
塞罕坝国家森林公园褐柳莺繁殖生物学研究
2022-10-08 03:16汪挥胜祃来坤王姣姣侯建华
四川动物 2022年5期
汪挥胜,祃来坤,2,王姣姣,侯建华*
(1.河北大学,河北 保定 071000;2.河北民族师范学院,河北 承德 067000)
鸟类生活史是生态学研究的重要领域,对生活史理论的发展作出了重要贡献(Partridge&Harvey,1988)。了解鸟类生活史特征对于理解其种群动态、个体特征以及温带、热带物种之间的差异具有重要作用(Martin,1995)。繁殖是鸟类生活史中最重要的组成部分,包括配偶选择、巢址选择、产卵、孵化和育雏(郑光美,2012),这些通过自然选择形成的一系列适应性行为,有利于个体和种群的生存(Saether & Bakke,2000)。但全球仅1/3的鸟类有较详细的繁殖资料,而雀形目Passeriformes不足7%(Xiao,2017),因此,繁殖资料的收集仍是现阶段鸟类研究中重要和长期的工作。
褐柳莺隶属于雀形目柳莺科Phylloscopidae,在河北塞罕坝国家森林公园为夏候鸟,活动于滦河岸边不同盖度和高度的灌木丛中,种群密度大(Liu,2017)。目前,仅有1篇关于俄罗斯远东地区褐柳莺应对花鼠捕食的反捕食策略的研究(Forstmeier &Weiss,2004),缺乏褐柳莺的完整繁殖信息。本研究于2021年5—8月对繁殖于塞罕坝国家森林公园的褐柳莺进行了详细研究,并采用全程录像的方法对其繁殖过程进行了观察。
1 研究区域与研究方法
1.1 研究区域
塞罕坝国家森林公园(116°51′~117°39′E,42°2′~42°36′N,海拔1 350~1 650 m)位于河北省最北端,植被群系主要包括落叶针叶林、常绿针叶林、针阔混交林、阔叶林、灌丛或灌草丛、草原与草甸等。属寒温带大陆性季风气候,年均气温-1.2℃;年均降水量452 mm,年均相对湿度68.8%;无霜期69 d,雪季可达7个月(Liu,2019)。根据褐柳莺的繁殖习性,调查范围集中在塞罕坝国家森林公园三道河口管理处与五间房之间的沿朝格都尔河河道两侧的林地、灌丛及苗圃地(图1)。
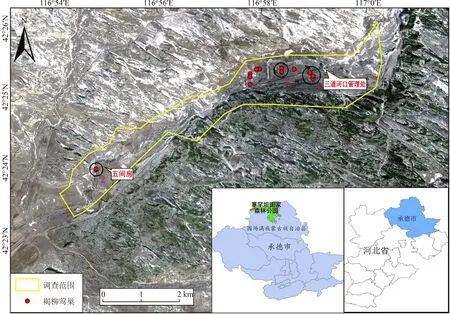
图1 研究区域及巢分布Fig.1 Study area and nest distribution
1.2 研究方法
2021年5—8月通过跟踪处于繁殖阶段的成鸟和系统搜索法寻找褐柳莺的繁殖巢(正在筑巢或者至少包含1枚卵或者幼鸟)(Martin & Geupel,1993),搜索区域包括研究区的针叶林、灌木林、灌草丛以及人工苗圃,找到后用GPS进行定位,共18个,对其中13个使用微型摄像机(HiLEME,中国)进行持续监测,其余5个采用隔天人工探查的方式进行监测,直到繁殖失败或者雏鸟离巢结束。
