
接种菌根真菌和氮添加处理对樟子松苗木根际微生态环境的影响
2022-10-04 03:18郝龙飞邵东华刘婷岩许吉康张之月于凡舒
西北林学院学报 2022年5期
郝龙飞,小 红,邵东华,刘婷岩,许吉康,张之月,于凡舒
(内蒙古农业大学 林学院,内蒙古 呼和浩特 010019)
土壤酶是生态系统中不可缺少的一类蛋白质化合物的总称,主要来自微生物和根系分泌以及植物残体分解等[1]。土壤水解酶在土壤碳(C)、氮(N)、磷(P)养分循环过程中被广泛研究,其中β-1,4葡萄糖苷酶(BG)、β-1,4-N-乙酰-氨基葡糖氨糖苷酶(NAG)、亮氨酸氨基肽酶(LAP)、磷酸酶(AP)在土壤C、N、P转化和迁移过程中发挥着重要作用[2-3]。土壤酶对外界环境变化过程较为敏感,进而反映土壤微生物的活性[4]。全球变化是导致环境变化的主要因素之一,其中N沉降增加导致陆地生态系统发生了一系列变化,如土壤酸化和微生物群落结构改变,导致土壤酶活性发生相应变化,从而影响生态系统的养分循环[5]。研究表明,土壤酶化学计量比可反映微生物群落的新陈代谢及养分需求与环境中养分有效性之间的生物地球化学平衡,并用来评价微生物 C、N、P 养分资源需求[6]。马伟伟等[7]对高山林地土壤酶活性的研究发现,高N质量分数抑制BG酶、NAG酶和AP酶活性。宰学明等[8]通过对菌根化滨梅(Prunusmaritima)苗的研究发现,菌根能增加根际土壤酶的活性,导致土壤有效养分生态化学计量比发生改变[9]。总结以往研究发现,关于根际土壤酶及其化学计量比对N添加和菌根效应多为单一因素的响应研究,二者交互作用的相关研究较为缺乏。
本研究以菌根依赖型树种樟子松(Pinussylvestrisvar.mongolica)苗木为材料,通过对比分析N添加对菌根苗和非菌根苗土壤胞外酶活性及其计量比、土壤理化性质以及微生物养分限制的影响。拟揭示菌根苗与非菌根苗根际土壤胞外酶活性和化学计量特征对不同N添加水平的响应差异,不同N添加水平下,菌根菌如何调控苗木根际土壤微生物养分限制,以期为全球气候变化背景下探究生态系统稳定性提供理论参考。
1 材料与方法
1.1 试验材料
2019年2月,将樟子松种子用2%的KMnO4溶液消毒30 min,然后用无菌水冲洗干净;在25 ℃恒温光照培养箱中催芽,待种子萌发后播入装有高温高压灭菌基质(蛭石与土体积比为1∶2)的花盆中(D=15 cm),并放入人工气候室培育(温度25 ℃,最大湿度60%,最大光照强度10 000 Lx)培养2月后待用。
1.2 试验设计
1.2.1 菌剂制备 选用樟子松优势菌根真菌:褐环粘盖牛肝菌(Suillusluteus)、厚环粘盖牛肝菌(Suillusgrevillei)、黄褐口蘑(Tricholomafulvum)、浅灰小牛肝菌(Boletinusgrisellus)、粘盖牛肝菌(Suillusbovinus)、球根白丝膜菌(Leucocortinariusbulbiger)、浅黄根须腹菌(Rhizopogonluteolus)和彩色豆马勃(Pisolithustinctorius)。上述8种菌剂均采用MMN培养液与蛭石配置成固体培养基,高温高压灭菌1 h,待冷却后分别接种外生菌根真菌,置于25 ℃培养箱内进行暗培养扩繁,25 d长满菌袋后备用。
