
羊毛性状相关蛋白、信号分子及基因的研究进展
2022-10-12 13:17冯梦雨姬凯茜伍修锟辛国省
中国畜牧杂志 2022年10期
冯梦雨,姬凯茜,伍修锟,辛国省,杨 果*
(1.宁夏大学农学院,宁夏银川 750021;2.中国科学院西北生态环境资源研究院,甘肃兰州 730030;3.甘肃省寒区旱区逆境生理与生态重点实验室,甘肃兰州 730030;4.宁夏大学生命科学学院,宁夏银川 750021)
羊的皮毛主要是用来保暖御寒,减少物理伤害,随着自然选择与人工驯化逐步进化。羊毛是纺织业的重要原料,羊毛品质直接影响养羊业和毛纺工业的发展,决定羊毛品质的重要羊毛性状包括绒长、细度、强度和卷曲度等。随着居民生活水平提高,市场对高端羊毛制品的青睐度越来越高,培育出优质高产毛用性状的羊是养羊业发展的必然趋势,然而我国现有的羊毛生产能力尚未达到稳定生产高品质羊毛水平,提高羊毛产量、改善羊毛品质亟待解决。随着现代分子生物学的发展,一系列与羊毛性状密切相关的蛋白、信号通路以及基因被陆续发现。本文从羊毛、毛囊结构的生长发育特点、角蛋白(Keratin,KRT)等相关蛋白、Wnt 家族、成纤维细胞生长因子家族(FGFs)等毛囊生长调控的信号分子以及相关基因等方面进行综述,以期为今后羊毛产业与毛用羊品种的选育提供理论参考。
1 羊毛、毛囊的结构和发育
羊毛大由两层毛构成,包括由次级毛囊(Secondary Hair Follicle,SF)生成的细毛和由初级毛囊(Primary Hair Follicle,PF)生成的粗毛,细毛为无髓毛,粗毛为有髓毛。羊毛品质是由多种因素决定的。Lv 等发现绵羊孕期的营养限制会抑制毛囊发育,导致毛囊密度降低。
羊皮由表皮和真皮构成,通过皮下疏松结缔组织与深层组织相连。表皮极其薄,由外到里依次为角质层、颗粒层、棘细胞和基底层。真皮可分为毛囊层和网状层,毛囊层以疏松结缔组织为主,含有皮脂腺、毛囊和汗腺等等结构。毛干是由皮质层、有或者无髓质和角质层构成,毛干外侧是内根鞘(Inner Root Sheath,IRS),由内根鞘小肋、赫胥黎层和亨利层3 个连续层组成。毛囊由内向外为内根鞘、伴随层(Companion Layer,CL)和外根鞘(Outer Root Sheath,ORS)等组成连接到皮肤,如图1 所示。
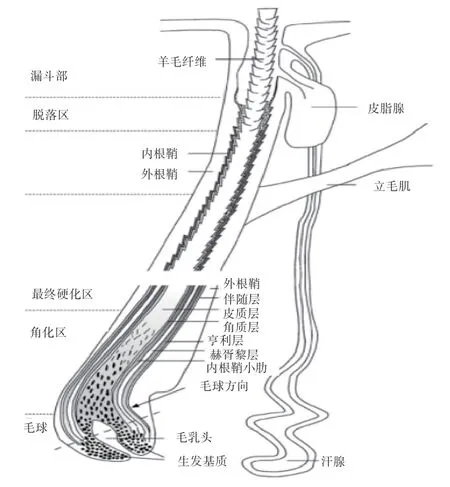
图1 毛囊生长期的结构示意图[7]
