
玉米糖肽的体外抗氧化活性
2022-09-28 08:10王晓杰刘晓兰
中国粮油学报 2022年8期
王晓杰, 童 玲, 刘晓兰
(齐齐哈尔大学食品与生物工程学院;黑龙江省玉米深加工理论与技术重点实验室,齐齐哈尔 161006)
转谷氨酰胺酶(Transglutaminase,TGase),全称为蛋白质-谷氨酰胺-γ-谷氨酰转移酶,属于转移酶类(EC 2.3.2.13)[1]。根据酰基受体的不同,TGase可以催化交联、酰基转移 (即酶法糖基化)和脱酰胺3种类型反应,这3种反应均可以用于改善蛋白质的功能性质[2-4]。相对于TGase催化的蛋白质交联反应的研究,TGase途径的蛋白质酶法糖基化反应的研究起步较晚。与美拉德反应相比,TGase途径的酶法糖基化反应条件更温和,产物更安全。因此,TGase催化的酶法糖基化反应已经应用于多种蛋白质/多肽的改性中,包括酪蛋白、大豆蛋白、鸡肌动球蛋白、虾原肌球蛋白、黑豆蛋白、乳清蛋白、豌豆蛋白、玉米醇溶蛋白和谷蛋白、鱼皮胶原蛋白肽、小麦谷蛋白肽等[5-10]。在TGase催化的酶法糖基化反应中,蛋白质/多肽功能性质的改善程度与底物蛋白及氨基糖的性质等因素有关。例如,当以D-氨基葡萄糖为酰基供体,修饰鸡肌动球蛋白、鸡胸肉肌原纤维蛋白、大豆蛋白及酪蛋白时,由于D-氨基葡萄糖附着所赋予的亲水性,改善了原蛋白的溶解性、乳化性和凝胶特性[11-13];而修饰乳清分离蛋白时,D-氨基葡萄糖的共价结合使乳清分离蛋白的乳化性质降低[8];修饰酪蛋白酸钠水解产物时,在TGase处理前后产物的抗氧化活性之间无显著性变化[14]。
玉米肽是以玉米蛋白为原料,经蛋白酶水解或微生物发酵后获得的小分子质量多肽。玉米肽特殊的氨基酸组成与排列顺序决定了其具有多重的生物活性,特别是在抗氧化活性方面研究报道较多。相对于已应用的抗氧化剂如GSH、维生素C来说,玉米肽的抗氧化活性较低,不利于功能性食品的开发。但是,玉米肽属于天然、安全的抗氧化剂,如果能进一步改善其抗氧化活性,将促进玉米加工企业及功能食品等相关产业发展。玉米糖肽是玉米蛋白先经蛋白酶水解获得的低分子质量玉米肽,再在TGase催化下与氨基糖共价结合的产物。与玉米肽相比,玉米糖肽的溶解性显著增加,但其生物活性的改善程度鲜有研究。因此,本实验以D-氨基葡萄糖共价修饰玉米肽的产物-玉米糖肽为原料,利用体外化学法和H2O2诱导的Caco-2细胞的氧化性损伤模型对玉米糖肽的抗氧化活性进行了研究,为抗氧化活性玉米糖肽作为功能性食品应用于食品工业提供参考。
1 材料与方法
1.1 材料与试剂
