
高油酸油菜遗传育种研究进展
2022-10-11 03:04咸志慧龙卫华谭筱玉胡茂龙浦惠明
生物技术进展 2022年5期
咸志慧,龙卫华*,谭筱玉,胡茂龙,浦惠明
1.江苏省农业科学院经济作物研究所,南京210014;
2.江苏农林职业技术学院农学园艺学院,江苏 镇江212400
油菜(主要指甘蓝型油菜,Brassica napusL.)是我国重要的油料作物之一,常年种植面积7.0×106hm2,居全球油菜生产国种植面积首位[1]。菜籽油是油菜生产的最主要农产品,也是我国居民消费最广泛的植物油种类之一。脂肪酸几乎是菜籽油的全部组分,决定了菜籽油的品质。脂肪酸改良一直是油菜品质改良的主要内容[2]。
21世纪初,我国油菜育种界通过引进特异种质资源改良原有油菜品种,基本实现了所有审定油菜品种的双低化(菜籽饼低硫苷和菜籽油低芥酸),实现了我国油菜生产的第三次飞跃,同时完成了我国菜籽油的第一轮品质改良[3]。由于双低菜籽油中芥酸含量急剧降低(低至<1%),菜籽油的脂肪酸组分得到优化,其中不饱和十八碳脂肪酸(C18∶1≈62%,C18∶2≈19%,C18∶3≈9%)总含量超过90%,饱和脂肪酸(主要为C16∶0和C18∶0)仅占约7%,因此十八碳脂肪酸的改良成为新一轮菜籽油品质改良的目标。
高油酸(C18∶1)菜籽油一般是指油酸相对含量超过75%的菜籽油[4]。与普通双低菜籽油相比,高油酸菜籽油具有三大优势:①货架期较长,耐贮藏性更优。在十八碳多不饱和脂肪酸中,亚油酸(C18∶2)和亚麻酸(C18∶3)是人体的必需脂肪酸,其中亚油酸有2个双键,亚麻酸有3个双键,两者化学结构均不稳定,极易氧化分解而变味,产生对人体有害的物质(包括反式脂肪酸)。高油酸菜籽油中亚油酸和亚麻酸总量降至10%左右,在满足人体所需的脂肪酸前提下双键含量更少,成分更稳定。②对人体健康更有益,保健功能也更强。医学研究表明,高油酸在降低对人体有害的低密度脂蛋白含量的同时,不影响有益高密度脂蛋白的含量,减少胆固醇形成,预防人体心血管疾病。国外流行病学调查发现,地中海地区国家的居民比较长寿,心血管病发病率较低,这与当地人们普遍食用富含油酸的橄榄油有关[5]。③烹饪特性更好。高油酸菜籽油在加热到较高温度时不冒烟,既适用于家庭烹调,也适用于餐饮业加工一些存放时间较长的快餐食品和糕点[6]。④高油酸菜籽油因其甲酯化程度高,燃烧值高,更有利于生产生物柴油[7]。因此,提高油酸含量,同时降低多不饱和脂肪酸和饱和脂肪酸的含量,成为菜籽油品质改良的主要目标。当前我国高油酸遗传育种已取得不少成果,本研究将围绕高油酸油菜的种质创制、遗传分析、基因克隆及品种选育等方面进行总结,以期为高油酸油菜品种选育提供参考。
1 高油酸油菜种质资源创制及遗传模式
在传统的油菜种质资源库中并未发现高油酸种质资源的存在[8],因此各国科研人员试图通过人工诱变、基因干扰、基因编辑等传统或现代生物技术来创建油菜高油酸资源,相关研究取得了一系列的成果。