1.2.2 接菌处理 2019年4月,设置2种接菌处理,分别为混合接菌处理(以下简称+M)和未接菌处理(以下简称-M)。接菌方法为:在育苗盆底部放入适量灭菌基质,然后取上述培养好的8种菌根真菌菌剂等质量混匀后,平铺于灭菌基质上,同时选取培养2个月的长势良好的苗木栽入育苗盆中,尽量使苗木根系与菌剂充分接触,每盆接菌量20.0 g,每盆栽植5株幼苗,每盆质量控制1.0 kg;对照处理加入经灭菌的20.0 g固体混合菌剂,同样方法栽植苗木,浇透水,将不同处理的育苗盆随机排布,置于内蒙古农业大学实验苗圃(111°43′15.88″E,40°48′49.48″N)温室大棚内培养。
1.2.3 模拟氮添加处理 2019年6月,测定+M处理苗木菌根侵染率达到了23.9%(-M处理苗木菌根侵染率为0%),开始模拟N添加试验。根据试验区N沉降背景值为34.3 kg·hm-2·a-1[10],设置4个N梯度处理:不施氮(0N,0 kg·hm-2·a-1)、低氮(LN,15 kg·hm-2·a-1)、中氮(MN,30 kg·hm-2·a-1)、高氮(HN,60 kg·hm-2·a-1)。用自来水溶解的KNO3和NH4Cl(NO3-∶NH4+为1∶1)作为N添加溶液,隔10 d定量施入不同浓度的N溶液100 mL·盆-1,施N操作分10次施入。接菌处理包括混合接菌和未接菌,N添加包括4个N梯度,共计8种处理组合,每个处理组合设置75株苗木重复,共培育600株苗木。采用喷洒方法将N施入育苗盆中,模拟降雨方式将N带入土壤中,同时施入也更为均匀。
1.3 土壤速效养分和胞外酶活性测定
2019年9月,N添加试验结束,间隔15 d后,测定N添加和接种处理下苗木单株生物量为0.21 g。各处理中分别取75株苗木的根际土壤,25株苗木的根际土混合1个土壤样品,每个处理3次重复。
土壤碱解N采用碱解扩散法;土壤有效P测定采用浸提-钼锑抗比色法;土壤速效K含量用火焰光度法测定[11]。土壤胞外酶活性的测定包括土壤C相关酶[β-1,4 葡萄糖苷酶(BG)]、土壤N相关酶[亮氨酸氨基肽酶和β-1(LAP)和4-N-乙酰-氨基葡糖苷酶(NAG)]、土壤P相关酶[碱性磷酸酶(ALP)],具体测定参考R.L.Sinsabaughetal[12]的方法,以对硝基苯酚的浓度表征土壤胞外酶活性[13]。
酶化学计量特征计算公式:
C∶NEEA=lnBG/ln(LAP+NAG)
(1)
C∶PEEA= lnBG/ln(ALP)
(2)
N∶PEEA= ln(LAP+NAG)/ln(ALP)
(3)
Vector L=SQRT((C∶NEEA)2+(C∶PEEA)2)
(4)
Vector A=DEGREES(ATAN2(C∶PEEA,C∶NEEA))
