号、31号、32号、33号、34号和49号,其中3号是25号、26号的母本,4号是27号的母本,5号是29号、30号、31号、32号、33号和34的母本,‘绿宝701’是28号、33号和49号的父本。

具体编号见表2。下同。See Table 2 for specific numbers. The same below.图 1 基于遗传距离的49份菜心的UPGMA聚类图Fig. 1 UPGMA dendrogram of 49 Chinese flowering cabbages based on Nei’s genetic distance
14个品种分散在各类中,35份杂种倾向于按照母本亲缘关系分类,表明杂种更多地遗传了母本的DNA甲基化状态,母本DNA甲基化对杂种表观遗传多样性具有较大影响。
2.5 主成分分析
利用GenAlEx 6.41软件包中的PCA模块进行主成分分析(图2),结果显示49份菜心明显分为I、II、III、IV和V五组,五组分布与聚类分析的五类结果基本一致,表明F-MSAP检测效率很高,准确性也很高。第一和第二主坐标的贡献率分别为19.44%和11.81%,可解释31.25%的表观遗传变异。
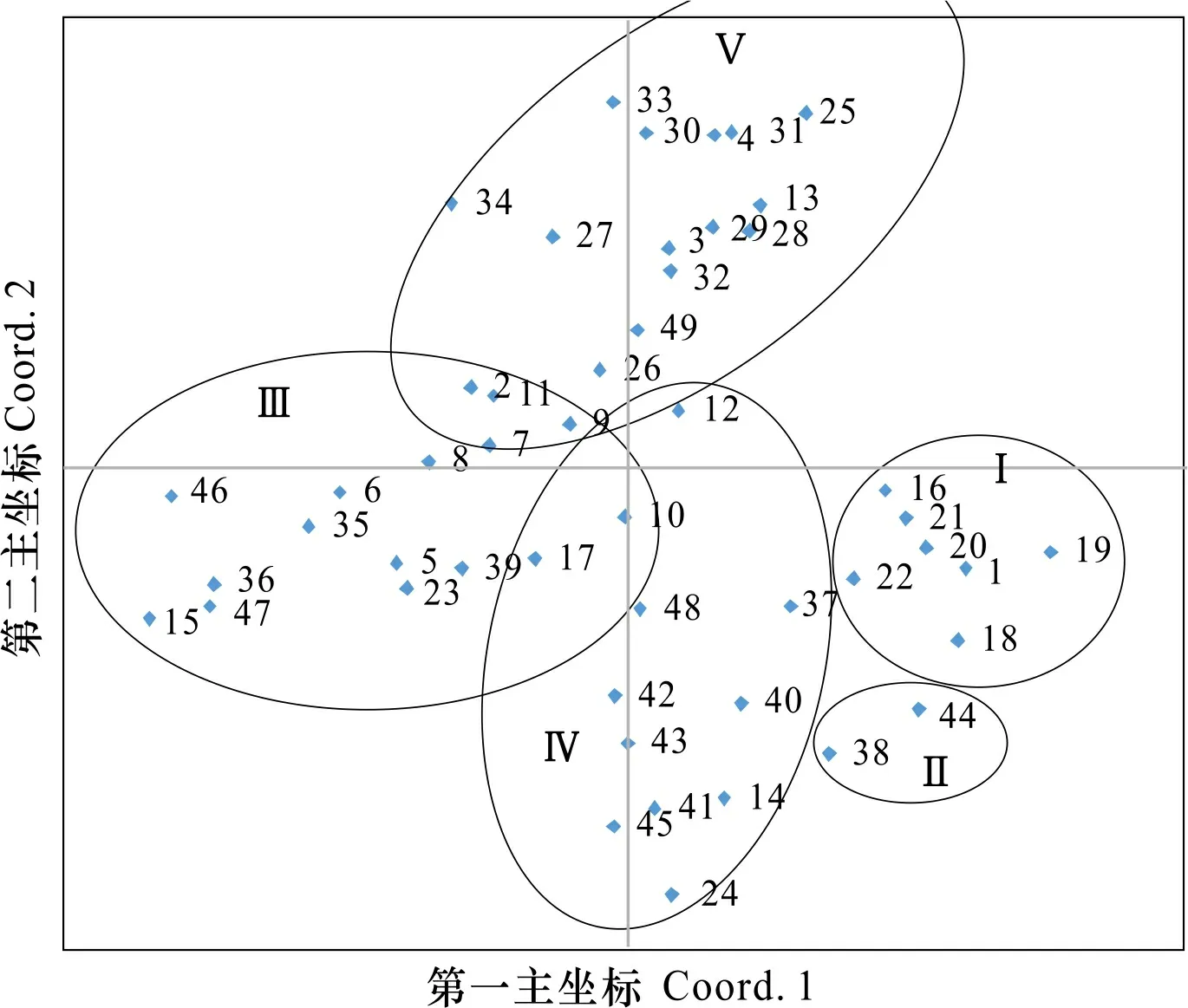
图 2 49份菜心主成分分析Fig. 2 Principal coordinates analyses(PcoA) of 49 Chinese flowering cabbages
3 讨论与结论
3.1 DNA甲基化多态性分析
经典的MSAP利用聚丙烯酰胺凝胶电泳检测DNA甲基化多态性,F-MSAP则是利用荧光标记引物结合毛细管电泳检测,因此检测效率和灵敏度得以较大提高,徐青等(2005)利用F-MSAP检测发现鸡F代的甲基化多态模式约有95%来自亲本,变异的甲基化位点约5%,姜群等(2014)发现F-MSAP检测牡蛎的效率是MSAP的2倍以上,多态性检测率提升了9%。F-MSAP不足之处是只能识别基因组的 CCGG 位点,不能检测CHG序列以及非对称的CHH序列,实际的甲基化水平有所低估(McClelland et al., 1994; Salmon et al., 2008),另外MSAP基于AFLP技术开发,检测的多态性片段大小有所限制。与十字花科亲缘植物相比,49份菜心的DNA甲基化多态性较高,高于拟南芥(24%~34%)、白菜幼苗茎尖(30.42%)、甘蓝(53.3%~60.7%)、油菜种子(15.7%)、芥蓝(47%)、甘蓝型油菜基因渗入系(33.4%~39.8%)的DNA甲基化多态性 (Cervera et al., 2002;Li et al., 2002;陆光远等,2005;Salmon et al., 2008;史卫东等,2012;Zhang et al., 2013),也高于菜心ISSR(56.31%)、SRAP(40.2%)、SCoT(36%)的基因组多态性(孙雪梅等,2010;李桂花等, 2012;史卫东等,2015)。与菜心AFLP和SCoT分析相比,F-MSAP进一步提高了鉴定效率和准确性(Shi et al.,2011;史卫东等,2015),主成分分析也比较一致,表明F-MSAP可以作为一种检测菜心DNA甲基化的有效方法,极大地提高菜心DNA甲基化的检测效率和灵敏度。
