
云南东部江川生物群中的香肠状疑难化石
——Tawuia化石属性的新解释
2022-09-26 06:17李玉兰王浩刘傲然李明梁永忠周颖唐烽任留东
地质论评 2022年5期
李玉兰,王浩,刘傲然,李明,梁永忠,周颖,唐烽,任留东
1)贵州大学资源与环境工程学院,中国贵阳,550025;2)自然资源部地层与古生物重点实验室,中国地质科学院地质研究所,中国北京,100037;3)贵州省地矿局111地质大队,中国贵阳,550081;4)河北地质大学地球科学学院,中国石家庄,050031;5)云南云天化环保科技有限公司,中国昆明,650300;6)伦敦大学学院地球科学系, 英国伦敦,WC1E6BT
内容提要:云南东部伊迪卡拉(震旦)系顶部灯影组旧城段的江川生物群是华南又一大类丰富且多样化的宏体化石群,其以碳质压膜的方式保存。在该生物群中,我们发现大量香肠状碳质压膜的宏体化石,与成冰(南华)纪全球大冰期之前中、新元古代地层中广泛分布的Tawuia碳膜化石的大小、形态非常相似。其最大宽度达4.5 mm,最大长度可达4 cm,长宽比均>2,膜面光滑,未见其他稳定纹饰,有的化石标本边缘碳质保存较多,可见深黑色边缘环带。但部分化石保存的形态与常见的直棒状I形、U形、C形Tawuia化石相比却较为特殊,呈现直角L形、8字形、宽U形、环状O形等多种保存形态,个别标本弯折部位可见明显的碳质增厚和横脊纹残留,边缘残存有大小不一的刺状突起构造。这类特异形态保存的似Tawuia化石显然不能解释为以往较为公认的浮游型叶状体藻类,而可能与冰期前的新元古代早期鲁西、淮南宏体化石群中的疑似蠕形动物亲缘的化石有传承关系。上述新发现的化石形态表明:似Tawuia的碳膜宏体化石可能是多源属性的早期多细胞生物。其中短带至长带状叶状体,可见短柄突起及疑似固着器构造的化石可确认为底栖宏体藻类;多数的叶状体推测为香肠状或长囊状,生活史中包含远洋浮游的类型,可能归属于原始不分枝的多核体藻类或早期分化出的中空囊状的多细胞藻类;本文展示的部分规则变形的似Tawuia化石,更可能是两侧对称动物亲缘的,生活于近海的底栖生物体发生主动扭曲后,被特异埋藏所呈现出的特殊保存形态,而随机浮游或表栖的多细胞藻类死亡沉积时不会形成类似形态的化石。
Tawuia化石最早由Hofmann依据加拿大麦肯齐山前寒武系小达尔群(Little Dal Group)产出的毫米级以上肉眼可见的一类宏体化石而命名为Tawuiadalensis,原始的属征记述一般为两端封闭呈半圆形的似棒状、香肠状,层面上保存为条带状的碳质压膜化石,边缘轮廓光滑连续,两侧平行或至一端略成锥状,膜面平坦无饰或在端部少数可见同心圆状构造及深黑色的轴线条带(Hofmann and Aitken, 1979),但当时所指定的模式种则呈现中部强烈弯曲的U形。而在随后的修订中,Hofmann(1985)将这类化石中多有发现的向一侧弯曲保存为C形、U形和表面出现边缘褶皱作为原生构造纳入特征描述。
20世纪70年代以后,中国学者也陆续发现在华北燕山地区和华北东缘胶辽徐淮地区的新元古代成冰系上、下地层中广泛出露与TawuiadalensisHofmann and Aitken, 1979相似的宏体化石,并将其中平直保存的短条/短棒状化石划分出来命名成新类别TawuiasinensisDuan, 1982,也曾将发现于华南西南缘昆明地区的新型梭状化石命名成新类别TawuiafusiformisXing, 1985,但后者一直没有发表规范的新种描述。其后,又记述了在河北怀来的龙凤山生物群(杜汝霖和田立富, 1985; 刘傲然, 2018)、苏皖北部的淮南生物群(符俊辉, 1989; 钱迈平等, 2009)、辽宁大连的辽南生物群(洪作民等, 1986, 1991)、海南石碌的Chuaria—Tawuia生物群(张仁杰等, 1989, 1991)和北京地区青白口纪(唐烽, 1995)所发现的Tawuia类化石。近年来,在鲁西石旺庄组及淮南寿县刘老碑组也新发现并对比研究了保存更加完美的Tawuia化石(Li Guangjin et al., 2020; Tang Qing et al., 2021a)。