碱性蛋白酶Alcalase,酶活力6.28×105U/mL;Caco-2细胞;玉米醇溶蛋白、1,1-二苯基-2-三硝基苯肼(DPPH)、2-脱氧-D-核糖,均为色谱纯;噻唑蓝(MTT)、D-氨基葡萄糖盐酸盐,均为分析纯;TGase,酶活力1 000 U/g;DMEM(高糖)培养基;胎牛血清;超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、谷胱甘肽还原酶(GR)、γ-谷氨酰半胱氨酸合成酶(γ-GCS)测定试剂盒。
1.2 仪器与设备
NDA701杜马斯定氮仪,TU1901紫外可见分光光度计,IM-45L-1-25制冰机,LD-53真空冷冻干燥机,NV 4750E二氧化碳培养箱,MB100-2A微孔板恒温振荡器,EnSpire多功能酶标仪,SX-500高压蒸汽灭菌锅。
1.3 方法
1.3.1 玉米肽的制备
40 g玉米醇溶蛋白用蒸馏水配成底物质量浓度0.05 g/mL的悬浮液,在pH 8.5、温度60 ℃、Alcalase酶与玉米醇溶蛋白的质量比3%、时间2.0 h条件下酶解2.0 h。酶解结束后,收集上清液经冷冻干燥后获得玉米肽[15]。
1.3.2 玉米糖肽的制备
将玉米肽配成质量浓度为0.03 g/mL的溶液,按玉米肽与D-氨基葡萄糖的质量比1∶3加入D-氨基葡萄糖,用2 mol/L NaOH调节pH至7.7,按加酶量55 U/g蛋白加入TGase,在44 ℃恒温水浴振荡器中糖基化反应7 h。7 h后,将反应物立即放入85 ℃水浴中灭酶5 min。冷却至室温,4 000 r/min离心10 min,收集上清液过截断分子质量300 u的纳滤膜,透过液经冷冻干燥后获得玉米糖肽混合物[16]。
1.3.3 玉米糖肽的体外消化
将底物质量浓度为0.1 g/mL 的玉米糖肽和玉米肽溶液分别用浓盐酸调节pH至2.0,按酶与底物的质量比为2%加入胃蛋白酶,37 ℃条件下水解90 min。用6 mol/L NaOH调节pH至7.5,按酶与底物的质量比为4%加入胰蛋白酶,37 ℃条件下水解240 min。酶解产物经沸水浴灭酶10 min,冷却至室温后冷冻干燥,获得玉米糖肽和玉米肽消化产物。
1.3.4 利用体外化学法研究玉米糖肽的抗氧化活性
用3种自由基(包括DPPH、羟基和超氧阴离子)清除能力、Fe2+螯合能力和还原力表征玉米糖肽的抗氧化活性,参照Wang等[17]描述的方法进行测定。
1.3.5 体外消化对玉米糖肽分子质量分布的影响
采用配有凝胶层析色谱柱Superdex Peptide 10/300 GL的蛋白质纯化系统测定。色谱柱先用蓝色葡聚糖2000测定外水体积V0,用标准蛋白(aprotinine, 6 500 u;杆菌肽, 1 400 u;氧化型谷胱甘肽, 612 u;还原型谷胱甘肽,307 u)进行校正。将样品配成蛋白质量浓度为2 mg/mL的溶液,经离心和0.22 μm微孔滤膜过滤后加载于色谱柱。上样体积100 μL,流速0.25 mL/min,检测波长214 nm。溶解样品及色谱柱的平衡和洗脱均采用pH 7.0、20 mmol/L的磷酸盐缓冲液(含有0.15 mol/L NaCl)。基于样品组分的洗脱体积(Ve)计算有效分配系数及样品组分的分子质量。
