
低盐度养殖凡纳滨对虾对饲料甘氨酸锰为锰源的锰营养需求
2022-10-01 15:51程开敏刘艳妮刘丽燕
中国饲料 2022年16期
程开敏, 刘艳妮, 刘丽燕, 胡 蝶
(1.广东粤海饲料集团股份有限公司, 广东湛江 524017;2.广东省水产动物营养与饲料企业重点实验室, 广东湛江 524017;3.广东粤盛生物科技有限公司, 广东广州 510380)
锰是维持大脑正常生理功能的重要元素, 在机体各组织中广泛分布, 线粒体中的锰含量远比细 胞 内 其 他 细 胞 器 丰 富(Watanabe等, 1997;Lall, 1989)。锰在蛋白质、脂类与碳水化合物代谢过程中充当许多金属酶系统的重要激活因子(Davis和Gatlin, 1996), 而且也是金属酶的重要组分(Watanabe等, 1997)。由于具有重要的生理生化功能, 锰早已被认为是重要的微量元素成分之一, 这一点早已在哺乳类与鸟类的营养研究中得到印证。关于锰在鱼类营养需求研究中报道较多, 由于试验种类与大小、养殖环境及基础饲料中的锰含量不同, 鱼类对锰的需求范围为2.4~50 mg/kg(魏 万 权 等, 2001;赵 振 山 等, 1995;王道 尊 等, 1994;Lall, 1989;Gatlin和Wilson, 1984;Satoh等, 1983, 1987, 1989, 1991;Knox等, 1981;Ogino和Yang, 1980)。尽 管 鱼 类 可从水体中摄取锰用以生长需求, 但摄入锰量不足时会表现出营养缺乏症, 如生长缓慢、食欲低下、脊椎变形、机体平衡性下降、眼球白内障等, 严重时甚至影响性腺发育与繁殖机能(Watanabe等, 1997;Gatlin和Wilson, 1984)。关 于 对 虾 对 锰的营养需求也有部分报告, 诸如中国对虾(李荷芳等, 1993)、日本对虾(Kanazawa等, 1984)和凡纳滨对虾(Davis等, 1992;董晓慧等, 2005;鲁耀鹏等, 2018)等, 但从其生长表现、实验设计、养殖水体、实验结论等方面综合而言, 有关对虾对锰的营养需求有必要再行技术论证与补充完善, 尤其是过量添加会增加较大的环境负荷。近些年, 淡水养殖凡纳滨对虾发展迅猛, 据统计, 我国淡水养殖凡纳滨对虾产量比重超过养殖总量的50%(2021年中国渔业统计年鉴), 但目前缺乏低盐度养殖下的凡纳滨对虾对饲料锰营养需求的相关研究。