微型摄像机安装在可完整监视到整个巢口的树枝上或者人工辅助放置在巢旁的木棍上。收集和记录繁殖巢以下参数:(1)巢参数:巢材、巢距地面高度(cm)、巢所在树高(cm)、巢口直径(mm)、巢深(mm)、巢长(mm)、巢宽(mm)和巢高(mm);(2)卵参数:包括窝卵数、卵径长(L/mm)、卵径宽(B/mm)、卵体积(V=0.51LB)(Hoyt,1979)和卵重(g);(3)雏鸟参数(从雏鸟出壳日起,每隔2 d测量一次雏鸟身体参数,测量时间为每天下午):雏鸟体质量(g)、跗趾长(mm)、嘴峰长(mm)和雏鸟离巢日期;(4)繁殖成功巢指至少有1只雏鸟存活到离巢,繁殖成功率为繁殖成功巢占所有巢的比例(Yang.,2011)。长度采用游标卡尺(卡夫威尔,中国;精度:0.01 mm)测量,质量采用电子秤(DH,中国;精度:0.01 g)测量。为了减少干扰,所有巢参数的测量均在该巢繁殖活动结束后进行。
1.3 数据分析
2 结果
2.1 繁殖期及繁殖成效
褐柳莺于4月末至5月上旬到达研究区,5月底开始筑巢,繁殖活动持续到8月中旬,8月下旬到9月上旬开始陆续迁走,繁殖期持续约4个月,6—7月为营巢高峰期,88.89%(16/18)在这期间产下了第一枚卵。
褐柳莺繁殖成功率为27.78%(5/18)。在13个失败巢中,9巢被捕食,巢捕食导致繁殖失败占比69.23%。根据录像监控以及现场痕迹调查,6巢的捕食者为赤狐,1巢捕食者为牛头伯劳,其他巢的捕食者未被记录(表1)。
2.2 巢及巢址特征
18个褐柳莺巢为侧面开口的球形巢(图2),均分布在人工苗圃中,其中,云杉幼苗上9个,占总数的50.00%;樟子松幼苗上8个,占44.44%;地面1巢,占5.56%。巢位高度为(31.83±5.40)cm(=18),巢所在树高为(188.44±20.89)cm(=18)。巢口直径(56.51±2.08)mm、巢深(73.79±2.81)mm、巢长(132.23±3.30)mm、巢宽(106.07±4.68)mm、巢高(99.91±2.93)mm。巢距地面的高度显著低于巢距灌木顶部的距离(=-4.873,<0.01),表明巢筑在植被中下部。
2.3 产卵及孵卵行为
首枚卵出现在5月28日,最晚为7月14日。褐柳莺每天06∶00—09∶00产下1枚光滑无斑点的白色卵(图2)。窝卵数(4.00±0.15)枚(=10),主要为4枚(8/10);孵卵期为(14.00±0.45)d(=5);卵径长(16.88±0.16)mm(=17),卵径宽(13.11±0.16)mm(=17),卵体积(1 484.15±32.74)mm(=17),鲜卵重为(1.39±0.02)g(=17)(表1)。
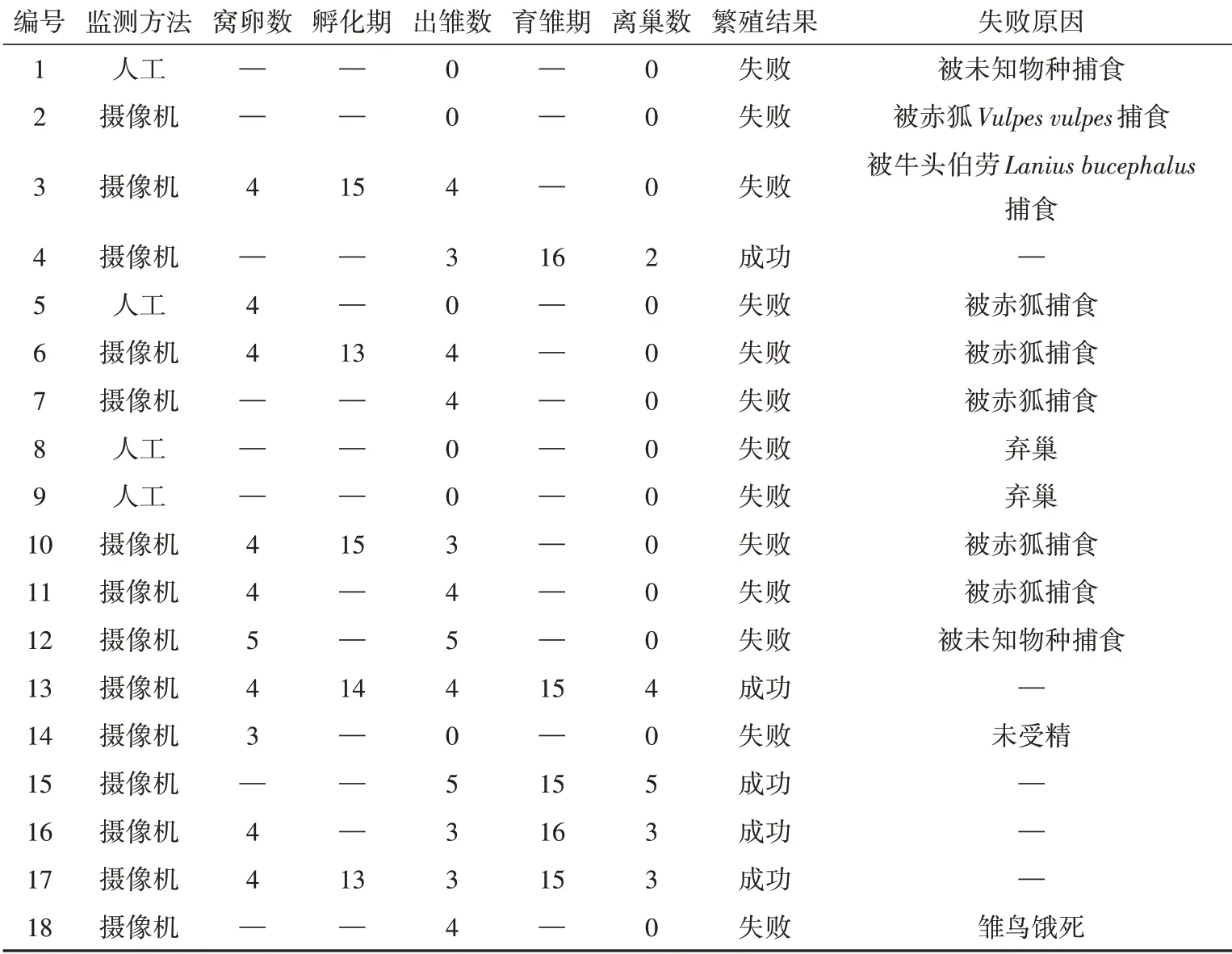
表1 2021年塞罕坝国家森林公园褐柳莺繁殖数据Table 1 Breeding data for Phylloscopus fuscatus in the Saihanba National Forest Park in 2021