根据毛囊发育的特点,分为形态发生和周期性再生2 个阶段。第一阶段,毛囊在胚胎发育过程中形成,出生后不久,形成成熟毛囊,大约在妊娠期第70 天首先形成初级毛囊,随后形成次级毛囊,妊娠期第105 天时,初级毛囊周围可见双子叶皮脂腺,次级毛囊芽向下延伸并深入真皮,皮脂腺开始形成,第135 天时,初级毛囊周围已形成完整的皮脂腺和汗腺,次级毛囊仅有不发达的皮脂腺,出生后35 d 长出毛干。第二阶段,毛囊成熟并形成比较复杂的结构,毛囊分为两部分:一个是上部恒定部,另一个是下部循环部,主要包括含有毛囊基质细胞的毛球。在生长初期,毛乳头细胞能激活增殖的信号通路诱导毛囊基质细胞增殖,生长末期,增殖分化速度减慢,毛囊细胞开始凋亡,随后为退行期,毛囊变得凹陷,毛球收缩向上移动,包括毛囊基质细胞在内的细胞都经历了凋亡过程。最后为休止期,大部分细胞活动停止,毛囊处于静息状态,在这一时期结束时,毛球再生,毛囊重塑到生长期状态,开始下一个生长周期,即毛囊的周期性再生,如图2 所示。
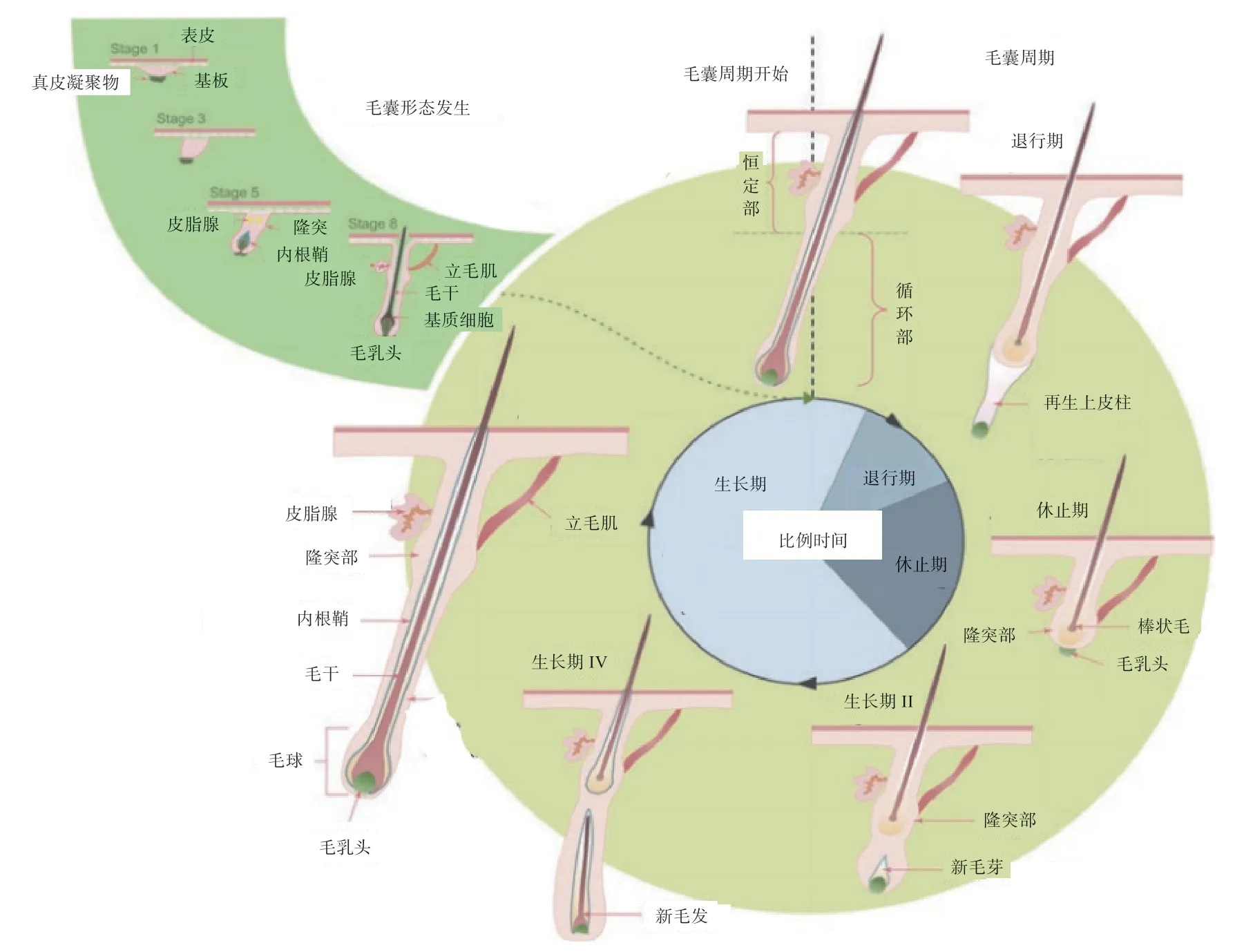
图2 毛囊形态发生和周期性再生过程示意图[8]
毛囊产生的纤维差异也是羊毛性状存在差异的主要原因,如粗毛羊表现为长而粗的纤维,细毛羊表现为均匀的细纤维。
2 与羊毛性状有关的关键蛋白
角蛋白有-角蛋白和-角蛋白2 种类型。羊毛的主要成分是-角蛋白。1935 年Goddard 等对羊毛蛋白质进行鉴定和分类,主要分为S-羧基甲基角鲨胺A(SCMK-A)和S-羧基甲基角鲨胺B(SCMK-B)2 类。Marshall 等在1982 年发现蛋白SCMK-As。随后认为SCMK-As 是角蛋白中间丝(Keratin Intermed iate Filaments,KIFs),SCMK-Bs 是角蛋白关联蛋白(Keratin-Associated Proteins,KAPs)。KRT 形成KIFs,KAPs 形成交联中间丝的基质,形成羊毛纤维。因此,羊毛的基本特性是由KIFs 和KAPs 决定的。
2.1 KIFs KIFs 主要在皮质中合成,分为I 型角蛋白中间丝(KIF1s)和II 型角蛋白中间丝(KIF2s)2 种类型,分别由和基因家族编码。羊毛的I 型角蛋白中间丝(KIF1s)呈酸性,其大小范围为392~416 个氨基酸,而II 型角蛋白中间丝(KIF2s)是中性或碱性的,其大小为479~506 个氨基酸。KIFs由低硫多肽亚基组成,低硫多肽含有高浓度的赖氨酸、天冬氨酸、谷氨酸和亮氨酸,这种组成有利于-螺旋形成。虽然KIFs 含硫较低,但末端结构域含有大量的半胱氨酸残基,通过二硫键与KAPs 交联形成结构蛋白,结构蛋白的二级结构为4 个-螺旋和非螺旋结构相连,含有大量二硫键。Yu 等已在绵羊中鉴定出17 个KIFs,这些KIFs 分别归入上述的2 种KIFs 中,KIF1s 包括10 个,分别是KRT31、KRT32、KRT33A、KRT33B、KRT34、KRT35、KRT36、KRT37、KRT38、KRT39、KRT40,KIF2s 包括7 个,分别是KRT81、KRT82、KRT83、KRT85、KRT86 和KRT87。