高油酸油菜种质资源创建常用的方法是人工诱变,其中物理诱变和化学诱变是两种主要诱变方式。美国科学家最早采用5%甲基磺酸乙酯(ethyl methyl sulfone,EMS)溶液处理油菜种子,在M2代发现了低多不饱和脂肪酸株系,F4代筛选到油酸含量大于87%的突变体[9]。1995年Rukker等[10]报道了利用EMS诱变获得了两株油酸含量分别为74.1%和74.6%的油菜品系。随后,有研究采用两次EMS诱变的方法逐渐提高油酸含量,得到了两株油酸含量分别为76.1%和76.6%的品系[11]。我国学者也利用不同方法开展高油酸油菜诱变研究。例如,在油菜小孢子培养过程中,用EMS诱变处理适期发育的小孢子后再对小孢子苗单株产生的种子进行筛选,获得了油酸含量为80.3%的高油酸突变体材料[12]。利用8~10万伦琴60Co射线辐射双低油菜干种子,并对辐射后代进行高油酸极端选择,近红外方法检测发现M3代油酸含量迅速提高,多数植株油酸含量在70%以上,最高油酸含量达93.5%[13]。以高浓度EMS溶液(1.5%)诱变处理油菜种子,在M1代就筛选到油酸 超 过70%的 单 株[14]。以 低 浓 度EMS溶 液(0.4%)诱变油菜得到突变体库,在M3代也找到油酸含量为75.02%的高油酸突变株系[15]。此外,通过航天诱变的方法获得了一个油酸含量达87.22%的突变株系[16]。以萌动发芽油菜种子为对象,利用60Co-ɣ射线辐照结合在M6代筛选得到一系列油酸含量稳定在73.51%~85.47%的后代株系[17]。以上研究结果表明,利用人工诱变筛选高油酸种质的思路和方法是可行的,诱变效率也相对较高。
此外,利用生物技术(包括基因干扰、基因编辑)也成功创制了多个高油酸种质资源。自2000年首次报道了利用共抑制技术获得油酸含量高达89%的油菜株系后[18],国内外学者先后采用RNA抑制技术(RNAi)得到了一系列油酸含量显著提高的株系,如干扰BnFAD2获得了油酸含量高达约80%的油菜转基因植株[19-22]。随着基因编辑技术在油菜上的利用,各国学者也得到成功的结果[23-25]。
阐明油菜高油酸性状的遗传模式,对于其育种利用至关重要。以高油酸亲本(C18∶1含量大于75%)和普通油酸亲本构建DH群体,对两年三点的油酸含量进行遗传力分析表明,高油酸性状遗传力达到0.99,环境因素影响极小[26]。通过对高油酸亲本DMS100(C18∶1=77%)与普通品系衍生DH群体的油酸含量性状进行QTL定位,发现其高油酸性状主要由一个主效位点控制[27]。以高油酸(含量87.22%)自交系Y539(C18∶1=87.22%,C18∶3=1.94%)与普通油酸杂交构建F2群体,通过高油酸性状紧密分子标记确定其高油酸性状是由A05和A01上的两个主效位点控制[16]。利用高油酸品系SW Hickory(C18∶1=78%)与普通油酸杂交构建DH群体,通过QTL定位确定其高油酸性状位于A5染色体上,并根据这个主效QTL开发了紧密连锁分子标记[28]。利用高油酸亲本H005(C18∶1=83.10%)与普通油酸含量亲本构建了六世代遗传分析群体,结果表明H005的高油酸性状由两对主效基因控制,且具有加性和显性效应[29]。