图2 褐柳莺的巢址生境(左)、巢和卵(右)Fig.2 Nest-site(left),nest and egg(right)of Phylloscopus fuscatus
雌鸟产完最后1枚卵或倒数第2枚卵开始孵卵,开始孵卵后雌鸟在巢中过夜。根据2巢的完整录像发现亲鸟日孵卵频次在整个孵卵期没有明显的波动,而日孵卵时长波动较大,但每日04∶00—20∶00的总孵卵时长都大于600 min。不同巢的每小时孵卵频次和孵卵时长不同,但都表现出相同的变化趋势:黎明的孵卵频次低,时长较长;黄昏孵卵频次与其余时间无明显差异,但孵卵时长较长;每日孵卵的最短时长出现在09∶00—13∶00,从黎明到中午孵化时长呈递减趋势,中午到黄昏时长呈递增趋势(图3)。每日翻卵频次为(163.21±11.15)次(=4),各巢翻卵频次未呈现规律性变化。
2.4 育雏及雏鸟发育
育雏期为(15.4±0.2)d(=5)。根据录像,雏鸟破壳后卵壳被雌鸟叼出巢。在育雏过程中,亲鸟在雏鸟排出粪囊后,将粪囊叼离巢区。雏鸟孵出当日,亲鸟坐巢频次显著增加,在雏鸟孵出后0~5 d,亲鸟的坐巢次数都高于孵化期,但坐巢的时间总体呈下降趋势,直到雏鸟11日龄,亲鸟完全停止暖雏行为,再继续喂养雏鸟(5.6±0.2)d(=5),至雏鸟离巢(图3)。

图3 褐柳莺的孵卵/暖雏行为Fig.3 Incubation/brooding behaviour of Phylloscopus fuscatus
从每日育雏频次来看,从第一只雏鸟孵出,到雏鸟离巢的前一天,亲鸟每日喂食频次逐渐增加,直至雏鸟离巢当日,喂食频次才有所减少。在每小时喂食频次上,05∶00—20∶00的每小时喂食频次并未出现较大波动,但04∶00—05∶00的喂食频次明显少于其他时间段,而总体呈现出黄昏喂食频次略低于其他时间段。每日清巢次数为(30.90±1.93)次(=2)。雏鸟排便总是在亲鸟喂食后发生,雏鸟排便的频次和亲鸟喂食的频次高度相关(r=0.705,<0.01)(图4)。

图4 褐柳莺喂食行为Fig.4 Feeding behaviour of Phylloscopus fuscatus
雏鸟部分日龄外部形态如下:1日龄雏鸟闭着眼睛,皮肤大部分裸露无毛,仅头顶和眼周有少量绒毛;皮肤肉红色,眼泡黑色且突出,不能站立,蜷缩成团。3日龄雏鸟皮肤颜色加深,背部及翅膀皮下出现黑色羽管。6日龄雏鸟头部呈黑色,背羽、尾羽和初级飞羽开始生长。9日龄雏鸟飞羽和头部羽毛完全覆盖皮肤,仅背部有皮肤裸露,并且能够伸长脖子乞食。12日龄雏鸟全身被羽,飞羽前端为蓝色,末端为黑色,能够长久站立。15日龄雏鸟羽毛和成鸟相似,颜色比成鸟略深,尾羽较短。