2.2 KAPs KAPs 由10~30 个蛋白质组成,第一类为高甘氨酸-酪氨酸角蛋白(High-Glycine-Tyrosine KAPs,HGT-KAPs),甘氨酸和酪氨酸含量为35~60 mol%,主要由基因家族和基因编码;第二类为高硫角蛋白关联蛋白(High-Sulfur KAPs,HS-KAPs),半胱氨酸含量为小于或等于30 mol%,主要由和基因家族、和等基因编码;第三类为超高硫角蛋白关联蛋白(Ultra-High Sulfur KAPs,UHSKAPs),半胱氨酸含量大于30 mol%,主要由和等基因家族编码。到目前为止,已在绵羊中鉴定出35 个KAPs,这些KAPs 被归入以下17 个家族中(表1)。

表1 羊已鉴定的KRTs 和KAPs 家族成员及特征
KAP1 家族在绵羊中已鉴定出KAP1.1、KAP1.2、KAP1.3 和KAP1.4。该家族蛋白氨基酸序列高度保守。KAP2 家族属于HS-KAPs,目前包括BIIIA3A和BIIIA3,该家族蛋白含有24 mol% 的半胱氨酸残基和半胱氨酸五聚体重复结构(CCXPX)。KAP3家族属于HS-KAPs,存在3 个主要蛋白(KAP3.1A、KAP3.2、KAP3.3)和1 个次要蛋白(KAP3.1B)。KAP4 家族属于UHS-KAPs,该家族蛋白含有一个半胱氨酸五聚体重复结构。KAP5 家族属于UHS-KAPs,发现了和,该家族蛋白富含半胱氨酸、丝氨酸或甘氨酸的重复结构。KAP6 家族属于HGT-KAPs,鉴定出KAP6.1、KAP6.2、KAP6.3、KAP6.4 和KAP6.5,该家族蛋白由甘氨酸-酪氨酸和甘氨酸-酪氨酸-甘氨酸的重复单元组成。KAP7 家族属于HGT-KAPs,只鉴定出了KAP7.1。甘氨酸-酪氨酸重复结构的数量较少。KAP8 家族属于HGT-KAPs,包括KAP8.1 和KAP8.2。KAP8.2含有很高的天冬氨酸和谷氨酸。KAP11 家族是由KAP11.1 单个成 员组成 的HS-KAPs。KAP13 家 族属于HS-KAPs,只鉴定出了KAP13.3,含有较高的丝氨酸、苏氨酸、精氨酸和甘氨酸,但不含丙氨酸,许多残基可能会磷酸化,为碱性蛋白。KAP20 家族属于HGT-KAPs,鉴定出绵羊KAP20.1和山羊KAP20.2。绵羊KAP20.1 多肽甘氨酸和酪氨酸含量较高。山羊KAP20.2 多肽甘氨酸含量较高。KAP21 家族属于HGT-KAPs,绵羊在该家族中鉴定出KAP21.1 和KAP21.2,该蛋白含有较高的甘氨酸和酪氨酸。