3.2 表观遗传多样性分析
Shannon多样性指数是评价种内和种间遗传多样性水平的指标,其值越大表示遗传多样性越高,49份菜心的表观Shannon多样性指数(0.142 7)小于基因组ISSR(0.229)、AFLP(0.472)和SCoT(0.217)的Shannon多样性指数(孙雪梅等,2010;Shi et al., 2011;史卫东等,2015),表观遗传距离(0.009 4)小于ISSR(0.029~0.344)、AFLP(0.112)、SCoT(0.428)等基因组标记的遗传距离(孙雪梅等,2010;Shi et al., 2011;史卫东等,2015),据此推测菜心表观遗传多样性较低且低于基因组遗传多样性,与芥蓝的表观遗传多样性水平较低的结果相一致(史卫东等,2012),与西瓜、水稻和辣椒的基因组甲基化多样性高于遗传多样性的结果不同(Nimmakayala et al., 2011;彭海等,2014;徐小万等,2021),这可能与不同的比较方法有关,也可能与不同物种之间表观遗传多样性和基因组遗传多样性存在较大的差异有关。本研究还发现菜心自交系亲本的表观遗传距离大于商品种,表明自交增加了表观遗传多样性水平,丰富了遗传背景,商品种表观遗传多样性最低,但商品种杂交增加了表观遗传多样性。
3.3 DNA甲基化分析
研究表明在不同生态型拟南芥上35%~43% 的 CCGG位点为DNA甲基化敏感位置,而在同一生态型内则高度保守(Cervera et al., 2002),甘蓝品种或品系具有更多的甲基化片段(Salmon et al., 2008),甘蓝型油菜基因渗入系呈现高频率的高甲基化(Zhang et al., 2013)。本研究菜心自交系亲本、自交系杂种、商品种和商品种杂种的未甲基化、半甲基化和全甲基化比例各占1/3以上,但全甲基化在商品种较高,表明菜心以全甲基化模式为主,菜心与拟南芥、甘蓝具有相似的甲基化变化,这是否是十字花科植物的普遍现象,还有待研究。
菜心自交系的DNA 甲基化水平降低,与大白菜自交系的 DNA 甲基化水平降低相一致(Liu et al., 2018),从甲基化组角度说明菜心与大白菜亲缘关系较近,因而具有相似的表观遗传变化。拟南芥杂种的甲基化模式变化经常发生在亲本之间的差异甲基化区域(Greaves et al., 2012),亲本之间的甲基化差异可能是亲本和杂种之间甲基化差异的主要原因(Shen et al., 2012)。菜心单自交系杂种和双自交系杂种的甲基化水平都比亲本高,与拟南芥 C24/Landsberg F杂种的甲基化水平增加相一致(Greaves et al., 2012),但两个杂种的DNA甲基化模式变化有所不同,因此造成亲本自交系和商品种的DNA甲基化差异。