1.3.6 利用H2O2诱导的Caco-2细胞氧化性损伤模型研究玉米糖肽的抗氧化活性
1.3.6.1 玉米糖肽对Caco-2细胞毒性的测定
采用细胞存活率表征玉米糖肽是否对Caco-2细胞产生毒性效应。取96孔板,每孔加入100 μL、细胞密度为1×105个细胞/mL(以下实验均采用该密度)、对数生长期的Caco-2细胞悬液,在37 ℃的二氧化碳培养箱中培养24 h。实验分为样品组和对照组,样品组中每孔加入100 μL玉米糖肽溶液,使终浓度分别为5、25、50、100、200、500、1 000 μg/mL,对照组每孔加入100 μL DMEM培养基,37 ℃培养6 h。培养结束后,吸弃孔内液体,细胞用磷酸盐缓冲液(PBS,0.01 mol/L、pH 7.2~7.4,下同)洗2次,然后每孔加入100 μL PBS缓冲液和终质量浓度为0.5 mg/mL的MTT溶液,37 ℃培养4 h后用酶标仪检测570 nm波长下的吸光值,并计算Caco-2细胞的存活率。

1.3.6.2 H2O2氧化应激条件下玉米糖肽对Caco-2细胞存活率的影响
将100 μL对数期Caco-2细胞悬液接种至96孔板,培养24 h后,每孔加入100 μL终质量浓度分别为5、50、100、200、500、1 000 μg/mL (同时设置对照组和H2O2组)的玉米糖肽溶液,作用6 h后吸弃孔内液体,加入200 μL 200 μmol/L H2O2溶液培养4 h进行造模,根据1.3.6.1中 MTT法测定细胞存活率。
1.3.6.3 玉米糖肽对H2O2诱导的氧化性损伤Caco-2细胞内抗氧化酶活力的影响
按1.3.6.2的方法细胞培养和造模,玉米糖肽的终质量浓度分别为5、25、50、75 μg/mL。培养结束后,加入适量胰蛋白酶消化细胞,并收集细胞沉淀。用PBS缓冲液洗涤细胞1次后加入高效RIPA裂解液,4 ℃裂解20~30 min,10 000 r/min 离心5 min后,收集上清液于-80 ℃保存备用。取适量裂解液放置于冰水浴中,按照试剂盒说明书测定SOD、CAT、GR和γ-GCS活力,酶活力单位以U/mg表示。
1.3.7 数据处理
应用SPSS Statistics 19.0统计软件进行数据处理,所有数据均以“平均值±标准差”表示,多样本均数间比较采用One-way ANOVA检验,各组间多重比较采用LSD法,P<0.05认为具有显著性差异。