KAP22 家族属于HGT-KAPs,只鉴定出了KAP22.1,该蛋白甘氨酸和酪氨酸含量大约一半以上,甘氨酸-酪氨酸和甘氨酸-酪氨酸-甘氨酸的重复次数较少。KAP24 家族可能属于HS-KAPs,成员为KAP24.1。丝氨酸和酪氨酸含量高,二聚体甘氨酸-酪氨酸重复序列在该蛋白中较少。KAP26 家族属于HS-KAPs,目前只报道了KAP26.1,该蛋白有非常高的丝氨酸含量。KAP28 家族属于HS-KAPs,只鉴定出了KAP28.1,丝氨酸、苏氨酸和酪氨酸含量较高。KAP36 家族属于HGT-KAPs,只鉴定出了KAP36.1,该蛋白甘氨酸和酪氨酸含量高,不包含任何半胱氨酸。
3 与羊毛性状相关的重要信号分子
在胚胎发育期间,毛囊的形态发生依赖于真皮和表皮间一系列信号的调控,诱导2 个细胞群体的有序增殖和分化,最终形成毛干、根鞘和毛乳头。Wnt 家族、成纤维细胞生长因子家族(Fibroblast Growth Factors,FGFs)、骨形态发生蛋白家族(Bone Morphogenetic Proteins,BMPs)等信号分子可能与羊毛调控有关,控制羊毛的生长。
3.1 Wnt 家族 Wnt 家族是一类脂质修饰的分泌型糖蛋白。Wnt 信号主要分为3 类:第一类是Wnt/-连环蛋白(-catenin)信号通路,第二类是Wnt/ 平面细胞极性通路,第三类是Wnt/Ca信号通路。其中经典信号通路是Wnt/-catenin 信号通路,包括Wnt 配体与跨膜蛋白Frizzled(Fz 或Fzd)受体及其辅助受体低密度脂蛋白相关蛋白LRP6 结合、-catenin 信号在胞质聚集并转位入细胞核内激活转录因子复合物淋巴增强因子1(Lef1)/T 细胞因子3(Tcf3)以及下游靶基因的表达3 个部分构成。Wnt/-catenin 信号通路中,-catenin是一种胞内糖蛋白,当Wnt 信号被激活,Wnt 配体与Fzd 受体及其辅助受体LRP6 形成的复合物结合,这种结合导致支架蛋白Dishevelled(Dvl)的募集,进而使-catenin 的破坏复合物失活,该复合物由酪蛋白激酶1(CK-1)和糖原合成激酶3(GSK-3)等组成,因此,GSK-3功能抑制和-catenin 活性增加,-catenin 释放到细胞质,结合Lef1/Tcf,从而调控Wnt 下游靶基因转录,如和等,促进细胞增殖分化。反之,当Wnt 经典通路关闭时,GSK-3水平增加将促进-catenin 降解。