以高油酸种质B161(C18∶1≈85%)与3个不同遗传背景的常规品系构建油酸的六世代遗传分离群体,混合遗传分析表明,该种质中高油酸含量性状均由两对主效基因控制,加性效应为主要遗传效应[17]。从现有高油酸性状的遗传模式结果看,油酸含量在75%~80%的种质资源,其高油酸性状一般由一对基因控制;油酸含量大于80%的种质资源,其高油酸性状一般由两对基因控制。
2 油菜高油酸基因的克隆与突变位点
油菜为异源四倍体植物,在其基因组测序完成之前进行基因克隆的难度较大,但模式生物拟南芥脂肪酸的合成调控途径为解析油菜脂肪酸含量调控途径提供了参考。脂肪酸合成过程大致为:乙酰辅酶A作为脂肪酸碳链的基本单元,通过还原、脱水和还原等步骤循环逐步合成长链(16-或18-碳)植物脂肪酸。位于质体和内质网中的各种去饱和酶负责催化长链脂肪酸变成单不饱和(棕榈烯酸C16∶1和油酸C18∶1)或多不饱和(亚油酸C18∶2和亚麻酸C18.3)脂肪酸,其中Δ-12脂肪酸去饱和酶2(Δ12-FAD2)可将油酸前体转化为亚油酸前体,它也是油料作物种子中油酸含量的主要控制酶[30]。
在油菜中,虽然也有不少研究者进行了Bn-FAD2基因克隆[31-33],但直到2013年才首次系统研究了BnFAD2多个同源基因的特征[34]。油菜中4个BnFAD2基因分别为BnFAD2-A5、BnFAD2-C5、BnFAD2-A1和BnFAD2-C1,其中位于A基因组的两个同源基因来源于白菜(Brassica rapaL.),另外两个位于C基因组的同源基因来源于甘蓝(Brassica oleraceaL.)。位于A5和C5染色体上的Bn-FAD2均为组成型表达,在幼苗、叶片、茎干、根、花等不同组织以及种子不同发育时期均有表达;位于A1上的BnFAD2缺失一段序列成为非功能基因;而位于C1染色体上的BnFAD2在不同发育时期的种子中均有表达,在根、花和角果皮中保持极低表达,为种子特异性诱导表达基因[35]。随后对3个BnFAD2基因的功能元件进行了深入研究。采用酵母功能互补实验进行功能验证,发现Bn-FAD2-A5和BnFAD2-C5基因的去饱和能力接近,均大于BnFAD2-C1基因。BnFAD2-A5和BnFAD2-C5是影响油菜种子油酸积累的主效基因[36]。采用Western blot技术分析发现,BnFAD2-C5启动子区域-1 257~-1 020 bp和-319~-1 bp能够诱导报告基因在种子发育中期高效表达,其内含子(+631~+1 033 bp区域)具有增强启动子转录的功能[37]。BnFAD2A5在其5′-非翻译区含有一个内含子(1 192 bp),该内含子具有启动子活性,-220~-1 bp是最小启动子区,-220~-110 bp和+34~+285 bp是赋予高转录水平的两个重要区域。GATA家族的转录因子可以与BnFAD2-A5和BnFAD2-C5启动子中的GT元件发生互作产生正向调节基因[38]。BnFAD2-C1的调控区存在潜在顺式作用元件,通过茉莉酸诱导处理可使其基因表达量发生变化[39]。