2 结果与讨论
2.1 糖基化修饰对玉米肽及体外消化对玉米糖肽抗氧化活性的影响
2.1.1 DPPH自由基清除能力
以GSH为阳性对照,测定糖基化修饰对玉米肽以及体外消化对玉米糖肽DPPH自由基清除活性的影响,结果如图1所示。
TGase催化的糖基化反应使玉米肽对DPPH自由基的清除能力显著增加(P<0.05);经胃蛋白酶和胰蛋白酶两步消化后,玉米糖肽的DPPH自由基清除能力显著增加(P<0.05),而玉米肽质量浓度为1.5、2.0 mg/mL时DPPH自由基清除能力也显著增加。在质量浓度为2 mg/mL时,玉米糖肽消化产物的DPPH自由基清除率为78.27%,比消化前提高19.07%,比玉米肽高30.8%,与0.5 mg/mL GSH的清除能力相当。相对于玉米肽而言,玉米肽与D-氨基葡萄糖发生糖基化反应后,D-氨基葡萄糖分子上具有还原性的半缩醛羟基以及碳氢链上的羟基结构能够提供更多H+将DPPH自由基还原成稳定的DPPH-H,从而糖基化产物具有更强的DPPH自由基清除能力。同理,体外消化过程可以将更多氢或电子供体暴露出来,可以更有效的捕获DPPH自由基,导致消化产物具有更强的DPPH自由基清除能力。García-Mora等[18]研究了体外消化对小扁豆肽抗氧化活性稳定性的影响,发现消化酶的作用会增加小扁豆肽的抗氧化活性,这是因为小扁豆肽经消化释放出的较小的肽片段和氨基酸会对抗氧化活性产生加性和协同的生物学效应,与本研究的结果一致。

注:相同质量浓度不同小写字母表示差异显著(P<0.05),下同。图1 糖基化修饰对玉米肽及体外消化对玉米糖肽DPPH自由基清除能力的影响
2.1.2 羟基自由基清除能力
以GSH为阳性对照,测定样品对羟基自由基的清除活性,结果如图2所示。
糖基化修饰使玉米肽对羟基自由基的清除能力显著增加,尤其是在1.5 mg/mL时,其清除能力与同浓度GSH相当。经胃蛋白酶和胰蛋白酶消化后,除0.5 mg/mL外,玉米糖肽的羟基自由基清除能力虽然增加,但与消化前相比无显著性差异(P>0.05)。在质量浓度为1~2 mg/mL范围内,与玉米肽相比,玉米糖肽的羟基自由基清除能力增强,分析有两方面原因:玉米糖肽作为氢供体与羟基自由基反应,将其还原成稳定的结构而被猝灭;玉米糖肽作为金属离子螯合剂与过渡态金属离子如Fe2+发生螯合作用,破坏Fenton 反应或者Haber-Weiss反应,抑制了羟基自由基的产生,进而减少了戊糖氧化产物的生成。