DKK1 通过负调控胰岛素样生长因子结合蛋白5(IGFBP5),在羊毛弯曲形成过程中起作用。Wnt在毛囊形态发生和发育过程中起主要调节作用。Wnt10b/DKK1 可以调节毛囊大小和间距。Wnt10b也可以在毛囊周期性生长中起促进作用。Kandyba等发现Wnt7b 是毛囊周期的重要调节因子,是BMP通路的直接靶点,Wnt7b 缺失导致毛囊激活延迟。马森研究发现绒山羊Wnt/-catenin 信号通路与绒毛周期性再生有关,使绒毛快速生长。Wang 等过表达-catenin 转基因绵羊进行研究发现,-catenin 通过激活通路的下游基因,增强和的表达,使毛囊密度和毛重增加。
3.2 FGFs FGFs 及其受体构成了一个复杂的信号转导系统,参与毛囊的生长发育过程,是毛发生长的抑制剂。绵羊由3 个外显子和2 个内含子组成,F是mRNA 不含外显子2 和含有部分外显子3 的剪接变异体。研究表明,FGF5s 对绒山羊毛乳头细胞FGF5 的活性具有抑制作用。Li 等发现FGF5s 通过拮抗FGF5 功能诱导毛发生长,增加羊毛的长度和重量。Zhang 等将成纤维细胞生长因子5()基因敲除后,羊毛和毛囊密度显著增加。柳楠等发现mRNA 在腹部的表达量明显高于颈部,可能与毛囊发育负调控有关。Zhao 等发现FGF18 在敖汉细毛羊中表达下调,FGFR3 蛋白也下调,起抑制作用。Huh 等发现成纤维细胞生长因子20(FGF20)控制毛囊发育中真皮聚集物以及毛干的形成。
3.3 BMPs BMP 是转化生长因子(Tranforming Growth Factor-,TGF-)超家族成员之一。有研究表明,BMP 信号对毛乳头细胞(DPCs)的功能必不可少,并且也是毛发必需的复杂上皮-间充质信号的关键。转录因子Sox2 通过BMP 介导上皮-间质间的相互作用,Bmp6 表达增多抑制细胞迁移和BMP 抑制剂Sostdc1 表达减少,来调节纤维的生长。
Noggin 作为BMP2 的抑制剂,起着强抑制基因表达的作用,而且Noggin 具有提高皮肤中音猬因子(Sonic Hedgehog,SHH)表达的功能,Noggin 对BMP2 的抑制作用部分通过SHH 实现。苏蕊发现基因在脱落期次级毛囊周围高表达,抑制毛囊生长。mRNA 的表达量从生长期到过渡期逐渐升高,在过渡期达到最高峰,到退行期开始降低至生长期水平。Wu 等研究发现BMP6 抑制Wnt 信号通路,而Wnt10b 抑制BMP 信号通路,因此它们之间的平衡调节毛囊周期性生长从休止期向生长期转变。殷金凤等发现湖羊基因与羔皮性状(大花、中花和小花)有关,同一时期基因在大花、中花中的表达量高于小花。
4 羊毛性状相关基因及变异
羊毛性状的差异很大程度上来自于遗传,还受到营养、性别、年龄、种群管理和生理状况(怀孕和哺乳)等非遗传因素影响。虽然通常认为羊毛的生产性状受多个基因的影响,但也可能有单基因对羊毛影响较大。王杰等发现存在主基因效应。已有研究证明和变异表达对羊毛性状具有重要影响。