为提高微量矿物元素吸收利用率, 实行增效减排, 国际营养与饲料学界早已研发出有机螯合矿替代无机矿的技术做法, 并已在畜禽饲料应用上取得良好效果。本试验基于以上种种考虑, 以甘氨酸锰为锰源, 用半精制饲料研究极低盐度(2‰)养殖条件下凡纳滨对虾对饲料锰的需求量, 以期为低盐度养殖凡纳滨对虾的商业饲料开发与健康养殖提供参考。
1 材料与方法
1.1 试验饲料配方 本试验所用饲料是以酪蛋白与明胶为蛋白源的半精制饲料, 基础饲料配方见表1。该基础饲料中锰含量为2.2 mg/kg, 基础饲料中的粗蛋白质、脂肪及总能含量分别为37.9%、6.8%和17.32 kJ/kg。以甘氨酸锰(广州天科公司馈赠, 锰含量为16.5%, 氨基酸含量为45%)为锰源, 在基础饲料中分别添加0、5、10、20、30、60、90 mg/kg的锰。同时使用HP3510型火焰原子吸收分光光度计实测实验饲料锰含量, 分 别 为2.22, 7.34, 12.70, 21.35, 32.11, 59.72和90.13 mg/kg。在配制饲料过程中, 用SiO2平衡不同水平锰在饲料中的添加量。
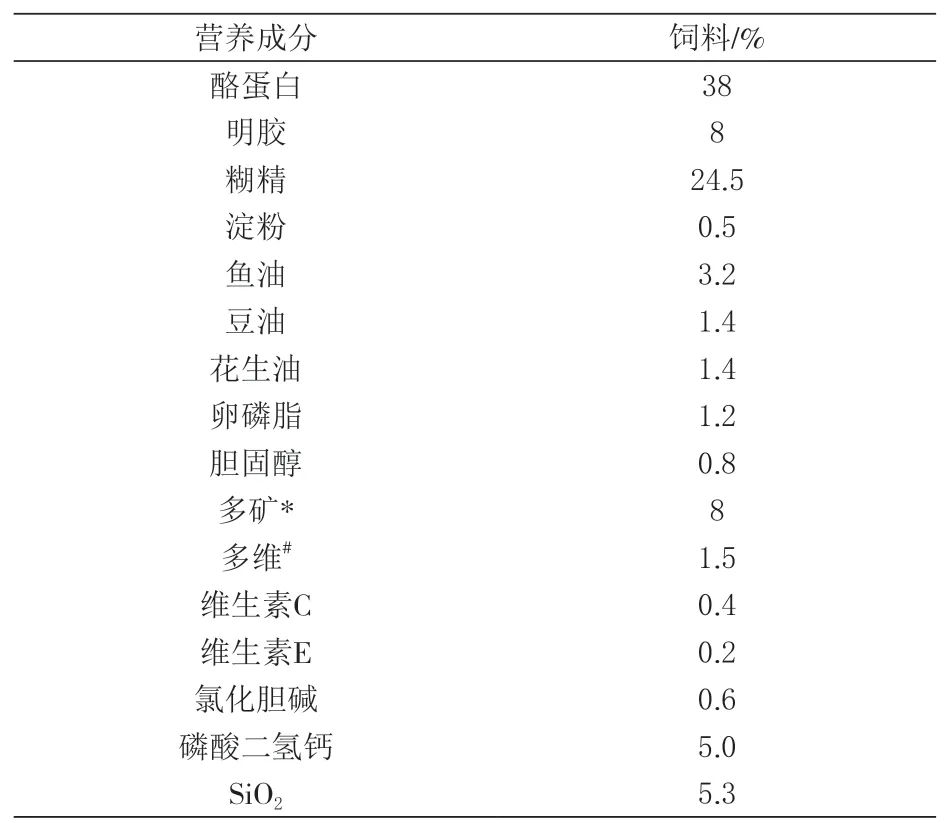
表1 试验饲料的基本配方
各饲料成分(干粉)预先在搅拌器中搅匀后缓慢加入鱼油与磷脂, 再加入适量的双蒸水进一步搅拌, 然后通过双螺杆挤调机制成粒径分别为1.0和1.2 mm的饲料, 放入空调房中风干揉搓成适宜长度, 封存放入-20℃冰箱中备用。
1.2 对虾养殖与环境监控 从粤海种苗场购买2万尾凡纳滨对虾幼苗(PL-20)(虾苗养殖环境海水盐度为28‰), 在粤海饲料对虾养殖基地的水泥池中进行低盐度的淡化驯养, 直至淡化水体的最终盐度为2‰时为止, 淡化期间水温(24±2)℃。在养殖试验开始前用基础饲料养殖2周以便让对虾适应饲料转换, 同时尽可能降低对虾体内贮存的锰含量。
试验初期每个玻璃钢桶(1个玻璃钢桶体积0.4 m3, 高0.8 m)放养30尾虾苗, 试验初期将平均体重(0.41±0.03)g的对虾分为7组, 每组3个重复。在整个试验期间, 养殖水体盐度控制在2‰, 试验全过程持续充气增氧。每天按对虾体重(湿重)的8%~10%分4次投喂(投喂时间分别是7:00、11:00、18:00和22:00), 每次投饲量分别是每天总饲量的25%、23%、32%和20%。每天记录对虾的摄食与生长动态, 并根据前1 天对虾的摄食动态适当调整每天的投饲量。每天适量换水1次, 同时排出粪便与残饵。本次养殖试验为期10周, 试验结束前1 天停止投喂饲料。