体外消化不能破坏玉米糖肽对羟基自由基的清除能力,这可能归因于3个方面:与玉米糖肽的氨基酸组成有关。Ren等[19]发现含有Pro、Glu和Asp的玉米蛋白水解物对消化酶具有很强的抵抗力,而玉米糖肽富含Pro、Glu和Asp[20];与玉米糖肽的分子结构有关。玉米糖肽的分子质量小,空间构象简单,又由于胃蛋白酶和胰蛋白酶的位点特异性,两步消化对玉米糖肽的组成和构象影响较小,进而没有对玉米糖肽的羟基自由基清除能力产生显著性影响;与制备玉米糖肽的蛋白酶种类有关。Xie等[21]研究了体外消化过程中酪蛋白抗氧化肽的稳定性,发现碱性蛋白酶Alcalase 可以促进抗胃肠消化肽的产生。
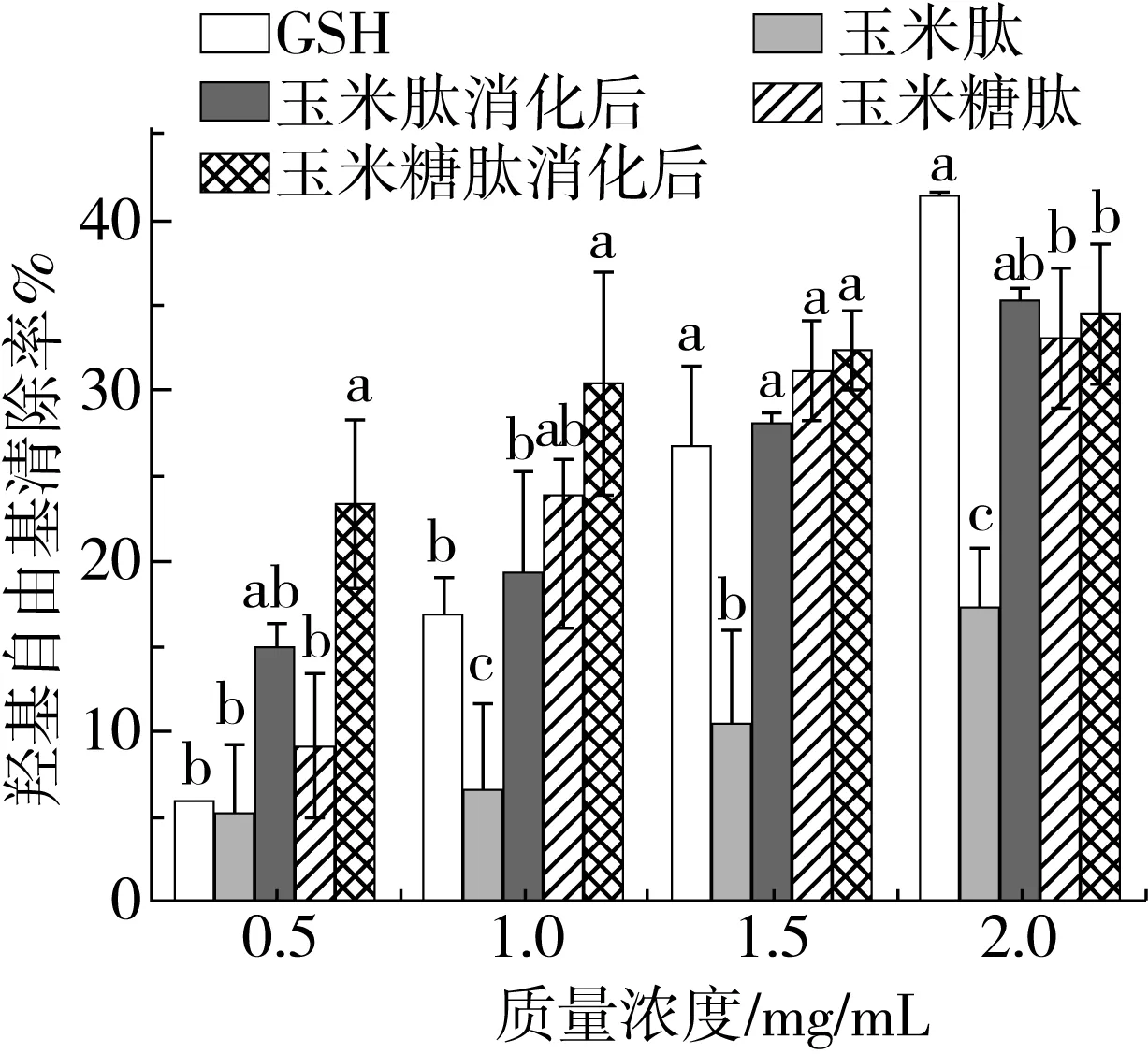
图2 糖基化修饰对玉米肽及体外消化对玉米糖肽羟基自由基清除能力的影响
2.1.3 超氧阴离子自由基清除活性
以GSH为阳性对照,测定样品对超氧阴离子自由基的清除活性,结果如图3所示。D-氨基葡萄糖的共价修饰使玉米肽的超氧阴离子自由基清除能力显著增加(P<0.05),且胃蛋白酶和胰蛋白酶的消化作用不能破坏玉米糖肽的超氧阴离子自由基清除能力。在质量浓度为2 mg/mL时,玉米糖肽消化产物的清除率为32.46%,比消化前高3.63%;玉米肽消化产物的清除率为21.93%,比消化前高2.44%;玉米糖肽的清除率为28.82%,比玉米肽高9.33%,可能是因为D-氨基葡萄糖的多羟基结构和玉米肽分子中的抗氧化活性氨基酸残基使玉米糖肽成为良好的供氢体,猝灭了超氧阴离子自由基。

图3 糖基化修饰对玉米肽及体外消化对玉米糖肽超氧阴离子自由基清除能力的影响
2.1.4 还原力
还原力也是待测样品抗氧化能力强度的测定,结果如图4所示。TGase催化的糖基化反应使玉米肽的还原能力显著增加。在质量浓度为0.5 mg/mL时,玉米糖肽的还原力为0.909,比玉米肽高0.765,是同浓度GSH的3.56倍,说明D-氨基葡萄糖的共价结合使玉米肽成为良好的电子供体,具有更强的还原力。经胃蛋白酶和胰蛋白酶消化后,玉米糖肽和玉米肽的还原力与消化前相比均增加,可能归因于2个方面:随着水解程度的增加,更多的极性或带电荷氨基酸侧链基团暴露出来;在消化过程中,肽键断裂提供了质子和电子的额外来源,以维持高氧化还原电位。Zhu等[22]研究了体外消化对玉米醇溶蛋白源玉米肽还原力的影响,也发现经胃蛋白酶1 h和胰酶2 h处理后消化产物的还原能力增加。
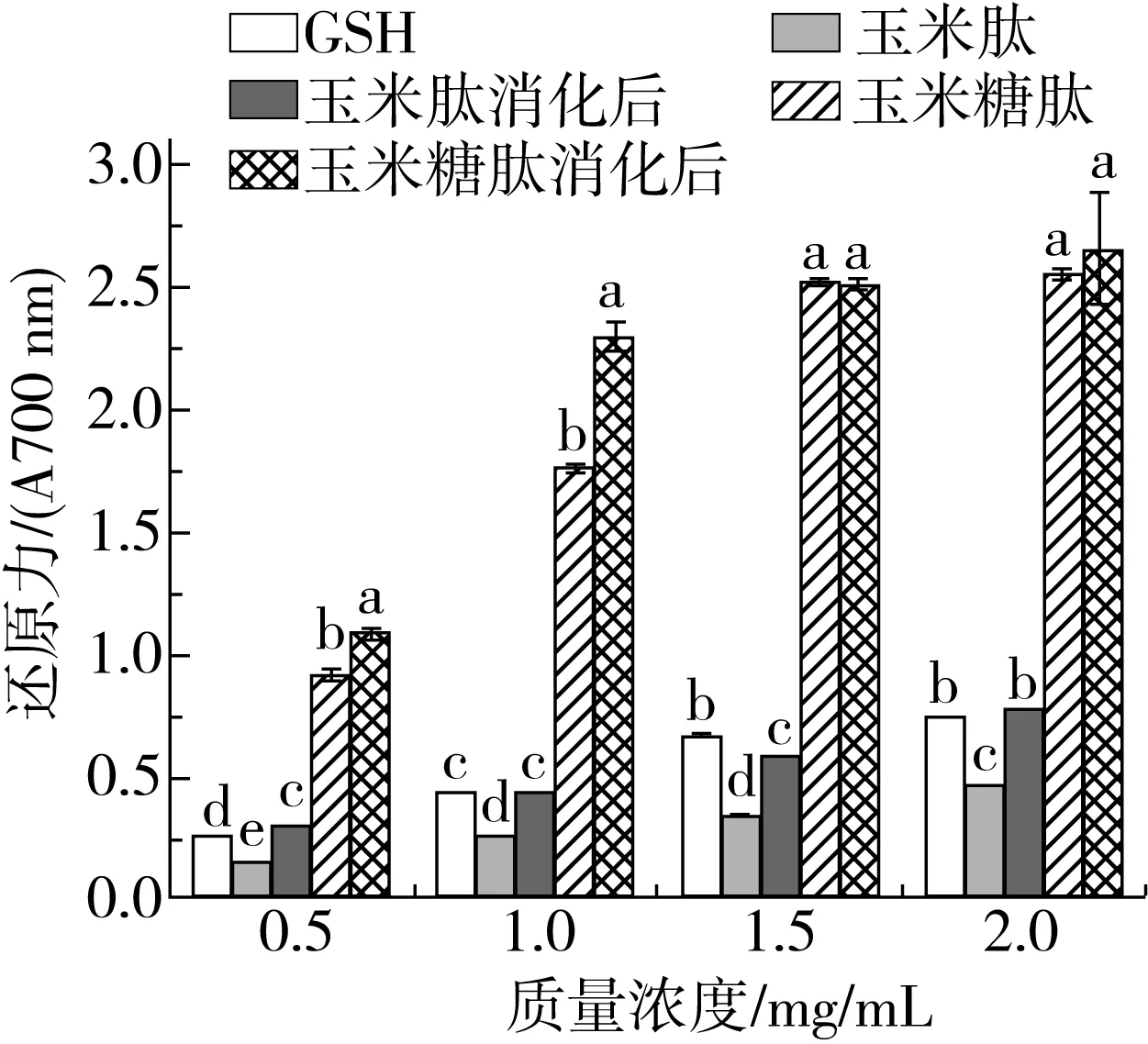
图4 糖基化修饰对玉米肽及体外消化对玉米糖肽还原力的影响
2.1.5 Fe2+螯合能力
Fe2+是过渡态金属离子中最强大的助氧化剂,其具有通过Fenton反应或Haber-Weiss反应产生羟基自由基的能力[23]。因此,具有Fe2+螯合能力就能抑制羟基自由基的产生,进而延迟氧化反应,待测样品Fe2+螯合能力的测定结果如图5所示。
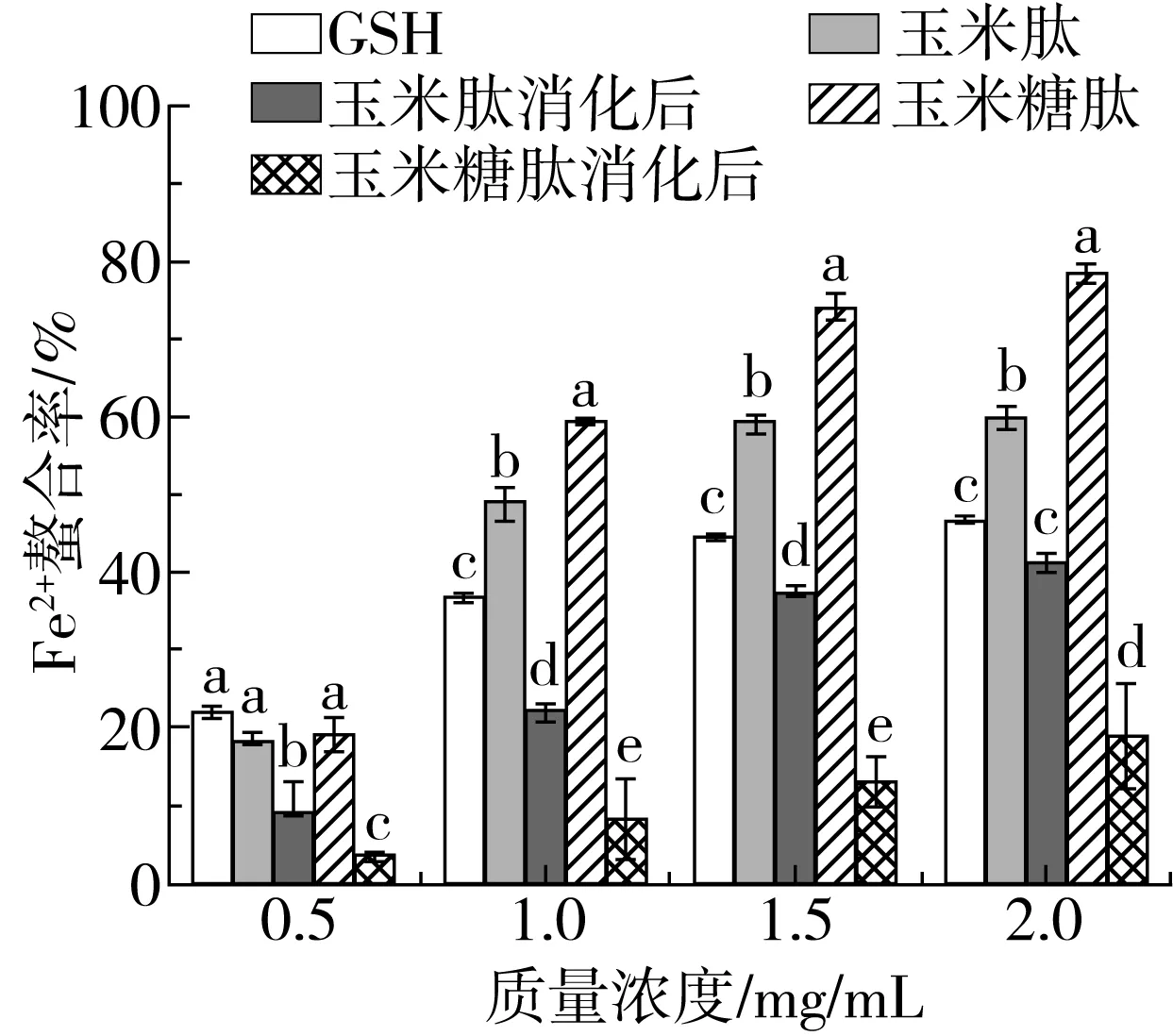