菜心自交系随着不断自交纯合,基因组杂合性减低,表现为DNA甲基化水平降低和未甲基化水平升高,菜心商品种为开放授粉种子,基因组杂合性较高,表现为全甲基化水平较高。据此推测菜心单自交系杂种的全甲基化水平较高可能是由于自交系与商品种杂交造成的,与拟南芥正反交 F中一个亲本等位基因的甲基化水平改变为另一个亲本甲基化水平有相似之处(Greaves et al., 2012)。菜心双自交系杂种表现为高水平的去甲基化,与小麦黑麦远缘杂交后代的半甲基化水平极显著高于双亲本的半甲基化水平有相似之处(朱朝阳等,2018)。DNA去甲基化的功能和机制是生物学倍受争议的研究领域,活跃的DNA去甲基化对修剪基因组的甲基化模式很重要,甲基化和去甲基化的动态调控对于保持植物表观基因组的可塑性也很重要(Zhu et al., 2007)。菜心亲本差异、自交或杂交引起后代DNA甲基化模式和水平变化,表明菜心基因组可塑性可能很强,表观遗传多样性形成机理还有待深入研究。
3.4 杂交对表观遗传多样性的影响
表型与分子标记都可以检测菜心亲缘关系,但两者的鉴定结果并不十分相符,可能的原因包括品种资源的遗传背景比较复杂,基因组分子标记优化不足或不适合,以及不能检测出表观遗传变化等。本研究49份菜心的聚类结果清晰可靠,14份菜心品种呈现较大的表观遗传变化,其中7份自交系亲本分散在各类中,与自交增加了表观遗传距离相一致,说明自交增加了自交系亲本的表观遗传差异。研究表明拟南芥互交杂种具有完全相同的遗传构成,杂交后母本和父本基因组可能不会平等地影响杂种DNA甲基化的变化,杂种的甲基化重塑可能有利于杂种优势(Shen et al., 2012)。本研究35份自交系杂种和商品种杂种分布在各类中,倾向于按照母本亲缘关系分类,与杂交稻甲基化状态与母本较近的结果相同(彭海等,2014),与拟南芥不同基因型的甲基化多态性不相关,亲缘关系紧密的拟南芥并不归为一类的结果不同(Cervera et al., 2002),与拟南芥的基因甲基化与亲缘关系聚类分析结果不相关的结果不同(Matthew et al., 2007),菜心母本亲缘关系可能对杂种表观遗传多样性具有较大影响,因此在利用不同菜心亲本配组时,要注重对母本的选择,依据母本亲缘关系对杂种后代分类可能会起到事半功倍的效果,对菜心种质资源研究和育种具有极其重要的指导意义。
综上所述,通过F-MSAP检测菜心DNA甲基化变化,揭示了菜心表观遗传多样性形成机理,分析和预测了杂种后代的表观遗传差异,提高了鉴定效率和准确性,为进一步开展杂交育种提供了理论基础和技术支持。
猜你喜欢
江苏农业科学(2022年6期)2022-04-15
三农资讯半月报(2020年13期)2020-07-31
福建基础教育研究(2019年10期)2019-05-28
农民致富之友(2017年10期)2017-06-15
农民致富之友(2016年5期)2016-10-21
江苏农业科学(2016年4期)2016-06-14
广西农学报(2015年3期)2015-10-14
长江蔬菜·学术版(2014年4期)2014-05-09