而在华南扬子地层区的伊迪卡拉(震旦)纪地层中,除了20世纪80年代开始在云南昆明晋宁区的灯影组旧城段发现了形态可以归属Tawuia的碳膜化石(邢裕盛, 1985),其他地区迄今均未见明确的报道。本世纪之初,唐烽等依据前人的资料线索在云南玉溪江川区伊迪卡拉系近顶部旧城段发掘出丰富多样的宏体化石群——江川生物群(唐烽等, 2006, 2009, 2015; Tang Feng et al., 2007; 顾鹏等, 2018),其中包含的似Tawuia碳膜化石,与全球其他地区产出的相似类型比较,显得更加多样化,本文在此进一步归类记述了4大类型,并依据保存形态特异的部分弯曲化石,对以往普遍认定似Tawuia化石的藻类属性提出质疑和新的解释,讨论对比了华北地区东缘和华南扬子西南缘等地产出的弯曲变形的似Tawuia化石,推测了某些特殊构造的成因。
1 江川生物群Tawuia化石产地及地层分布
研究区位于滇东玉溪地区,位于扬子陆块区之上扬子新元古代—中生代裂陷带。地层区划隶属华南地层大区扬子地层区康滇地层分区之昆明地层小区。研究区出露地层由老到新有前震旦系的澄江组、南沱组及震旦系陡山沱组、灯影组(东龙潭组后称藻白云岩段,与上覆旧城段、白岩哨段共同组成灯影组,白岩哨段顶部部分地区沉积较深水泻湖相黑色硅质页岩,曾经被定义为小歪头山段),寒武系渔户村组(狭义的分为中谊村段和大海段)、筇竹寺组、沧浪铺组、龙王庙组、西王庙组、娄山关组及二叠系阳新组。江川生物群的化石剖面主要位于云南昆明市晋宁区六街至王家湾县道旁的灯影组连续露头剖面和玉溪市江川区桃溪村至古埂,以及江城镇侯家山至清水沟的矿山公路北侧灯影组中上部(图1a)。化石产出层位具有较好的分带性,经过近年来的深入采掘研究,自下而上识别出5个宏体化石组合(唐烽等, 2015; 顾鹏等, 2018),分别是灯影组中部旧城段的A—D组合以及位于灯影组上部白岩哨段的E组合:
A)Shaanxilithes陕西迹化石层,
B)Chuaria—Tawuia—Pumilibaxa化石层,
C)Shouhsienia—Vendotaenia—Palaeocolpomenia* 化石层,
D)Longfengshania—Cycliomedusa*—Eoslivera* 化石层(*即将发表的新类别),
E)Vendotaenia—Tyrasotaenia带藻类化石层。
其中,B—D组合(图1b)富集保存的化石矿坑出露点在侯家山村西南约900 m,海拔2106 m,北纬24°27′39.3″,东经102°46′48.4″。含化石层位岩性下部主要为泥质页岩、泥质白云质粉砂岩互层,向上过渡为泥质粉砂质白云岩,风化后常呈黄灰色,具水平微层纹,应属潮间潮下砂、泥坪环境(罗惠麟等, 1988),碳膜保存的宏体化石在风化的层面上出露明显,个别化石膜面黄铁矿化甚至进一步氧化呈褐红色,非常易于识别。
笔者等特别描述及对比的Tawuia类碳质压膜化石,产自侯家山化石矿坑的下部,即上述旧城段江川生物群B组合中(图1b; Tang Feng et al., 2021)。
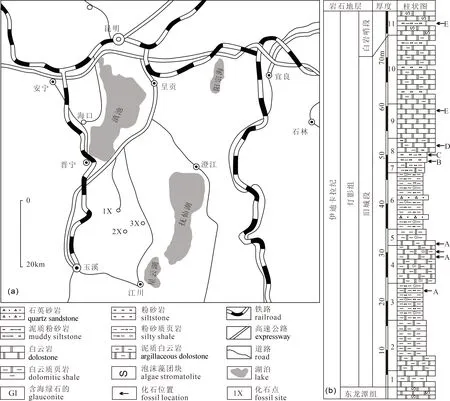
图1 滇东玉溪地区江川生物群化石剖面的地理位置图(a)及地层柱状图(b)(1X—王家湾;2X—古埂;3X—侯家山)
2 江川生物群中多种形态的Tawuia类化石
华南扬子区西南缘震旦纪晚期产出的Tawuia类宏体化石,与华北燕山地区尤其是华北地区东缘全球冰期以前青白口纪保存丰富的Tawuia化石形态相似,类型相当或更为丰富。常见一类压扁的香肠形和或长或短的直棒形及略有变形的哑铃形和鞋履形;第二类披针形为一端略细窄锲状,一端仍渐粗膨大,第三类梭形为两端渐细,中间膨大,但这两类的端部仍然浑圆封闭,符合Tawuia类的属征;第四类则是各种弯曲形,有C形、U形这些华北地区及全球其他产地均出露的形态,也有本地首次发现的更为罕见的L形、O形和8字形。