图5 糖基化对玉米肽和体外消化对玉米糖肽Fe2+螯合能力的影响
与玉米肽相比,糖基化修饰使玉米肽的Fe2+螯合能力显著增加(P<0.05)。在质量浓度为2 mg/mL时,玉米糖肽的Fe2+螯合率比同浓度玉米肽高18.67%,是同浓度GSH的1.68倍。Saiga等[24]发现酸性氨基酸残基(Asp和Glu)和碱性氨基酸残基(His、Lys和Arg)由于自身的电荷特性,侧链中的羧基和氨基在肽的Fe2+螯合能力中起着至关重要的作用。Yang等[25]也发现包含Glu、Gln和Lys的肽比不包含这些氨基酸的肽表现出更强的Fe2+螯合能力。玉米肽和玉米糖肽均富含Asp、Glu、His和Arg等残基[20],且玉米肽分子上共价结合的D-氨基葡萄糖的多羟基结构也有助于螯合Fe2+[26],因此,玉米糖肽和玉米肽均具有Fe2+螯合能力,且玉米糖肽的螯合效果高于玉米肽。
经胃蛋白酶和胰蛋白酶消化后,玉米糖肽和玉米肽的Fe2+螯合率均显著降低。孙宏[27]研究了体外消化对棉籽肽的Fe2+螯合能力的影响,发现经胃蛋白酶消化后棉籽肽的Fe2+螯合能力显著降低(P<0.05)。肽的金属离子螯合能力可以通过与带电荷的氨基酸残基间静电相互作用完成,也可以在结构上通过对过渡态金属离子的俘获作用完成[28]。经胃蛋白酶和胰蛋白酶的消化作用导致肽捕获金属离子的结构丧失,进而Fe2+螯合活性急剧下降。
因此,玉米糖肽的抗氧化能力与其供氢及电子转移能力有关,这些活性是糖基部分和作为底物的玉米肽的协同作用。
2.2 体外消化对玉米糖肽分子质量分布的影响
将样品配制成2 mg/mL的溶液,测定玉米糖肽经胃蛋白酶消化90 min和玉米糖肽经胃蛋白酶消化90 min和胰蛋白酶消化240 min时水解产物的分子质量分布,洗脱图谱如图6所示。
由图6a可看出,玉米糖肽的分子质量分布比较宽,主要由3个分子质量组分构成,其中保留时间大于18.69 min的分子质量小于300 u的组分质量分数为4.77%;在图6b中,经胃蛋白酶消化90 min后,玉米糖肽消化产物组分的保留时间向右移动,说明与玉米糖肽相比消化产物的分子质量减小,具体表现为保留时间为16.92 min组分的含量降低,而保留时间为18.38 min的分子质量小于300 u的组分质量分数增加,达到14.29%;在图6c中,玉米糖肽的分子质量进一步减少,由图6b的3个组分变成6个组分,主要组分的分子质量集中在保留时间17.03 min(52.08%),说明胃蛋白酶和胰蛋白酶的消化作用,进一步将玉米糖肽的分子质量降解为分子质量更小的肽段,甚至是游离氨基酸。综合分析,胃蛋白酶和胰蛋白酶的消化作用使玉米糖肽的分子组成发生变化,将玉米糖肽清除自由基的抗氧化活性释放出来,同时破坏了玉米糖肽螯合Fe2+的结构。