为了进一步发现能够影响油菜油酸含量的其他基因,科学家进一步利用全基因组关联分析、QTL定位等方法获得了其他油菜高油酸控制基因。尹明智等[40]发现DOF转录因子含有一个独特的富含Cys残基的单锌指DNA结合区域。将干扰载体pADOF1成功转入野生型拟南芥后,发现种子油酸含量明显上升,说明Dof转录因子在植物油酸代谢途径中有重要作用[40]。一个CCCH型转录因子BnZFP1通过提高diacylglycerol O-acyltransferase 1(Dgat 1)基因的表达,从而提高油酸的含量[41]。利用GWAS技术对375个双低油菜品系的油酸含量进行分析后发现了一个新的QTL位点,其候选基因编码一个金属离子结合蛋白,能使油酸含量提高3~5个百分点[42]。吲哚-3-丙酮酸单氧合酶基因BnaYUCCA10在角果期及开花5 d的花中的表达量与油酸含量呈极显著正相关,在高油酸材料中的表达量显著高于非高油酸材料的表达量,推测BnaYUCCA10基因可能参与种子的油酸代谢[43]。由此表明,影响植物油酸合成的基因可能并未被完全发掘,需要进一步探索。
油菜全基因组测序以及生物技术的发展有利于研究高油酸种质资源的功能基因以及功能基因上的突变位点(表1)。利用RNA干扰BnFAD2得到了高油酸油菜株系,表明BnFAD2是显著提升油菜油酸含量的关键基因。通过对不同油菜高油酸突变体控制基因的分析发现,目前大部分的高油酸种质,其控制基因均为BnFAD2,而且大部分突变位点均发生在BnFAD2-A5和BnFAD2-C5这两个基因上。
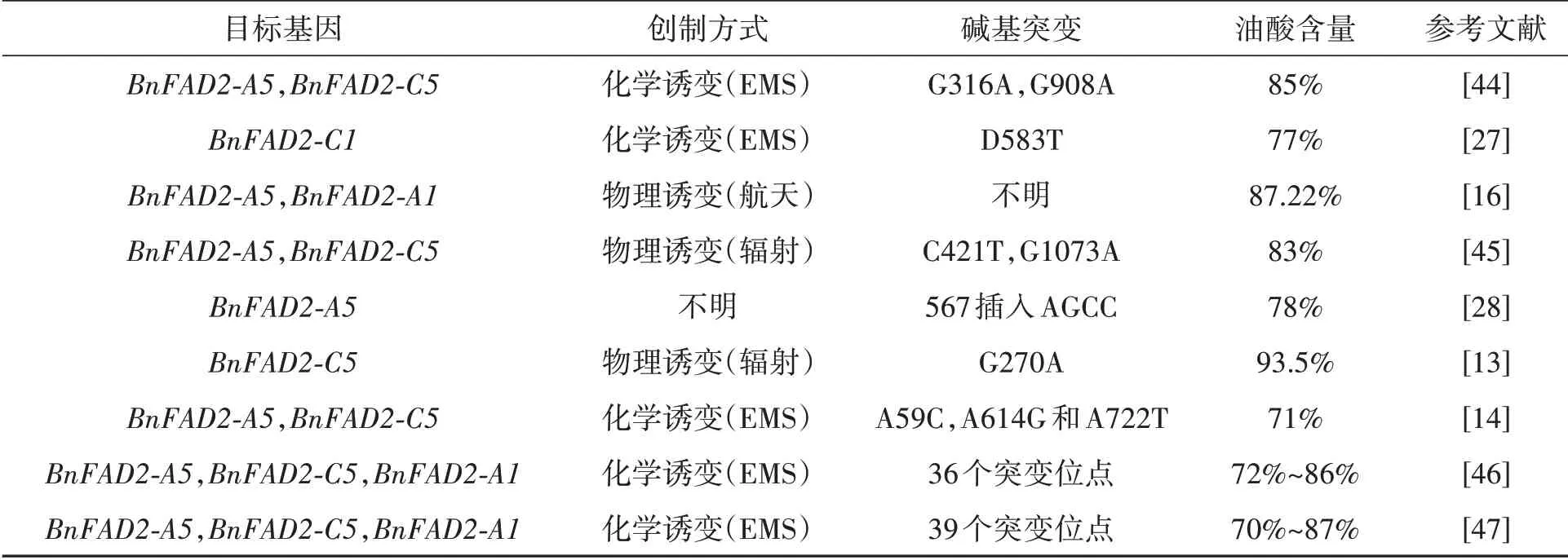
表1 非转基因油菜高油酸种质的BnFAD2突变Table 1 BnFAD2 mutation in non-transgenic rapeseed germplasm with high oleic acid
3 高油酸品种创制以及我国高油酸品种发展趋势
国际上以欧美国家最早开展高油酸油菜品种的选育工作。2004年,加拿大Cargill种子公司推出CNR604(油酸含量75%)、CNR603(油酸含量85%)等品系参加品比试验[48]。澳大利亚最早于2007年推出两个替代品种Cargill102和Cargill103[49]。欧洲第一个高油酸品种SPLENDOR于2004年在法国注册,油酸含量为76%,且具有较好的农艺性状(根系正常)。欧洲油菜育种家也将高油酸性状转入萝卜质不育系统培育三系杂交高油酸品种。
我国于21世纪初开展高油酸油菜品种的培育工作。湖南农业大学、江苏省农业科学院、浙江省农业科学院、西南大学等均自行创制了高油酸种质;浙江省农业科学院、湖南农业大学、华中农业大学、江苏省农业科学院、中国农业科学院油料作物研究所等单位陆续登记了高油酸新品种。浙江省农业科学院于2015年审定了高油酸品种浙油80,油酸含量超过80%,并且推出了“爱是福”高油酸健康菜籽油品牌[50]。