试验期间, 定期检测养殖水质状况, 水温、溶 解 氧(YSI-55溶 氧 仪)及pH, 分 别 为(28±2)℃, (8.10±5)mg/L和7.90±0.31。同时测定水质氨氮、亚硝酸盐与硝酸盐的含量, 分别 为(0.19±0.02)mg/L, (0.31±0.04)mg/L和(0.62±0.13)mg/L。本试验所用盐度为2‰的养殖用水, 预先调配并贮存在专用的蓄水池中备用。用火焰原子吸收法测定养殖试验用水的锰离子含量, 使其保持在0.01 mg/L以下。
1.3 样品采集与分析 试验结束时称重计数, 同时用1 mL的灭菌注射器从对虾的围心窦抽取血淋巴放入灭菌离心管中(每个平行分开放置), 静置于4℃条件下过夜。4℃条件下以4000 rpm的速率离心5 min, 取血清存于-80℃的超低温冰箱中。同时取肝胰腺并进行称重, 分别剥取对虾虾壳与腹部肌肉保存于-80℃用于进一步分析。
用105℃常压干燥法测定所有样品的水分, 用550℃高温炉灼烧法测定灰份含量, 用索氏抽提法测定肌肉粗脂肪含量, 用微量凯氏定氮法测定肌肉粗蛋白质。用试剂盒(购自南京建成生物试剂公司)测定血清Mn-SOD, SOD酶活定义为每毫升血清中SOD抑制率达50%时所对应的SOD量作为一个SOD活力单位(U)。所有样品的 锌(Zn)、铁(Fe)、铜(Cu)和 锰(Mn)等 微 量元素分析采用干法处理, 并将盐酸(HCl)与硝酸(HNO3)的混合液(HCl:HNO3=4:1)以1:10(m/V)的比例进行样品加热消解, 然后用HP3510型火焰原子吸收分光光度计测定。
本次试验涉及的公式如下:

1.4 数据统计与分析 本次试验采用SPSS软件程序进行统计分析。所有数据采用单因子方差分析, 采用Duncan’s法进行多重比较, P<0.05表示组间差异显著。
2 结果
2.1 对生长与成活率的影响 由表2可知, 10周试验表明, 成活率从76.67%到90.48%不等, 并没有因为处理组锰含量的不同而对成活率造成显著影响, 但生长却显著受到锰含量的影响(P<0.05), 锰含量为21.35 mg/kg时对虾增重率为986.67%, 显著高于对照组的712.47%。在增重率方面, 饲料锰含量超过60 mg/kg时会显著降低生长。
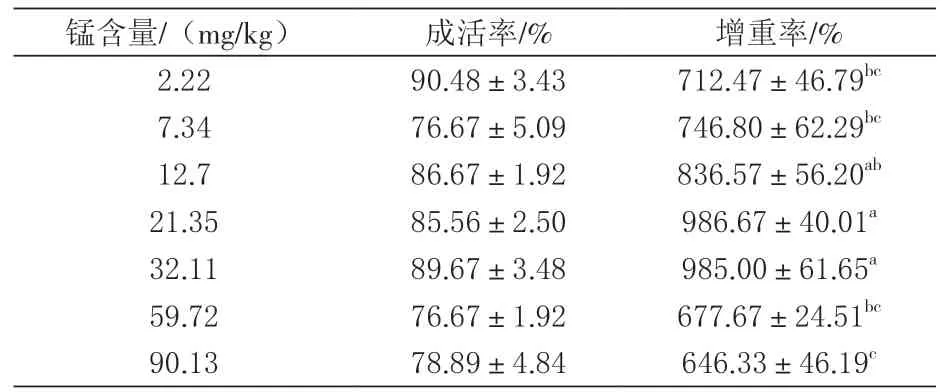
表2 2‰盐度摄食10周不同锰水平饲料的凡纳滨对虾增重率、成活率