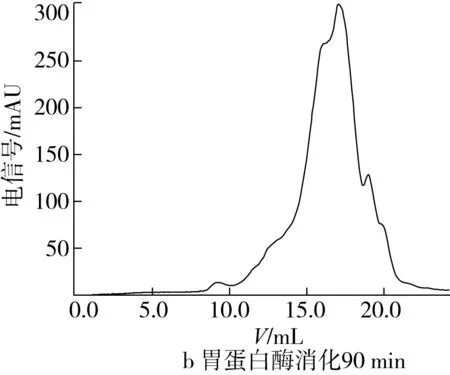
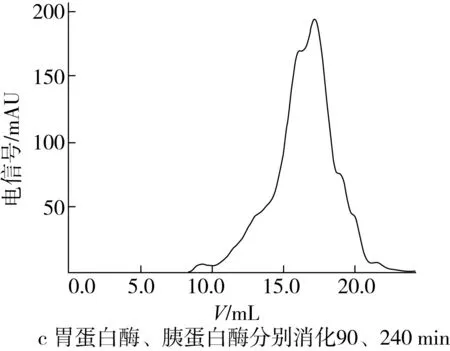
图6 玉米糖肽及其体外消化产物的凝胶层析洗脱图谱
2.3 利用H2O2诱导的Caco-2细胞氧化性损伤模型研究玉米糖肽的抗氧化活性
2.3.1 玉米糖肽对Caco-2细胞的毒性效应
细胞毒性分析是评估功能性食品生物安全性最重要的检测方法之一。不同浓度玉米糖肽作用Caco-2细胞6 h后,采用MTT法测定细胞存活率,以表征玉米糖肽的预处理是否对Caco-2细胞产生毒性效应,结果如图7所示。
在质量浓度为5~50 μg/mL范围内,随着玉米糖肽浓度的增加,Caco-2细胞存活率呈逐渐增加的变化趋势,但与对照组相比差异不显著(P>0.05);当玉米糖肽质量浓度为100 μg/mL时,Caco-2细胞存活率显著增加(P<0.05),增加了7.05%;玉米糖肽的浓度继续增加,Caco-2细胞的存活率略有下降,但与对照组相比也没有显著性差异(P>0.05),说明在实验浓度范围内,玉米糖肽对Caco-2细胞没有毒性,是安全的生物功能因子。
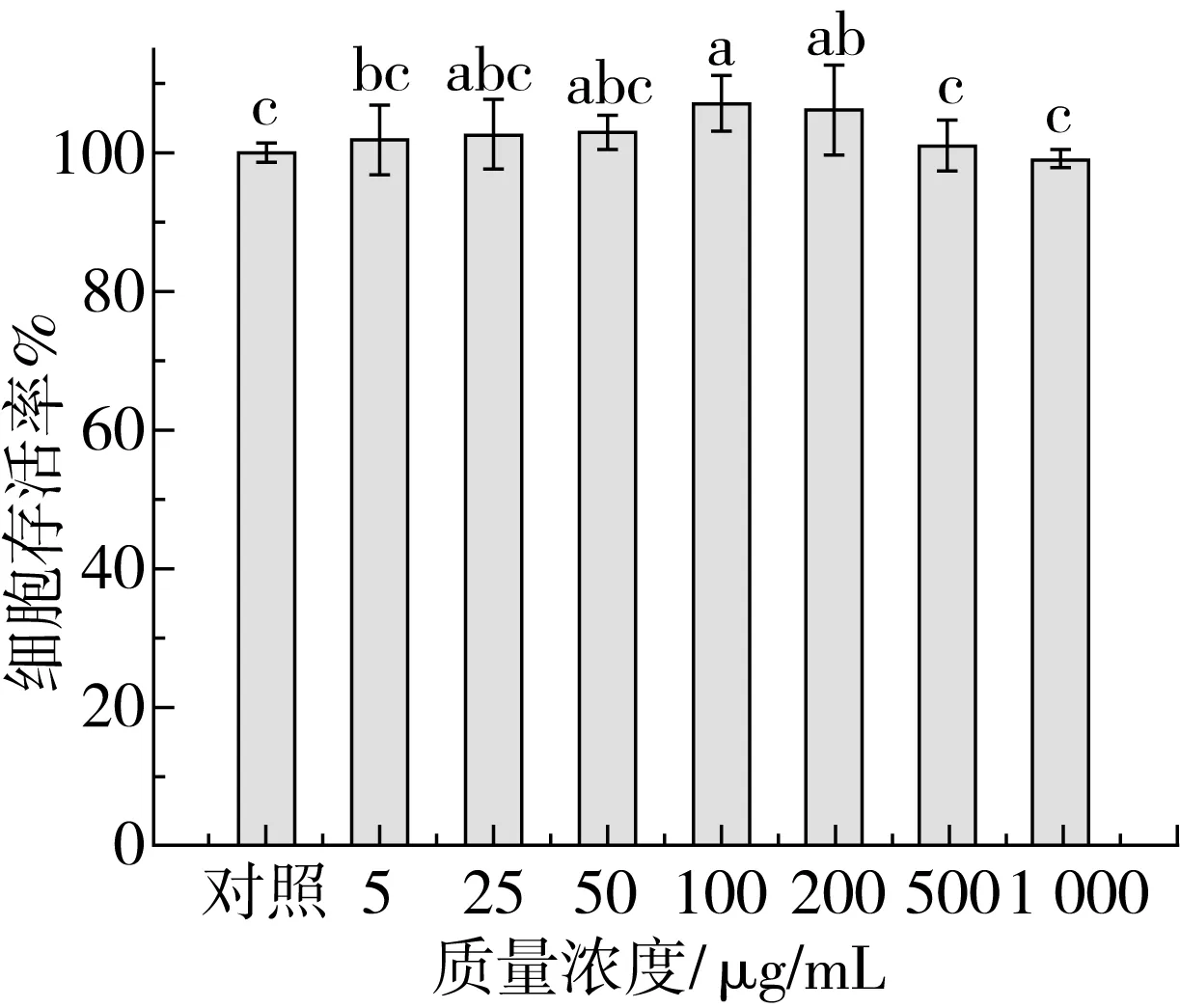
注:不同小写字母表示差异显著(P<0.05)。图7 不同浓度玉米糖肽对Caco-2细胞存活率的影响
2.3.2 玉米糖肽对H2O2诱导损伤的Caco-2细胞存活率的影响
Caco-2细胞先用玉米糖肽预处理6 h,然后暴露于200 μmol/L H2O2溶液中,研究玉米糖肽对H2O2诱导损伤的Caco-2细胞存活率的影响,结果如图8所示。与对照组相比,用200 μmol/L H2O2作用4 h后,Caco-2细胞存活率极显著降低(P<0.01),降低了40.5%,表明Caco-2氧化应激模型构建成功。玉米糖肽的预处理在一定程度上改善了受损细胞的存活率,且改善的效果呈剂量依赖性。与H2O2组相比,500 μg/mL玉米糖肽的预处理使Caco-2细胞存活率提高12.57%,1 000 μg/mL玉米糖肽的预处理使Caco-2细胞存活率提高18.65%,可能是因为玉米糖肽可以有效地进入细胞内并与氧化应激相关途径相互作用[29,30],加速清除细胞内过量的H2O2、超氧阴离子等活性氧自由基而提高了细胞存活率。
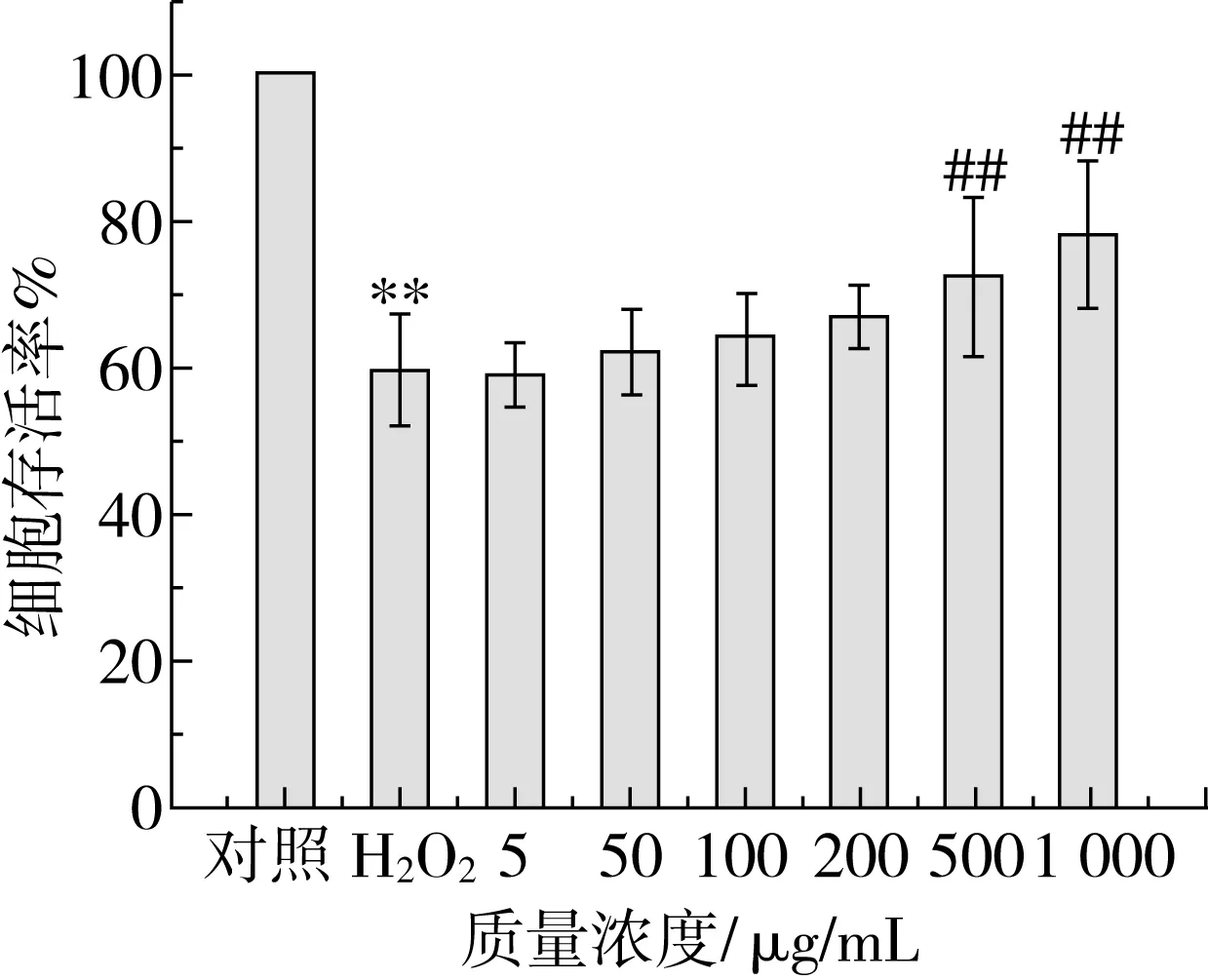
注:**表示与对照组相比差异极显著(P<0.01),##表示与H2O2组相比差异极显著(P<0.01)。图8 玉米糖肽对H2O2诱导的氧化应激条件下Caco-2细胞存活率的影响
2.3.3 玉米糖肽对H2O2诱导损伤Caco-2细胞内抗氧化酶活力的影响