(5)
式中:C∶NEEA、C∶PEEA、N∶PEEA分别表示土壤酶化学计量C、N比;C、P比;N、P比。相对较长的向量长度(Vector L)表示更大微生物相对C限制;向量的角度(Vector A)<45°表示微生物相对N限制的程度;向量的角度>45°表示微生物相对P限制的程度[14]。
1.4 数据处理
利用SPSS 23.0(SPSS for windows,chicago,USA)对接菌和N添加处理进行双因素交互作用分析,并用LSD多重比较法检验各处理间苗木根际土壤理化指标、胞外酶活性及其生态化学计量特征的差异性。采用R语言4.0.2(R development core team2021)进行偏最小二乘路径模型(PLS-PM)路径分析,采用Sigmaplot10.0(systat software inc.,san jose,USA)做图。
2 结果与分析
2.1 接菌和N添加处理对樟子松苗木根际土壤速效养分的影响
接菌和N添加处理的交互作用显著影响樟子松根际土壤中有效P和速效K。不同接菌处理下,随N添加量增加根际土壤中碱解N均呈增加的趋势,菌根苗根际土壤有效P和速效K分别在HN和LN处理下达到最大(表1)。2种接菌处理下,LN、MN、HN处理间碱解N无显著差异,但均显著高于0N处理。-M处理下,N添加处理间根际土有效P无显著差异;而+M处理下,HN处理的有效P较0N、LN、MN处理分别显著增加了44.7%、61.8%、57.1%。-M处理下,LN处理的根际土速效K较HN处理降低了12.8%(P<0.05);而+M处理下,LN处理的速效K较HN处理增加了11.0%(P<0.05)(表1)。
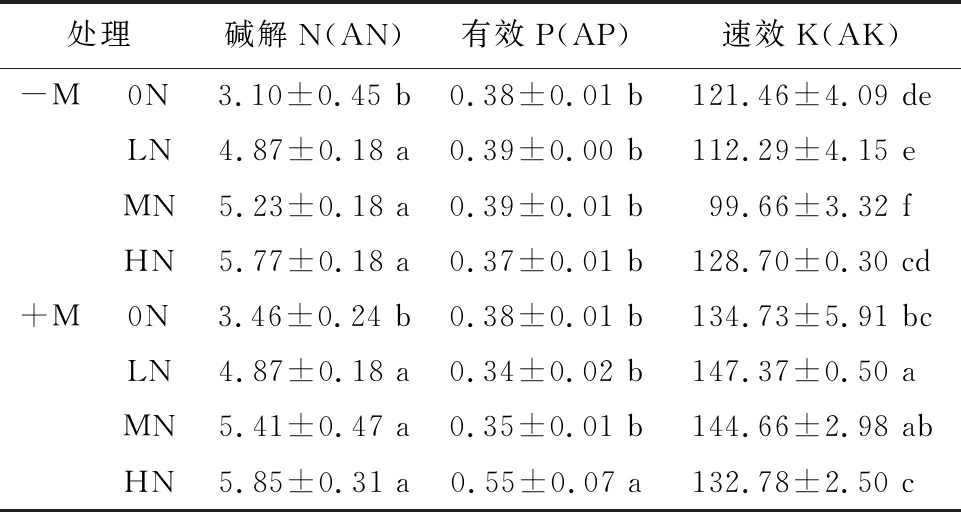
表1 接菌和N添加处理对樟子松根际土壤速效养分的影响
2.2 接菌和N添加对樟子松苗木根际土壤胞外酶活性的影响
接菌和N添加处理的交互作用显著影响LAP、NAG、ALP酶活性。-M处理下,随着N质量分数增加土壤中LAP酶活性呈下降的趋势,而NAG和ALP酶活性均呈增加的趋势;+M处理下,随着N质量分数的增加土壤中LAP、NAG、ALP酶活性均呈先增加后下降的趋势(表2)。LN、MN、HN处理下,+M处理的BG酶活性均显著低于-M处理。LN、MN处理下,+M处理的LAP、NAG和ALP活性均显著高于-M处理。HN处理下,+M处理的LAP酶活性较-M处理增加了60.8%(P<0.05),而+M处理的NAG和ALP酶活性较-M处理分别降低了18.6%(P<0.05)和30.3%(P<0.05)(表2)。

表2 接菌和氮添加处理对樟子松根际土壤胞外酶活性的影响
2.3 接菌和N添加对樟子松苗木根际土壤酶化学计量特征的影响