2.1 香肠形或直棒I形
呈直棒I形的T.sinensis(Duan Chenghua, 1982),是我国华北燕山地区和华北东缘胶辽徐淮地区新元古代早期地层中常见的类型,甚至在河北宽城、兴隆地区的中元古界常州沟组也曾报道过香肠形Tawuia状化石及可能的高级藻类多细胞组织结构(如假薄壁组织等)的发现(Zhu Shixing et al., 2000; 陈孟莪, 2000)。本文化石材料中同样多见直肠状的这类化石,两侧缘略收缩的鞋底形也有产出(图2f, m),且出露了许多形态更大的个体(图2e; Tang Feng et al., 2008, 图5);以及短棒形叶状体出现明显的固着端构造的个体,最近被划分为一新类别“古囊藻”(Palaeocolpomenia, Tang Feng et al., 2022, 图2);和长棒形叶状体出现细长拟茎状突起及纺锤状固着构造的个体,曾被命名为另一新类别“带状江川藻”(Jiangchuaniataeniophylla),推测属于新元古代初期的优势类群龙凤山藻科Longfengshaniaceae化石(Tang Feng et al., 2007, 图1, 17, 20)。尽管目前这些新出现类别的细胞结构仍然不明,但从形态上可以确定叶状体似Tawuia的宏体化石也有部分是底栖固着类型,而非以往认知的与圆盘形Chuaria紧密共生的浮游类型。
滇东旧城段泥质粉砂岩中保存的香肠、直棒形Tawuia状化石表面覆盖有厚薄不均的碳膜,多数膜面光滑无饰,仅环绕膜面周缘呈现一圈碳质增厚的环带,可以解释为生物有机质经埋藏被挤压到周边保存的现象;少数膜面出现裂纹(crack)、孔洞(hole)、皱纹(crinkle),但这些无定向、不规则、不稳定的纹洞(图2,箭头)显然不能代表这类生物的原生构造,而是后期风化及压埋作用所致;个别化石可见端部类似圆孔的开口(图2b,箭头)和局部残存的细密平行的横向纹饰(图2k,箭头),推测是原生构造,可能与部分化石的生物属性有关(见后文讨论)。

图2 滇东玉溪地区江川生物群香肠形或直棒形Tawuia类化石
2.2 披针形
该类化石曾经在苏皖北部的淮南生物群(符俊辉, 1989, 图2)及河北燕山地区的龙凤山生物群(刘傲然, 2018, 图3)中就有过报道,在滇东江川生物群中也很常见(图3)。在Hofmann(1979, 1985)对Tawuiadalensis化石的特征定义中将“有些化石末端变得细尖”即“披针形”包含其中,但在其文献中仅展示了个别这样形态的化石(Hofmann, 1985, 图36-3),而一端逐渐窄细,另一端粗壮膨大,应该是化石生物的原生形态,而不可能是肠形及棒形Tawuia化石压埋变形导致的。滇东这类化石碳膜保存较厚的标本呈现出明显折叠的褶皱纹饰,略见纵向分布,自窄细端向膨大端略有发散,个别可见肋状平行保存(图3a—d, h),这种现象表明化石生物体在膨大端可能为中空的囊状,而在窄细端则渐变为圆筒状,并可能有加长生长为固着端的趋势(图3a)。这种叶状体类似披针形态、在细尖端出现长柄和固着构造的过渡化石(图3),最早在本地报道过的龙凤山藻科化石Longfengshaniaceae中就有所展示(唐烽等, 2006, 图1-1, 2, 5, 7, 14, 19),所以这类Tawuia化石极有可能是一种底栖固着类型宏体藻类的还未分化出固着器官的过渡类型。
计算机信息化教育已经在我国初步发展起来,但仍远远不够。要想深化计算机信息化教育,我们的政府应该详细制定相关的政策并加强引导,支持建立专业计算机信息化教育机构;学校与企业之间应该加强合作;教师更应该转变现有的观念,加强计算机信息化学习,改变教学方式。相信在不远的将来,计算机信息化教育必将成为我国教育的主流模式,提供更多的专业人才,使中国在世界科技之林中立于不败的位置。
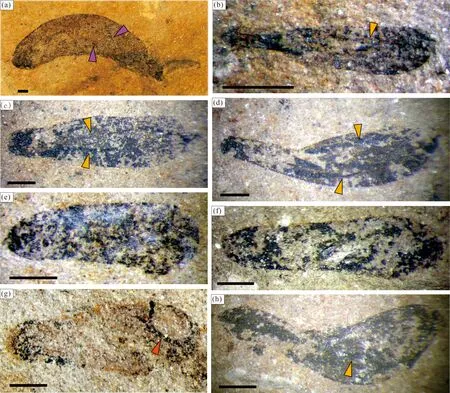
图3 滇东玉溪地区江川生物群披针形Tawuia类化石