2.2 对肝胰腺指数与血清酶活力的影响 由表3可知, 在极低盐度下, 不同锰含量对其肝胰腺指数造成显著影响(P<0.05), 饲料锰含量增加到60 mg/kg以上时, 总体表现出肝胰腺指数会显著降低。对虾血清Mn-SOD活力显著受锰含量的影响, 饲料锰含量为21.35 mg/kg时其活力最大, 随着锰含量的增加表现出下降趋势。
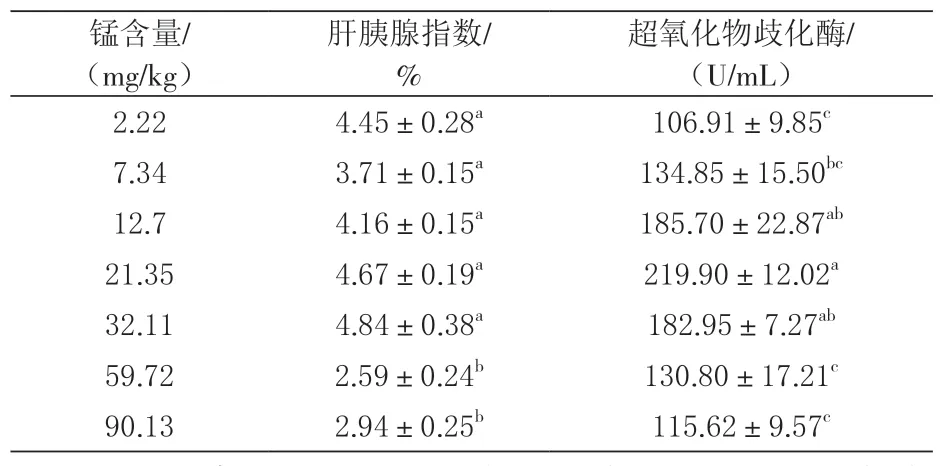
表3 不同锰水平对凡纳滨对虾肝胰腺指数及血清超氧化物歧化酶的影响
2.3 肌肉常规成分 由表4可知, 无论是肌肉水分、灰分、粗蛋白质, 还是粗脂肪含量都没有因为锰含量的不同而表现出显著性差异。
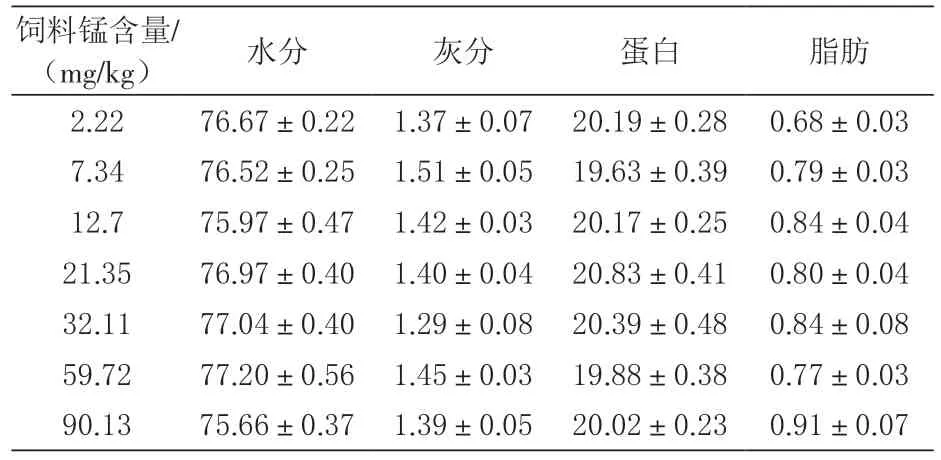
表4 摄食不同锰水平饲料的凡纳滨对虾肌肉 组成成分 %
2.4 对组织矿物元素的影响 由表5可知, 各处理组对虾肌肉中的Mn、Zn和Cu含量没有因摄食饲料锰含量的不同而有显著差异, 但对Fe含量影响显著(P<0.05)。由表6可知, 甲壳中的Fe和Cu含量没有显著差异(P>0.05);甲壳Mn含量与饲料锰含量相关, 摄食过量的锰会导致甲壳Mn含量显著升高(P<0.05);饲料锰含量显著影响甲壳中的Zn含量(P<0.05), 表现出饲料锰含量对甲壳Zn含量具有抑制作用。由表7可知, 肝胰腺中的Mn会随饲料锰含量的增加有提升趋势, 高锰可显著降低肝胰腺中的Cu含量(P<0.05), 而肝胰腺中的Fe与Zn则无显著性差异(P>0.05)。

表5 锰水平对凡纳滨对虾肌肉矿物元素含量的影响 mg/kg
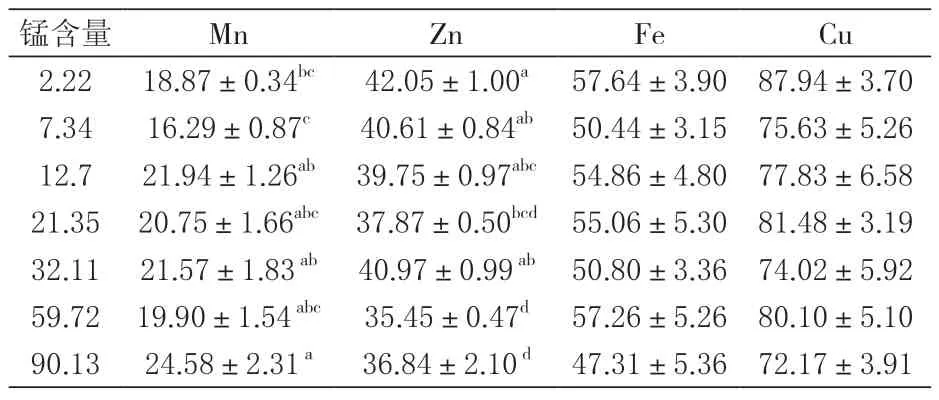
表6 锰水平对凡纳滨对虾甲壳矿物元素含量的影响 mg/kg

表7 锰含量对凡纳滨对虾肝胰腺矿物元素 含量的影响 mg/kg
3 讨论
本试验结果表明, 2‰盐度下半精制饲料中添加适量的锰有助于对虾的生长。尽管本试验中对虾的成活率没有显著差异, 但在生长性能方面表现出显著差异。与对照组相比, 21.35 mg/kg锰含量组对虾的增重率最大, 表明基础饲料中的锰含量不足以满足对虾生长需要, 必须在饲料中添加适量的锰, 而且在此水体中锰含量不足0.01 mg/L的条件下, 对虾不可能从水体中获得足够的锰来满足生长所需。在试验中, 对虾的生长未出现平台期效应, 可能是因为试验设计的锰梯度较大, 另一方面可能是对虾对锰的耐受限度较小。添加锰量>60 mg/kg则会抑制对虾的生长, 有关原因或代谢机制尚需深入, 但提示饲料锰的添加量应注意不宜过多。