接菌和N添加处理的交互作用显著影响根际土壤酶化学计量特征。-M处理下,随N质量分数增加,根际土壤中C∶NEEA、Vector A呈增加的趋势,C∶PEEA和N∶PEEA呈下降的趋势,Vector L呈先增加后下降的趋势(表3)。+M处理下,随N质量浓度增加,C∶NEEA、C∶PEEA、Vector L呈下降的趋势,而N∶PEEA和Vector A无显著差异。0N处理下,+M处理的C∶PEEA和N∶PEEA较-M处理分别降低了14.7%(P<0.05)和18.5%(P<0.05),而+M处理的Vector A较-M处理增加了12.4%(P<0.05)。LN、MN、HN处理下,+M处理的Vector L较-M处理分别降低了27.0%(P<0.05)、32.4%(P<0.05)和28.3%(P<0.05)。MN、HN处理下,+M处理的Vector A较-M处理分别降低了5.5%(P<0.05)和10.6%(P<0.05)。-M处理下,0N、LN、MN根际土壤C∶N∶PEEA均为1∶1∶1,HN转变为1∶1∶2,而+M处理下,0N为1∶1∶1,LN、MN、HN转变为1∶1∶2(表3)。

表3 接菌和氮添加处理对樟子松根际土壤酶化学计量特征的影响
2.4 微生物养分限制与土壤速效养分和胞外酶活性的关系
通过PLS-PM路径分析N添加处理、接菌处理、土壤速效养分和胞外酶活性对微生物养分限制影响的拟合度为0.67(图1A)。接菌处理(0.47)和N处理(0.44)对微生物养分限制的直接影响均为正效应。接菌处理通过影响土壤速效养分和胞外酶活性,最终通过胞外酶活性正效应作用于微生物养分限制。N添加处理通过影响土壤速效养分作用于土壤胞外酶活性,间接影响微生物养分限制。影响微生物养分限制的因子总效应分析发现,接菌处理调控微生养分限制的最重要的路径,效应系数顺序为接菌处理>N添加处理>土壤速效养分>土壤胞外酶活性(图1B)。
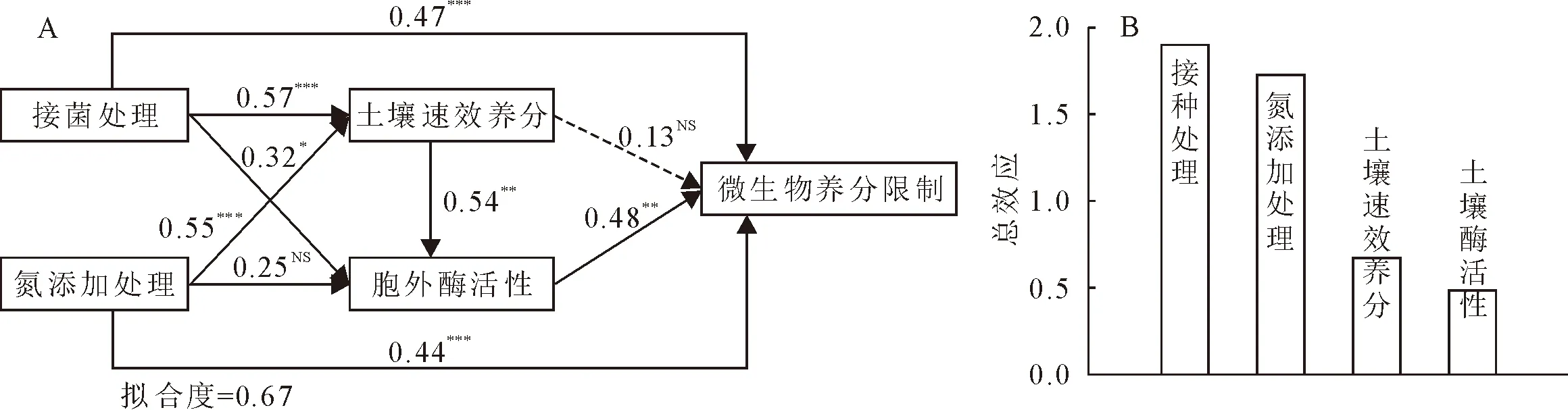
注:PLS-PM分析接种菌根菌和N添加处理下生态化学计量特征对微生物养分限制影响的主要路径(图A和B)。实线和虚线箭头表示正向和负向效应关系。箭头上的数字表示路径系数(*表示P<0.05,**表示P<0.01,***表示P<0.001,NS表示不显著)。
3 结论与讨论
3.1 接菌处理调控樟子松根际土壤微生态环境对N添加的响应
菌根共生体作为植物和土壤之间物质循环的纽带[15],对调控根际微生态环境的稳定性具有重要的作用[16]。不同接菌处理下,随N添加增加樟子松根际土壤碱解N呈增加的趋势,表明外源N输入已改变根际土壤的N平衡。研究也发现接种菌根真菌提高了高N环境中根际土壤有效P和速效K含量,原因可能是接种菌根真菌与植物形成共生体,且菌丝网络扩大了植物养分吸收范围,进而增强了植物根际微生物对高N添加的忍受能力[17]。土壤微生物与植物根系通过分泌根际土壤酶影响根际微生态环境中矿质元素的转化和分解[18-19]。研究发现,随着N质量分数的增加非菌根苗根际土壤中与N相关的酶活性呈下降的趋势,与P相关的酶呈上升的趋势,原因为外源进入土壤中减少土壤N相关水解酶的合成,N添加提高了植物对土壤P的吸收,导致土壤P相关水解酶增加。