4.1和基因以及基因变异 Mclare 等通过连锁分析将和定位到绵羊的1号染色体上,定位到绵羊的11 号染色体上,家族和家族位于11 号和3 号染色体。目前鉴定的基因位于1号染色体上(图4)。和基因家族位于11 号染色体(图4)。基因家族位于21号染色体(图3)。

图3 羊KRT 和KAP 基因在染色体上的位置示意图(如5.4 代表KAP5.4,K33 代表KRT33)[14]
和的基因变异与不同羊毛性状也有密切关系(表2)。研究表明,基因与羊毛亮度、长度和平均纤维直径等有关。晏华春等对基因进行研究发现,g.52C>T 处发生突变,羊毛纤维直径差异不显著。但Sulayman 等发现基因与纤维直径有关,基因也与纤维直径有关,基因与卷曲数有关,基因与卷曲和纤维直径有关。
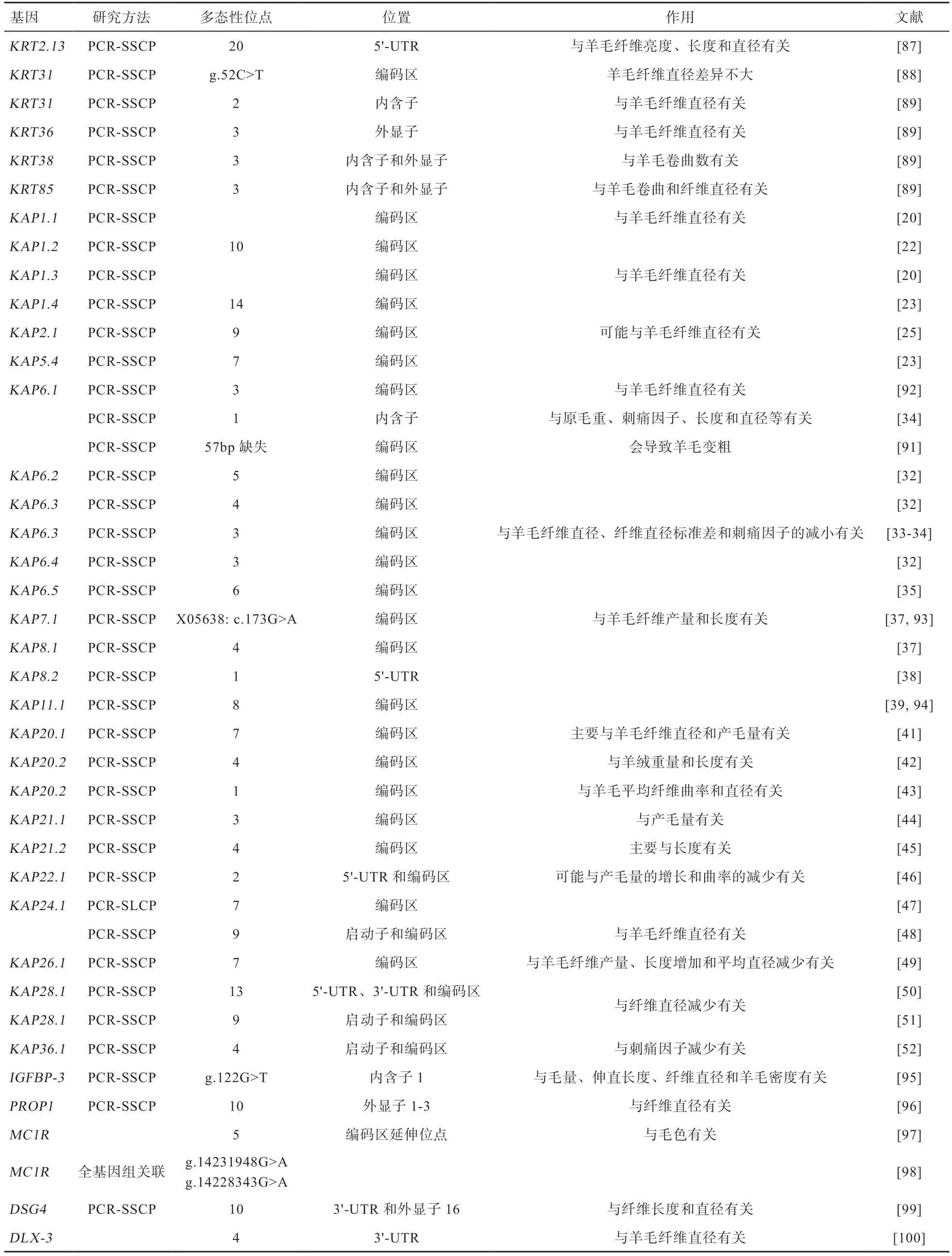
表2 羊KRT 和KAP 基因以及其他基因变异与羊毛相关性状的关联
在基因家族中,Gong 等发现绵羊基因中有14 个单核苷酸多态性(SNPs)位点;以及绵羊基因变异,共检测到6 个SNPs 和1 个长度多态性。随之在基因上,发现了10 个SNPs。Wang 等对基因而且检测到了该基因可能影响平均纤维直径。Parsons 等发现和与平均纤维直径有关。Zhou 等发现基因56 bp 缺失影响平均纤维直径。基因变异不仅会影响美利奴杂种羊和滩羊的羊毛纤维性状,而且还影响罗姆尼绵羊的羊毛纤维直径,也对Barki 绵羊的刺痛因子、纤维长度和直径等性状有影响。Li 等对基因进行研究,检测到5 种不同的核苷酸序列(A、B、C、F 和G),G 与纤维直径和刺痛因子的减少有关,而B 与这些性状增加有关。2020 年又有研究报道基因的新变异序列H,H 中非同义SNP(MN046971: c.71A>G)会导致酪氨酸转变为半胱氨酸,促进羊毛的生长。Gong 等在基因上检测到5 种DNA 序列。随后发现绵羊中也存在基因,并在TATA 盒上游21bp 处发现了SNP。Ullah 等对基因进行研究,发现与羊毛产量和长度有关。
在基因家族中,Gong 等对绵羊基因序列进行研究,发现存在1 个SNP(NM_001080740:c.334A>G)突变导致氨基酸变化。随后又有研究发现基因(登录号:HQ595347)存在3 个SNPs(c.240G>A、c.285C>G/A 和c.331A>G),但与羊毛性状关联有待进一步研究。Wang 等发现陇东绒山羊基因有3 个SNPs(NM_181616: c.37C>T、c.125T>C 和c.126G>A)与羊绒的重量和长度有关。而绵羊基因影响平均纤维直径和曲率。Gong等鉴定出了基因与毛重增加、平均纤维直径、刺痛因子和产毛量降低有关。