2.3 梭形或纺锤形
邢裕盛(1985)最早报道并将滇东产出的梭形Tawuia化石划分为新种Tawuiafusiformissp.nov.,但一直没有进一步描述和图示模式标本,应为无效种名。但近年来的采集研究表明,梭形或纺锤形态的似Tawuia化石在滇东震旦纪旧城段的江川生物群中多有发现,而在其他地层、其他地区中均未见报道,可能是一类分布不广的区域类群。这类化石外表形态与上述的披针形基本类似,但独有的特征就是中间略见粗壮膨大,两端均缩小封闭,保存的碳膜表面也是光滑无饰(图4)。叶状体藻类这样双向对称均匀生长的模式,在前寒武纪宏体生物群中还是首次发现。而现生高级藻类中叶状体也基本呈现一端膨大优势生长的体型,除非在一些扁叶状具中轴的藻类叶状体上可以不规则随机生长出有浮力的膨大囊泡(Tseng, 1983)。推测这类梭形古老化石膨大的中间部分也可能是囊状略微中空的适于远洋漂浮的结构,沉积风化后的化石体两端仍然保留有较厚的碳质压膜,表明藻体两个端部有机质富集,都可能是活跃的生长点,即经过一段时期的光合漂浮生活,任意一端均可能附着在较硬底质上继续底栖阶段的生长。这有助于及时有效地占据透光带的水体中或底质表面上的生态位,可能是藻类演化过程中更为原始的性状特征之一。
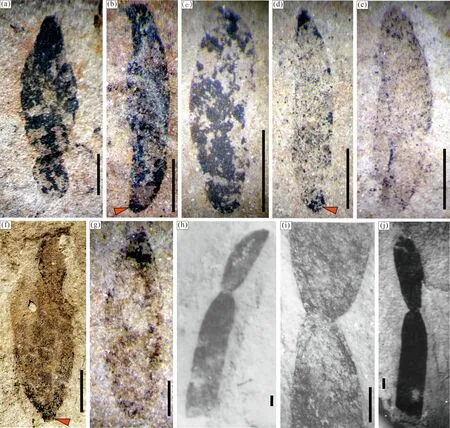
图4 滇东玉溪地区江川生物群梭形或纺锤形Tawuia类化石
唐烽(1997)曾图示出露于安徽淮南寿县刘老碑组黄绿色页岩中富产的宏体藻类化石,这是国内最早发现前寒武纪宏体化石的产地及层位。其中包含很罕见的梭形或纺锤形的Tawuia类化石,个别标本可见大小两个个体,呈藕节状接合在一起保存,观察其接合处,褶皱纹饰在接合部位自然汇聚,未见叠压现象,明显为两个叶状体端部紧密黏合形成(图4h—k)。推测一个可能是藻体沉积保存时,两个体分泌的藻胶黏液恰好黏连在一处,最近的文献(Li Guangjin et al., 2020, 图4p—s)图示了这种可能性;另一可能则是藻体生活史中的某个阶段,小的个体从大的母体顶端增殖出来,生长到一定长度自行脱落,独立生存;上述标本的完整保存,类似现代多年生褐藻的海带类藻体的一种生长方式,由叶状体的端部以中间分生组织生成一新的藻体,自老的藻体上渐渐脱落。本文展示的部分标本一端保存疑似脱落的平坦接合面(图4b, d, f),可以解释为上述藻体增生后脱落为浮游型个体的可能性;从化石出露层位的白云质、泥质粉砂岩发育水平微层理,和上覆薄层粉屑白云岩可见变形层理,反映出潮下低能的斜坡沉积环境(唐烽等, 2007, 图3),而安徽淮南刘老碑组可以肯定是沉积于水动力条件更弱的页岩相静水环境,所以会有梭形Tawuia类化石个体藕节状接合状态的特异埋藏保存。
2.4 弯曲形(C、J、L、U、O、8字形)
以往国内外文献报道的弯曲变形的Tawuia类化石常见有C形和U形,一般是化石体较为细长时保存这种形态的化石较多(Hofmann and Aitken, 1979; Duan Chenghua, 1982, 图4; 符俊辉, 1989, 图1-2; 张仁杰等, 1989, 图2; Singh, 2009, 图4; 钱迈平等, 2009, 图11-6, 8),特别在我国华北东缘新元古代早期地层中出露的更为丰富(Sun Weiguo et al., 1986, 图7; Li Guangjin et al., 2019, 图4c; Li Guangjin et al., 2020, 图4; Tang Qing et al., 2021a, 图2, 图4)。张仁杰等(1989, 1991)报道过海南石碌群碳质板岩中产出的典型案例,图示了很多C形(马蹄铁形)和U形的Tawuia类化石,并统计了两个群类长宽比(L/W)的分布规律,发现的化石几乎都位于长宽比在5~10 之间(张仁杰等, 1991, 图2);较为短粗的圆弧状C形或马蹄铁形与化石体更为细长、两端加长至平行并列甚至末端黏合的U形明显分为不同的两个类群,表明两类均是加长生长的同时也在增宽,而显然在一定的长度和一定的宽度后,C形弯曲变化为U形弯曲,两者不存在个体发育上的成长关系。