本试验综合评估对虾的增重率与Mn-SOD酶的活力, 推荐对虾对锰的需求量为21.35 mg/kg(甘氨酸锰为锰源)。这与董晓慧等(2005)的研究看法有点相近, 但也有不同结论认为海水养殖的凡纳滨对虾饲料锰添加量为150-200mg/kg(鲁耀鹏等, 2018)。有研究表明, 海水养殖的日本对虾不需要在饲料中添加锰, 添加30 mg/kg会抑制其生长(Deshimaru和Yone, 1978;Kanazawa等, 1984)。而Davis等(1992)认为, 海水养殖条件下, 半精制饲料中锰添加38 mg/kg与否, 对凡纳滨对虾生长没有显著性影响, 并推测对虾可从海水中吸收锰离子。本实验与上述结论有所不同, 可能与养殖水体的盐度和离子含量差异、精制饲料的有别、甚至锰源的选择与利用等因素有一定关系。关于对虾对矿物元素的需求研究较难精准, 一个重要的原因就是, 对虾具有从养殖水体中吸收矿物元素的生物学特性, 部分矿物元素甚至可能从基础饲料和水体中获取即可满足, 尤其是以鱼粉为主的饲料中含有一定的微量元素。本实验中, 2‰极低盐度的养殖水体肯定比海水中的矿物元素含量少得多, 在很大程度上降低对虾从水体中的摄入量, 用半精制饲料研究对虾对锰的营养需求相对更为精准, 可能极低盐度的养殖水体会因为环境胁迫导致对虾对其营养需求有所变化。
SOD酶活力已作为反映营养需求、免疫增强与养殖环境胁迫监控等方面的一个指标。在真核生物中, Mn-SOD主要存在于线粒体中, 在清除超氧阴离子的氧化代谢过程中发挥了重要作用(Wang和Chen, 2005;Belinky等, 2002)。Mn-SOD酶活可以被锰离子所诱导, 这在微生物细胞中已被证实。在甲壳动物的免疫体系中, 血淋巴中的SOD酶活力具有重要调制作用(Córdova等, 2002)。在本试验中, 血清Mn-SOD酶活显著受到饲料锰含量的影响, 在一定范围内随锰含量的增加而增加, 但锰含量超过21.35 mg/kg后, 该酶活性逐步下降。这与鲁耀鹏等(2018)在凡纳滨对虾的相关研究结论类似, 同样在虹鳟(Knox等, 1981)和一些陆生动物(De Rosa 等, 1980)实验中也有类似报告。