然而,N添加导致菌根苗根际土壤胞外酶活性均呈先增加后下降的趋势,表明菌根真菌改变根际土壤酶的原有的响应规律,原因可能为不同微生物生态策略存在差异,研究表明菌根真菌符合K生长策略,相较于细菌r生长策略,使生态系统相对稳定[20],因此接种菌根真菌提高了生态系统的稳定性。研究发现,一定范围的N添加,+M处理的LAP、NAG和ALP活性均显著高于-M处理(表2),表明N添加对苗木菌根的影响存在阈值,因为大量N输入一定程度上破坏菌根共生关系[13]。
3.2 菌根菌和N添加处理对根际土壤微生物养分限制的影响
土壤生态酶化学计量学反映了微生物对C、N和P的养分需求,在全球尺度和中国南北样带上,C、N和P获取酶的对数比趋近于1∶1∶1,表明土壤微生物营养元素的化学计量稳态[21-22]。研究发现,0N、LN、MN处理下非菌根苗根际土壤中C∶N∶PEEA均为1,表明微生物在低N输入的背景下,微生物营养元素获取酶之间趋于稳态,而HN添加打破了微生物养分平衡,以上结果也证明了根际微生态环境对N添加存在阈值范围。研究也发现随N质量分数增加,非菌根苗根际土壤微生物P限制增加。原因为非菌根苗随外源N输入,增加植物对土壤中P获取,导致土壤中微生物P限制增强[23]。研究发现,LN、MN、HN处理下菌根苗土壤微生物的养分获取酶生态计量关系同样被破坏,然而菌根苗根际土微生物P限制并无显著差异,一方面原因为菌根真菌改善了微生态环境[24],维持根际微生态环境的自稳态[25],另一方面原因为菌根真菌可以直接或间接影响微生物群落结构组成[26],其中土壤中磷酸盐溶解细菌提高了磷酸盐的溶解度[27],进而导致菌根苗根际土壤微生物P限制未发生明显改变。研究也发现,随N质量分数增加,非菌根苗根际土壤微生物C限制呈先增加后下降的趋势,而菌根苗呈下降的趋势。以上结果的原因可能是一定范围的N添加减少细根数量,同时增加根系寿命[28],导致土壤中C释放减少,根际微生物间C竞争增强,而高N输入破坏原有生态平衡[29],导致根际土壤微生物C限制降低。同时接菌处理促进植物生长,增强微生态环境对N添加的适应能力[30],因此,高N添加下接菌处理中土壤微生物C限制降低。通过路径分析也发现,接菌处理对微生物养分限制为正效应。因此,本研究发现菌根真菌有效调控了氮添加下根际微生态环境的稳定性,降低苗木根际微生物的C和P限制。
猜你喜欢
辽宁林业科技(2022年2期)2022-11-24
中国土壤与肥料(2022年4期)2022-06-14
中国食用菌(2022年5期)2022-06-08
林业科学(2022年2期)2022-05-11
草地学报(2022年4期)2022-04-25
四川农业科技(2021年7期)2021-12-01
农民致富之友(2019年8期)2019-05-22
农民致富之友(2019年4期)2019-03-13
北极光(2018年12期)2018-03-07
知识就是力量(2017年12期)2018-01-15