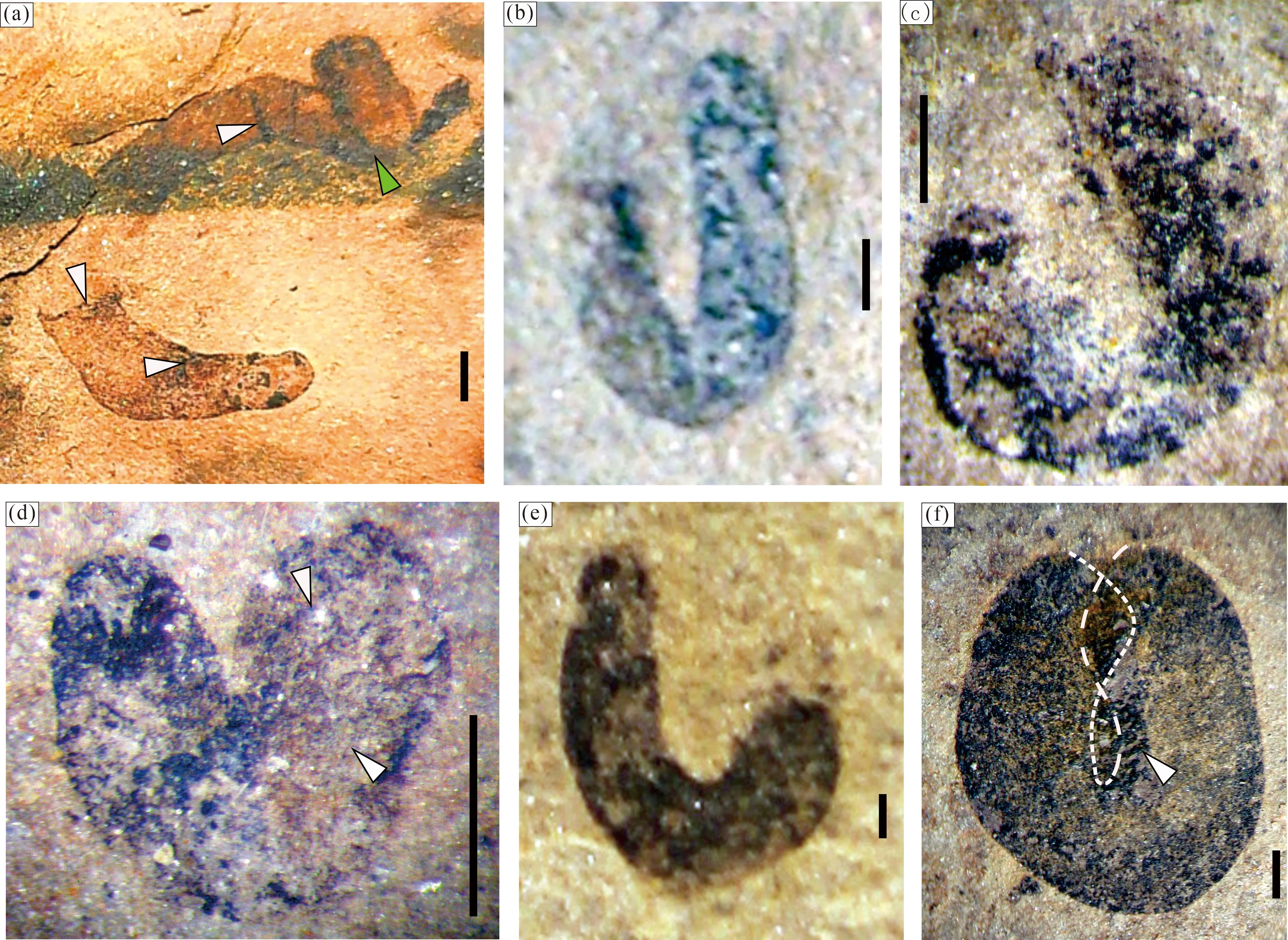
图7 滇东玉溪地区江川生物群中U形的Tawuia类化石
滇东江川生物群中C形和U形的Tawuia类化石并不丰富,没有进一步的统计分析。而且,滇东弯曲形的化石新发现了L形以及罕见的O形和8字形。
C形与直棒状化石(图2)相比多数个体较小,呈腰果状,保存长度5 mm以下,宽度不到2 mm,未见细长如马蹄铁形的化石,曲率小、圆周切角均为钝角(图5);L形略粗长,一般在个体中央位置打折成明显的直角或锐角,而不是圆弧状弯曲,两侧缘并不是一直保持平行延长,保存的宽度在端部可见膨大,个别端部还见到塌陷变形,表明生物体可能是圆筒状体或囊状体(图6);U形化石在江川生物群中更加少见,几乎是偏于一端弯曲的标本,拐点的一端生长较长而一端较短,这与其他地区的弯曲面两侧基本平行对称的U形化石保存不大一样(图7),个别粗壮的标本甚至弯曲成圆饼状,内侧缘部分重叠存留很厚的碳膜(图7f)。

图6 滇东玉溪地区江川生物群中折尺状或L形的Tawuia类化石
O形或环状的化石在滇东产地迄今只发现了4件标本(图8),可识别出是香肠状化石弯曲成闭合的环状保存而成,两端接合处均可见部分重叠,碳质膜面光滑,环状边缘清晰,内、外缘均保存有较厚碳膜,特别是内缘可见碳膜的斜面和横向纹饰,有明显的立体感,表明化石生物弯曲时可能是圆筒形的(图8e,g)。
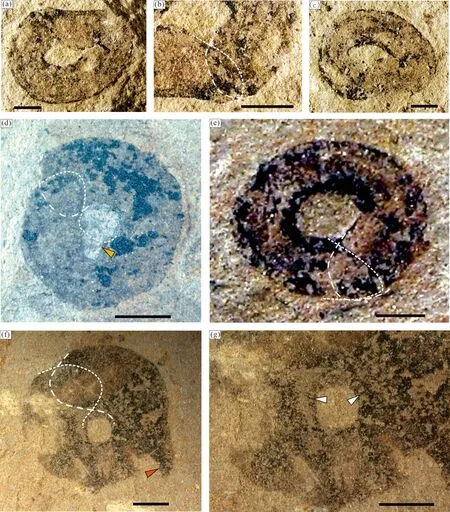
图8 滇东玉溪地区江川生物群中环状或O形的Tawuia类化石
本文记述的8字形化石仅有一例,明显为香肠状生物的两端不是如上述几种都是向一侧方向的弯曲,而是向两侧反向弯曲,保存的化石呈现8字形,一端强烈内卷闭合,中央残存较厚碳膜,另一端基本以相同曲率卷曲但未闭合,化石体边界清楚,弯曲两端部碳膜保存较多(图9a—c)。
此外,滇东化石群弯曲变形的Tawuia类化石中也包含有极少数的一端逐渐窄细而一端膨大浑圆的披针形个体(图9d,e),这也是在其他国内外相关生物群中迄今未见报道过的。

图9 滇东玉溪地区江川生物群中“8”字形和弯曲的披针形Tawuia类化石
3 对比及讨论
3.1 推测有动物亲缘的似Tawuia化石形态学及埋藏学对比
华北燕山地区发育大量的呈直棒形的T.sinensis(Duan Chenghua et al., 1982),很少发现其他变形弯曲的标本(刘傲然, 2019),更未见原始定义的较强烈弯曲呈“U”形的T.dalensis(Hofmann and Aitken, 1979)。而在华南的海南石碌群曾发现更多的Tawuia类化石呈“C”形或“U”形弯曲的种,比例较大(约84%; 张仁杰等, 1991),个体超大,特别是“U”形化石很多,且多数在化石体中部强烈弯曲,致使两侧躯体并列靠拢甚至接触叠压,在层面上更是不规则、无定向分布,非常醒目。对此,尽管更早的资料都将这类U形管化石称作“石碌虫”,但原作者(张仁杰等, 1989, 1991)显然默认了原命名者(Hofmann, 1985)将此弯曲类型作为主要原生特征归入宏观藻类的观点,而没有进一步做埋藏统计学的研究。苏皖北部的淮南生物群中也以个体很大的“C”形或“U”形弯曲的种T.dalensis最为发育, 许多碳膜标本可见明显的末端小圆盘及局部(近端部或弯折部)表面具臼齿状横肋和细密的横纹(钱迈平等, 2009; Li Guangjing et al., 2020; Tang Qing et al., 2021a);但早有学者认为具横向纹饰的化石,其大小和形态更加接近现代绿藻类中的蠕环藻(Neomerisannulata),而所有的弯曲形态都被推断是因埋藏状况呈现的各种变形(钱迈平等, 2009)。