2017年,华中农业大学在湖北省荆门市大面积种植的高油酸菜籽油获得成功,并与油脂加工公司合作开展高油酸油菜订单农业,产业化规模逐渐加大[51]。湖南农业大学与衡阳市合作发展高油酸油菜产业,组建了高油酸油菜产业技术创新战略联盟[52]。江苏省农业科学院利用自创高油酸种质转育骨干系,于2020年登记了高油酸三系杂交油菜品种益康1号(GPD油菜(2021)320278),油酸含量高达82.01%,目前该品种已在多点示范种植。与此同时,国内多个大型油脂加工厂已经表示出对高油酸油菜的极大兴趣,正在对接各高油酸油菜品种研究单位进一步开展合作。
4 当前高油酸油菜品种的不足及改良途径
尽管国内外均有高油酸油菜品种的登记和商业化生产,但这些品种的缺点是产量水平较低。油菜有3个BnFAD2同源基因能正常发挥功能,但目前可查询的商业化高油酸油菜品种主要利用BnFAD2-1和BnFAD2-2这两个组成型表达的高油酸突变基因。一个可能的原因是,这两个同源基因的组成型表达特征,高油酸品种的各个组织在全生育期油酸含量低于正常值。当遇到低温时,高油酸油菜较普通油菜细胞中脂肪酸不饱和度较低,抗低温凝固性能较差,营养器官细胞生长更容易受到抑制,故而在表型上表现为叶色更绿、生长缓慢等。正是由于营养阶段生物量的降低,造成高油酸油菜在花期和结角期植株相对较小、角果量较少,产量较低。
高油酸品种的改良,一是要保证高油酸基因不丢失,确保高油酸性状;二是要持续提高其产量水平。基于此,我们提出了高油酸品种的改良途径。①筛选高效实用型分子标记,防止基因丢失。油菜BnFAD2-1和BnFAD2-2虽位于不同的染色体上,但其表达框同源性超过97%。如利用分子标记进行后代单株筛选,必须设计高效准确的分子标记。②将高油酸基因转育至多种遗传背景。国内高油酸品种培育工作开启不久,育种体系尚未规模化。将高油酸基因转育至更多的遗传背景进行产量水平鉴定,是当前高油酸油菜育种工作亟需解决的问题。③进一步利用新的高油酸基因进行资源创制工作。先前研究中,BnFAD2-C1为种子特异型表达基因,但其突变体报道较少。如果对其开展定向突变工作,有可能获得营养生长不受影响的油菜种质,或许其产量性状与普通油菜无异。另外,随着基因编辑技术的兴起与应用,也可以对其他类型的油酸控制位点或基因开展定向突变工作。④与杂种优势利用途径相结合进一步提高产量。杂种优势利用是公认提高油菜单产的方式,将高油酸基因转育至杂交亲本中,利用杂种优势来提高高油酸品种的产量。
猜你喜欢
中国种业(2022年2期)2022-11-17
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
农村百事通(2022年16期)2022-07-06
煤炭与化工(2022年5期)2022-06-17
江苏农业科学(2022年3期)2022-03-03
中国种业(2022年1期)2022-01-27
中国种业(2022年1期)2022-01-27
环球时报(2020-11-26)2020-11-26
环球时报(2018-03-01)2018-03-01