Logistic回归拟合雏鸟跗趾长和体质量的生长曲线,获得3个身体参数的生长曲线拟合方程,体质量的为:=9.465 87/[1+14.844 94exp(-0.571 79)],跗 趾 长 的 为:=25.120 52/[1+3.794 8exp(-0.3)],嘴 峰 的 为:=6.740 56/[1+2.791 7exp(-0.263 63)](图5)。
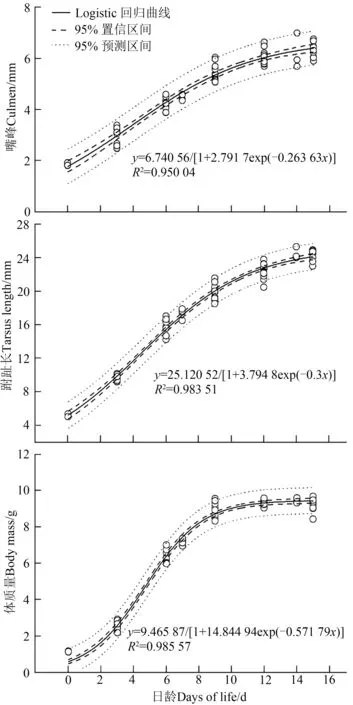
图5 褐柳莺雏鸟不同部位的生长曲线Fig.5 Growth curves of different body parts of Phylloscopus fuscatus nestlings
3 讨论
褐柳莺迁徙到塞罕坝地区繁殖的时间比更高纬度的俄罗斯远东地区的繁殖种群(5月末迁入)更早,而迁走时间与俄罗斯远东地区繁殖种群相近,这可能与不同纬度地区的气候以及迁徙距离差异有关。塞罕坝地区繁殖的褐柳莺种群越冬地为中国南部和东南亚地区,而俄罗斯远东地区繁殖的褐柳莺种群越冬地同样位于东南亚地区(von Blotzheim,1966)。在其他鸟类中发现,高纬度地区繁殖的种群需经过更长的迁徙路线从越冬地到达繁殖地(Burman,2018),相比于塞罕坝地区的种群,在俄罗斯远东地区繁殖的种群可能需要迁徙更远的距离,同时,繁殖地的气温也会影响越冬地鸟类开始春迁的时间(Kanerva,2020),俄罗斯远东地区春季回暖时间更晚。
鸟类窝卵数是鸟类学家长期以来一直探讨的问题,也是鸟类生活史对策理论的重要研究内容之一。窝卵数变异原因主要包括地理变异、季节性变化、其他因素(栖息地类型、种群密度等)(郑光美,2012)。俄罗斯远东地区纬度为59°51′N,褐柳莺的窝卵数平均为5.16枚(Forstmeier,2001);塞罕坝地区纬度为42°36′N,窝卵数平均为4.00枚,这与大多数研究一致,即窝卵数随纬度增加 而 增 加(Griebeler,2010;Winkler,2014)。目前关于窝卵数地理变异原因的讨论集中在资源供应、气候差异以及捕食风险3个方面(Bourgault,2010;Mérő,2014)。受塞罕坝地区与俄罗斯远东地区的纬度差异以及林地类型影响,塞罕坝地区窝卵数较俄罗斯地区小可能同时受到以上3个方面的影响。资源供应方面:塞罕坝地区为次生人工林地,相较于俄罗斯远东地区的原始林地,褐柳莺可获得的食物资源可能会更少;气候差异方面:俄罗斯远东地区的纬度更高,气温回暖时间更晚,因此褐柳莺迁到的时间更晚,而迁走时间相近,导致可用于繁殖的时间更短。在繁殖结束后,俄罗斯远东地区褐柳莺种群需要迁徙更长距离到达越冬地,在更长距离的迁徙中,幼鸟的死亡数量可能会高于迁徙距离更短的塞罕坝种群,因此俄罗斯远东地区更大的窝卵数可以更好维持种群数量稳定。从捕食风险来看:塞罕坝地区褐柳莺更小的窝卵数不仅适应了低纬度地区更高的巢捕食风险,同时减小了因巢捕食带来的损失。塞罕坝更长的繁殖季节,让其在首次繁殖失败后,有机会进行第二次繁殖,从而进一步减小巢捕食对种群数量稳定带来的影响。