现代藻类中圆筒状、长囊状且具横向环纹的种类非常罕见,浅水潮下带易钙化的蠕环藻(N.annulata)是最典型的类型之一(Tseng, 1983),该藻最早的化石记录尚不清楚。而现生最常见的具环纹蠕形生物的最早代表在寒武纪早、中期的云南澄江动物群和加拿大布尔吉斯页岩化石群就有所记录(Briggs et al., 1994; Hou Xianguang et al., 2017),现生典型种类是环节动物门的蚯蚓。其在生活状态下自主蜷曲的形态与本文引述、描述的弯曲型Tawuia化石相当(图10a—h),特别是与江川生物群中的小微个体出现的8字形和U形等及弯曲处具细密横纹的保存特征比较一致。由此,笔者等赞成前人提出的新元古代大冰期前即已广泛出现、形态构造简单的Chuaria—Tawuia组合可能是多源的,尤其是依据原始定义特征的Tawuia极可能是一个“大口袋”形态属(郑文武等, 1994; 陈孟莪, 2000),其亲缘属性复杂,可能更具多解性。其中许多弯曲形态的原定为多细胞宏体藻类埋藏变形的、近来又被认为是“巨细胞”多核体绿藻类的Tawuiadalensis(Li Guangjin et al., 2020; Tang Qing et al., 2021b),目前来看,更有可能具管状的蠕形动物亲缘。其一,在国内外主要的宏体化石群中产出的似Tawuia化石,如果是随机飘浮或表栖的多细胞藻类,必然在定向水流作用下,沉积时才可能呈现出特殊的弯曲保存形态;而在海南石碌、华北东缘的苏皖及鲁西和加拿大等地出露的类似形态的化石,在层面上却是杂乱分布,没有显示出任何定向的特点;化石富集层位也是页岩、泥岩或粉砂岩性及具水平微层理的静水或弱水动力沉积环境。其二,石碌的巨大弯曲的Tawuia化石丰度更是比直棒状的多出约4倍,应该不可能是同沉积层中直棒状化石受埋藏变形影响的结果,且化石富集层的石碌群第6层碳质板岩和其他几个著名产地化石层位的泥质/碳质页岩中均没有定向水动力的沉积记录。其三,中国石碌、淮南和加拿大小达尔的生物群中均出现较多的弯曲变形T.dalensis化石,只是大多数标本弯曲曲率最大点都在化石体中部,强烈弯曲还导致两端躯体平行甚至端部靠拢叠压,这种稳定姿态的保存方式是直棒状个体死亡后经水流冲刷、随机埋藏成因难以解释的;其四,江川生物群中化石B组合的产出层位岩性为粉砂质、泥质页岩和白云质、泥质粉砂岩互层,相邻层位存在陕西迹化石Shaanxilithes富集的泥岩和包卷层理的记录(图1b; 唐烽等, 2007),从化石组合变化和岩性过渡都表明是向上缓慢海侵渐次水深的弱水动力沉积环境,但产出的U型等多种形态弯曲变形的Tawuia类化石(本文图5—图9)明显微小,丰度也少,与香肠状、鞋屐状较大化石相比差别甚巨(图2~图4);所以,至少在江川生物群中这些变形的类别不大可能是死亡沉积的藻类被动扭曲后埋藏,更可能是小型或幼年管状或香肠状动物亲缘的活体如蚯蚓般(图10a—h)主动卷曲形成的保存状态(图10i)。

图10 现生蠕形的蚯蚓卷曲图(a—h)及弯曲的Tawuia类化石沉积埋藏前的生态复原图(i)
近年在滇东江川生物群主要产地层位邻近的磷矿区寒武系底部磷块岩层中,又发现有规则平行且细密的横、纵纹饰,在新类型的条带状宏体碳膜化石Rugosusivitta表面同时出现(Tang Feng et al., 2021);依据其扁带状折曲、横纹疏密相间和固着(较细)端出现多条平行纵纹等性状特征而被推测为扁形动物的始祖类别(唐烽等, 2020),在形态大小上基本可以与国外伊迪卡拉系—寒武系过渡地层中多见的印痕/铸型化石Harlaniella可以对比(Li Yulan et al., 2022, in review)。多种岩性层中印痕化石和碳膜化石共存有横、纵纹的镶嵌性状,表明在伊迪卡拉纪末期原口动物中两侧对称动物底部的无体腔或假体腔类型,已经有两种体制策略在不同的沉积环境中辐射发生;甚至管状有环节/体节或有体腔的蠕形动物已经出现丰富的化石记录(华洪等, 2020a)。为了增大生态空间和运动能力,“基础动物”开始首次创新爆发,个别两侧对称生命的先驱很可能已经演化出肌肉组织和神经组织(舒德干和韩健, 2020)。基础动物向原口动物演化的可能路径应该是纵向的直纹牵引肌(蛋白),通过差异牵引或螺旋滚动使躯体弯曲或扭曲前进;横向原始体节的发生更进一步让体节内体液游移从而驱动躯体蠕动成为可能,新报道的Rugosusivitta呈现横纹间距大至小的变化特征(Li Yulan et al., 2022, in review),这样蠕动应该是能量损耗较低的方式,也促使真体腔开始出现,可以有效隔离消化和循环器官的功能。