在基因家族中,基因与羊毛产量有关。与长度最相关。Li 等对基因变异进行检测发现3 种不同的核苷酸序列(A-C),B 与羊毛产量的增加和平均纤维曲率的降低有关。Zhou 等对绵羊基因进行分析发现7个SNPs 可能影响羊毛性状。陇东绒山羊的影响羊绒纤维直径。有研究发现,基因有2个SNPs(AM941740.1: c.215G>A 和c.277A>G),与羊毛产量和长度的增加、平均纤维直径和刺痛因子的降低有关。Wang 等对陇东绒山羊的基因序列进行分析,发现了5 种不同的DNA 序列(A-E),变异A 与平均纤维直径的减小有关。有研究发现绵羊也与平均纤维直径有关。Gong 等检测到基因有3 种不同的DNA 序列(A-C),B 与羊毛中刺痛因子的增加有关。
4.2 其他基因以及基因变异 除了上述和基因外,还有其他基因也在羊毛性状中发挥着重要作用,如表2 所示。沈敏等发现胰岛素样生长因子结合蛋白3 基因()与剪毛量、伸直长度和纤维直径等有关。垂体特异性转录因子祖蛋白基因()与纤维直径显著相关。Hepp 等发现黑色素皮质激素受体1()基因与羊毛颜色有关。比如已发现滩羊基因与毛色变化有关。Ling 等对桥粒芯蛋白4 基因()进行研究,发现与羊毛长度和卷曲有关。绵羊皮肤中同源盒蛋白基因,通过miR-188 影响mRNA 表达,导致羊毛卷曲发生变化。
5 小 结
综上所述,目前关于绵羊和山羊的羊毛相关蛋白角蛋白关联蛋白(KAPs)以及基因家族的多态性及其与羊毛性状的关系研究较多,对基因家族的多态性及其与羊毛性状的相关性的研究较少。虽然许多研究都探讨了基因多态性与羊毛性状的关系,但研究复合基因型对羊毛性状的影响比较少,毛囊周期调控涉及Wnt、BMPs 等信号分子之间复杂的相互作用还没有得到充分研究,不能完全正确理解毛囊周期,所以未来应积极进行进一步羊毛、毛囊生长发育调控机制的研究,探明毛囊的分子机制,不断挖掘与羊毛性状相关的新候选基因并对已发现的基因进行深入研究,找出基因影响羊毛性状的机理,以期为今后羊毛产业与毛用羊品种的选育提供理论参考,为养羊业更好地发展做出贡献。
猜你喜欢
中国畜牧杂志(2022年1期)2022-11-06
中国畜牧杂志(2022年9期)2022-09-14
中国畜牧杂志(2022年4期)2022-04-15
情感读本·道德篇(2022年2期)2022-04-12
儿童时代·幸福宝宝(2019年11期)2019-12-13
爱你·健康读本(2019年11期)2019-12-10
科技风(2019年15期)2019-10-21
赤峰学院学报·自然科学版(2019年9期)2019-09-10
文萃报·周二版(2019年37期)2019-09-10
数学大王·低年级(2016年8期)2016-05-14