孵卵投资是鸟类孵卵行为中最受关注的方面,亲鸟需权衡在觅食和孵卵之间的时间分配,因为它们需要在维持自身需求的同时保证胚胎的正常 发 育(Cooper & Voss,2013;Dammhahn,2018)。由于褐柳莺的雌雄二性区别不明显,无法通过视频准确判断雌性是否单独孵卵,但所有巢的监控记录中均未观察到交替孵化行为,且存在其他成鸟向孵卵成鸟递食的现象;同时在成鸟孵卵过程中,巢所在领地仍有雄性鸣唱,因此推测孵卵行为可能由雌性单独完成。根据白天每小时孵化时间的分配情况,亲鸟采取高频次、单次孵卵时间短的孵化策略,并且在09∶00—15∶00投入最少的时间,表明雌性觅食主要发生在这个阶段。这种孵化策略可能是亲鸟对自身需求以及胚胎发育需求进行权衡的结果。这种时间分配模式非常合理,中午由于气温上升,卵的温度会比上午和傍晚更高,此时昆虫也更加活跃,分配更多时间觅食既能保证卵的温度不会太低导致胚胎死亡,也能保证亲鸟获取充足的食物。Drent等(1970)发现,由于胚胎在孵化后期需要更多的热量,亲鸟会减少孵化频次,增加总体的孵化时长,但是本研究在孵化阶段没有发现这种后期孵化时长增加的现象。
清理粪囊是育雏期的一种重要行为,也是清巢行为的重要组成部分,不仅可以减少雏鸟被寄生虫感染的风险,亦可以降低捕食者通过粪便气味和痕迹定位巢的概率(Guigueno & Sealy,2012;Ibáñez-Álamo.,2014)。Quan等(2015)发现雀形目雏鸟排便和亲鸟喂食具有高度的协同性,雏鸟总是在亲鸟喂食后就进行排便,以此保证亲鸟能够及时叼走粪囊,本研究与上述结果相似,褐柳莺雏鸟的排便与亲鸟喂食行为较为统一。
繁殖成效分析表明,本地区褐柳莺的繁殖成功率仅为27.78%,其中巢捕食是导致繁殖失败的主要原因,这与许多柳莺属spp.鸟类报道的捕 食 率 相 似(Smith,2007;Roncalli,2016)。本地区主要巢捕食者为赤狐,这可能与赤狐在研究区较高的遇见率相关;此外,巢址选择也是影响巢捕食的重要因素(Jara,2020),本研究中褐柳莺主要在人工苗圃地中繁殖,这与其他地区的筑巢生境不同(Forstmeier & Weiss,2004),这种巢址选择模式是否是褐柳莺应对高捕食压力的繁殖对策以及对繁殖成效的影响需要进一步研究。
猜你喜欢
中国畜牧杂志(2022年2期)2022-02-20
东方娃娃·保育与教育(2021年10期)2021-12-08
疯狂英语·新阅版(2021年5期)2021-06-21
发明与创新·中学生(2018年11期)2018-11-30
国外畜牧学·猪与禽(2018年11期)2018-05-14
伙伴(2018年1期)2018-05-14
百科知识(2016年20期)2016-12-24
农村百事通(2016年19期)2016-11-22
农村农业农民·B版(2015年9期)2015-10-16