江川生物群中Tawuia类化石迄今所发现的弯曲变形类型最多(表1),对弯曲变形的机制解释更倾向于动物属性的早期生命自主卷曲后保存,而非水流冲刷弯曲后埋藏;在弯曲处碳膜横节纹保存显示可能有体壁环节的存在,新出露标本尤其在周缘普遍保存较厚的碳膜也表明这类变形化石的体壁可能是多层或很厚的有机质致密层,富含柔韧易于变形的肌肉组织,与早期多细胞藻类外壁组成应该是差别很大,抵抗沉积风化的能力更强。但是大约同时代国内外均已发生多层共管或套管的管状有节动物如Cloudina、Conotubus和Sinotubulites及相当形态化石类型的繁盛辐射(华洪等, 2020a, 2020b),相比于新元古代早期地层中即已出现并传承至伊迪卡拉纪末期的Tawuia弯曲变形类别,明显是蠕曲效率更高的进化类型。但是以上思考推测还需要更加充分的化石证据。
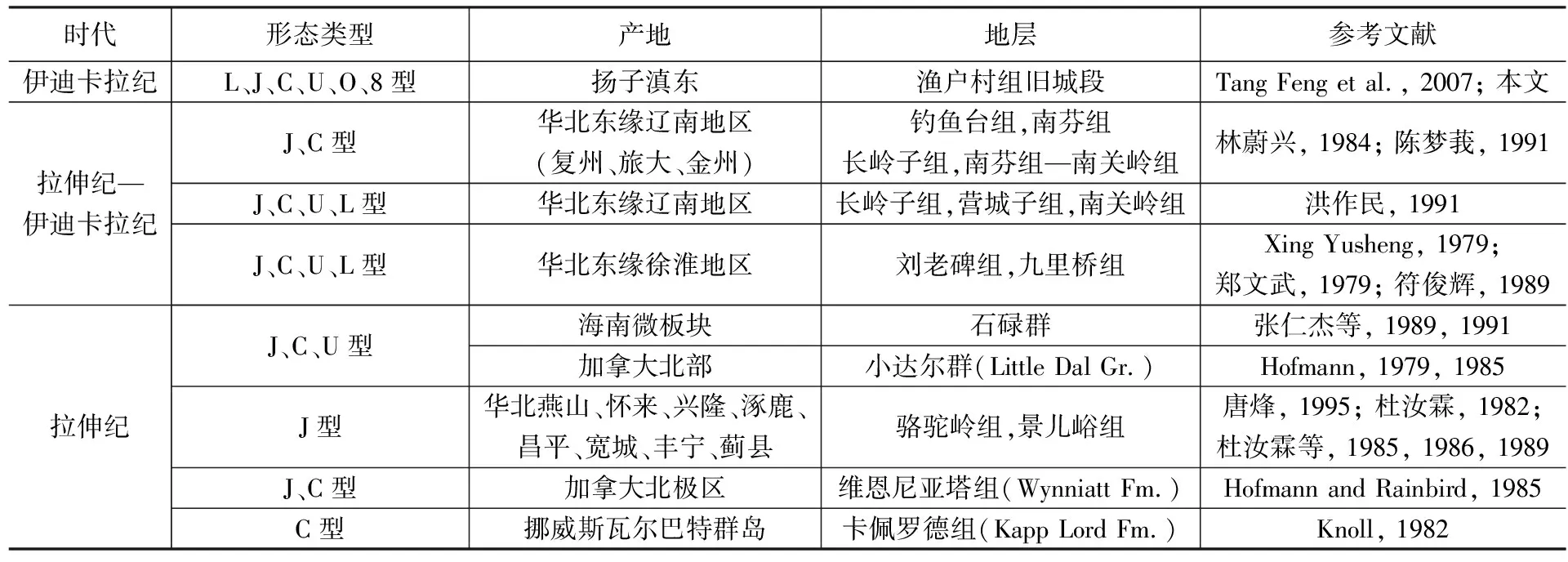
表1 弯曲变形的Tawuia类化石报道统计
3.2 Tawuia化石“盘口”形构造的地化分析与动物属性猜测
本文记述的香肠状Tawuia化石端部发现有椭圆形的缺口,暴露出碳膜缺失后的下部围岩(图2b);另外还发现有个别标本,膜面上保存功能不明的偏于一侧端部的环状碳质构造,而碳环中央碳膜较淡(图3g)。苏皖北部淮南生物群中香肠状的宏体化石也存在类似的构造(Dong Lin et al., 2008, 图8-1, 3; 钱迈平等, 2009, 图6, 11)。而与同物异名的小双盘体(郑文武等, 1994, 图1-4),乃至最新发现的华北东缘Tonian纪石旺庄组Tawuia端部明显的铁矿化似“盘口”构造均可能是同源的原生构造(Tang Qing et al., 2021a, 图6c—f)。运用新技术手段对该构造及其他化石区域进行了背散射电子扫描、能谱分析获得的图件(Tang Qing et al., 2021a, 图7),原作者观察推测为脱落构造(abscission structures),换言之,这样的圆盘构造具有吸附和脱落的功能,但经过我们的分析解读,得出了可能不同的推断:
(1)该圆盘区由颗粒状物质堆积而成,由贫铝、硅和富铁、氧的颗粒物质组成,碳、硫含量亦低,可得出该区域有机物及亲生物元素硫明显流失或者缺失,与化石体其余部位碳质含量高更有显著差别,而高于围岩的铁氧富集特征更可能表明这些颗粒为次生的磁铁矿、赤铁矿或褐铁矿一类物质。
(2)该圆盘区域显然没有附着黏土(因铝、硅含量很低),但它从环境中选择性吸附了铁离子。不论该类生物体是悬浮生活在水体中还是栖息于泥质基底上,该生物死后埋藏都保存在泥质页岩的浅海带,但却使圆盘与化石本体有显著差别(即圆盘不粘黏土),应该是死后该圆盘部位仍具有不被黏土污染的能力。
对上述两点差异,如是宏体藻类的脱落盘,则很难解释,真正的藻类接合/脱落面应该如前文图示(图4h—i)的藕节状接合的保存方式,碳质压膜的藻类化石仅可见较为平坦的端面,而不是较纯净的黄铁矿化圆盘状斑痕。对此用动物原口来推测则很好理解:因为早期动物死亡被压埋后,身体不同的组织器官均缓慢降解,但腐解速度不一样,往往动物表皮长期与环境直接接触,因适应当时环境而腐解较慢,但象内部的软体组织会在生命停止后优先降解腐败;腐败产生大量小分子物质,如蛋白质变为多肽再变为氨基酸,脂肪会降解成脂肪酸和醇类,多糖类会分解成小分子醇类等,最后释出二氧化碳、氨气、甲烷等,这些腐解后的小分子气体极易通过动物腔肠的腐烂通道被压移,而通过圆盘状原口或者后口(“盘口”)溢出,冲出的气体会将盘口附近的砂泥黏土吹走,即“吹除其附近的细小脏物”,通常在端口处呈较为平整的形态,不会形成不规则状或放射状。