为了进一步表征玉米糖肽的抗氧化能力,研究了其对氧性损伤Caco-2细胞内抗氧化酶活力的影响,结果如表1所示。与对照组相比,H2O2组细胞内CAT、SOD、γ-GCS和GR的活力分别降低了80.08%、31.69%、67.38%和70.40%,进一步说明Caco-2细胞氧化应激模型构建成功。与H2O2组相比,玉米糖肽以剂量依赖的方式拮抗了氧化应激细胞内抗氧化酶活力的降低。其中,50 μg/mL玉米糖肽的预处理可以使SOD、γ-GCS、GR和CAT活力显著提高(P<0.05),分别是H2O2组的1.09、2.75、2.54倍和3.21倍,75 μg/mL玉米糖肽的预处理甚至使4种内源抗氧化酶活力恢复到正常水平,与75 μg/mL玉米肽组的GR、SOD、CAT和γ-GCS活力相比分别高6.47%、0.88%、10.77%和0.96%[15],进一步说明D-氨基葡萄糖的糖基化修饰改善了玉米肽的抗氧化活性。这样,低剂量玉米糖肽的预处理可以通过增加Caco-2细胞内抗氧化酶的活力,加速细胞内活性氧自由基的清除,进而拮抗了H2O2诱导的Caco-2细胞的氧化性损伤。

表1 玉米糖肽对H2O2诱导损伤Caco-2细胞内 抗氧化酶活力的影响
3 结论
以还原型GSH为阳性对照,研究了糖基化修饰对玉米肽以及体外消化对玉米糖肽抗氧化活性的影响,同时构建H2O2诱导的Caco-2细胞氧化性损伤模型,在细胞水平上对玉米糖肽的抗氧化活性进行研究。D-氨基葡萄糖的糖基化修饰使玉米肽的抗氧化活性显著增加(P<0.05),且具有剂量依赖性,并且除Fe2+螯合能力外,体外模拟消化不能破坏玉米糖肽的自由基清除能力和还原力,说明玉米糖肽在胃肠道消化过程中仍能保持其抗氧化活性,能够以活性形式到达血液,进而在体内发挥其抗氧化活性。另外,质量浓度为5~1 000 μg/mL的玉米糖肽对Caco-2细胞没有毒性,质量浓度为75 μg/mL的玉米糖肽可以显著提高氧化损伤Caco-2细胞内SOD、GR、CAT和γ-GCS抗氧化酶的活力,从提高细胞抗氧化水平的角度提高了细胞存活率。在此基础上,需要对玉米糖肽的抗氧化机制进行研究。
猜你喜欢
肥料与健康(2022年3期)2022-12-04
复旦学报(医学版)(2021年5期)2021-10-13
农业与技术(2021年1期)2021-01-18
当代水产(2020年4期)2020-06-16
当代水产(2020年3期)2020-06-15
癌症进展(2018年11期)2018-12-30
科学中国人(2018年8期)2018-07-23
医学研究杂志(2015年7期)2015-06-22
医学研究杂志(2015年12期)2015-06-10
医学研究杂志(2015年12期)2015-06-10