本试验结果表明, 饲料锰含量虽然对对虾肌肉蛋白质与脂肪含量没有造成显著影响, 但会对对虾各组织中的铜、锌与铁离子的含量产生一定影响, 表现为肌肉中的铁、甲壳中的锌以及肝胰腺中的铜离子含量与饲料锰含量总体呈负相关。Mai和Tan(2000)发现, 鲍鱼(Haliotis discus hannai Ino.)在摄食硫酸亚铁与蛋氨酸铁时, 贝壳中的锰含量无显著影响, 但摄入硫酸亚铁时对其软体组织中的锰含量有抑制作用, 而蛋氨酸铁并没有对软体组织中的锰含量造成显著影响。有学者发现, 一些甲壳类, 诸如斑节对虾、墨吉对虾、加州美对虾, 其肌肉中锰含量与体重呈 负 相 关(Darmono和Denton, 1990;García和Fowler, 1972), 但凡纳滨对虾并没有表现类似趋势(Páez-Osuna和Ruiz-Fernández, 1995)。
本试验中, 锰含量在对虾肌肉中的差异不显著, 但甲壳与肝胰腺中的含量随锰含量的增加而显著上升。Davis等(1992)发现, 养殖海水的凡纳滨对虾摄食不同含量的锰, 其肝胰腺的锰含量没有显著差异, 但饲料中没有添加锰时, 甲壳中的锰含量显著下降。挪威龙虾体内锰离子的代谢动力学研究表明, 生活在不同锰离子的水体中, 其肌肉中的锰含量保持相对稳定, 但甲壳中的锰含量增加。在蜕皮后期, 甲壳组织钙化时锰离子可被吸收到碳酸钙框架中, 甚至通过甲壳表面的吸附作用直接富集水体中的锰离子, 并在体表形成黑斑;肝胰腺也具有富集锰离子的作用, 尤其是其他组织中的锰离子过剩时, 会直接转运到肝胰腺中导致锰含量的增加(Baden等, 1999。类似地, 鱼类骨骼中的锰含量与饲料锰也有相关性(魏万 权 和 李 爱 杰, 2001;Gatlin和Wilson, 1984;Satoh等, 1983;Knox等, 1981;Ogino和Yang, 1980)。
4 结论
综合对虾生长与有关生理活动等指标, 推荐极低盐度养殖条件下凡纳滨对虾对锰的需求量为20 mg/kg(甘氨酸为锰源), 添加锰具有一定的生理功能, 对不同组织微量元素含量与吸收具有一定的影响, 提示在淡化养虾与商业饲料开发中应关注锰添加的剂型与剂量。
猜你喜欢
河北渔业(2022年10期)2022-10-15
当代水产(2022年8期)2022-09-20
水产科学(2022年4期)2022-07-20
当代水产(2022年5期)2022-06-05
文萃报·周二版(2022年10期)2022-03-19
健康之家(2021年6期)2021-09-08
人人健康(2020年4期)2020-05-25
食品与生活(2018年7期)2018-09-19
漫画月刊·哈版(2017年7期)2018-01-29
恋爱婚姻家庭·养生版(2016年11期)2016-11-03