紧随后面的中分子醇类及脂肪酸类物质被压出,在盘口处形成一层黏滞的胶体,该胶体常溶掉氢为阴离子胶束,一般呈负电性(赵通林, 2018),进而吸附周围的铁离子、钙离子等正电性物质(硅酸盐及铝硅酸盐因表面带负电性(卢寿慈, 1988)而不会被吸附),最终使盘口出现了铝硅低而铁高的现象;后来在缓慢的化石化过程中,其身体其他部位的有机残留使其具有一定的高于环境的碳,并且铝硅等来自环境的元素也被吸附进去,导致与盘口处铁高、铝硅低的现象完全不同。后期的氧化中铁可能与氧结合造成出露的化石盘口位置铁氧富集呈现黄褐色的现象,而不是黄铁矿充填的现象。本文图示(图3g)的端部碳环构造也是在后期缓慢进行的干酪根(kerogen)化作用会使有机内含物逐渐富集,围绕盘口呈较为规则的环形碳质加厚沉积而保存下来的。另外从Tang Qing(2021a, b)、Li Guangjin(2020)等文献的化石图版中还可见到盘口中心很黑的碳质残留及轴部碳黑条带,特别是化石体中不规则分布的碳核体,我们的推测更可能是有些难于溶解的或被外层胶体包裹保护的大分子量有机物如干酪根、沥青质一直残留,并在生物尸体中运移富集保存的结果(周林飞等, 2016; 焦存礼, 2021, 私人通信)。
Tang Qing等(2021a, b)也认为这些Tawuia类的直棒形标本端部的黄色圆盘应是生物本身的构造,故其死亡后沉积下来的圆盘为规则的;但却认为黄色斑痕是大量的黄铁矿充填的现象,以及这些标本是多核体的真核藻类或外共生的真核生物共同保存的化石体。而一般情况下,若是体表较为均一的生物有机体,比如叶状体藻类,被埋藏时逐渐压实、放气、收缩等的化石化过程中,才会随机在边缘寻找压扁的薄弱口形成一些平整的不规则裂纹或放射性的构造。迄今已知的藻类生物从没有发育端部的功能不明的圆盘状原生构造。
不过这些形态特征简单的宏体碳膜化石表面构造的成因显然也存在多解性,如果要确定出最合理的生物属性解释,还需要更多的证据加持。
4 结论
国内、外比较公认的Tawuia化石出现于新元古代早期,基本形态为直棒I形和香肠形,若干化石富集的产地和层位中还伴生有弯曲马蹄C形和U形,直至新元古代全球大冰期前,保存形态均比较稳定。而冰期之后华南滇东新元古代末期江川生物群产出的Tawuia类化石则形态明显多样化,除了直棒形、哑铃形和鞋履形以外,比较多的出现了披针形和梭形(纺锤形)的类别,以及弯曲变形的L、C和U形,特别发现有环状的O形和反向弯转的8字形。依据这些弯曲化石赋存的沉积环境和保存状态,以及近期在华北东缘淮南和鲁西地区报道的完美化石记录,本文推断Tawuia大类亲缘属性复杂,部分直棒形、香肠形、披针形和梭形、纺锤形化石归属多核体或多细胞藻类基本无疑;而其他规则变形保存为C、L、J、U、O、S、8字形的化石很难解释为死亡藻类沉积埋藏时被动变形所致,更可能是两侧对称动物亲缘的近海底栖生物的主动弯曲状态,经特异埋藏而保存;部分保存有特殊盘口状构造的标本,进一步解读其地化分析结果也倾向于软躯体动物腐解风化缓慢压埋而成。
致谢:谨以此文祝贺杨文采主编80华诞!贵州省地质调查院副总工程师陈建书研究员、贵州大学王约教授多次参与野外工作,中国地质科学院硕士研究生顾鹏、云南大学博士研究生张光旭参与野外化石采集和剖面测制,耶鲁大学的Lidya TARHAN助理教授对本文的英文摘要进行了修改。
猜你喜欢
今日畜牧兽医(2022年10期)2022-12-23
奥秘(2022年7期)2022-08-09
奥秘(2022年4期)2022-04-26
当代水产(2021年8期)2021-11-04
潍坊学院学报(2020年2期)2021-01-18
今日农业(2020年15期)2020-12-15
科学(2020年4期)2020-11-26
军事文摘(2020年20期)2020-11-16
科学(2019年3期)2019-09-10
好日子(2019年5期)2019-07-19
