
广灵大尾羊FITM2 基因扩增、序列及表达特性分析
2022-10-12 13:17秦朋云乔利英刘建华潘洋洋侯文欣车雨彤刘文忠
中国畜牧杂志 2022年10期
秦朋云,乔利英,刘建华,潘洋洋,侯文欣,车雨彤,赵 祥,刘文忠
(山西农业大学动物科学学院,山西太谷 030801)
羊肉营养丰富,是高蛋白低固醇的肉类之一。中国传统文化认为羊肉具有一定食疗药补功效,因此其深受消费者青睐。而脂肪堆积过多会降低绵羊胴体品质和肉质口感,影响肉用绵羊的经济价值。脂肪组织作为动物体内维持能量代谢平衡的主要器官,是动物体内重要的储能部位。脂肪的沉积不仅受到营养等环境因素影响,更受到诸多遗传因素的调控。因此探究脂肪代谢相关的遗传因素,对改善绵羊脂肪沉积方式具有重要作用。
脂肪储存跨膜诱导蛋白(Fat Storage-Inducing Tra nsmembrane Protein,FIT/FITM)是一个独特的蛋白质家族,其家族包含2 个基因和,二者氨基酸序列相似性为50%,都定位于细胞内质网。主要在骨骼肌和心脏中表达,而在全身各组织中普遍表达,尤其在白色和棕色脂肪组织中高表达。研究发现,这2 种蛋白都有6 个跨膜结构域,N端和C 端均位于细胞质。参与将甘油三酯打包成脂滴的过程,促进脂滴在细胞中的积累,但不参与甘油三酯的合成。在肝脏中被鉴定为过氧化物酶体增殖物激活受体-(PPAR)激动剂诱导的转录物,后证明对脂滴的形成至关重要。的缺失导致细胞脂滴数量减少,过表达导致脂滴中脂质储存增加。研究表明小鼠脂肪组织中选择性敲除会导致脂肪营养不良和代谢功能障碍。这些结果都表明FITM2 蛋白对动物脂肪沉积和生命维持具有关键作用。
前期对于的研究主要集中在人、小鼠和猪中,缺乏对该基因在反刍动物中的机制和功能的认识,且关于的报道主要集中在生化作用,对于基因转录调控方面少见研究。基于本课题组前期对广灵大尾羊和小尾寒羊脂肪组织的转录组测序分析发现,基因在不同品种绵羊的不同组织中差异表达。本实验旨在通过克隆广灵大尾羊基因CDS 区,分析序列特征及蛋白特性,并通过生物信息学软件分析基因启动子和可能与之结合的转录因子,以及它们在前体脂肪细胞分化过程中的表达规律,为进一步研究广灵大尾羊基因在脂肪沉积过程中的作用提供理论依据。
1 材料与方法
1.1 样品采集 本实验采集4 月龄广灵大尾羊尾部脂肪组织,浸泡于含1%双抗(青霉素-链霉素混合液)的无菌PBS 溶液中,置于冰盒带回实验室用于分离前体脂肪细胞。随机选取3 只8 月龄广灵大尾羊安乐死后放血,采集尾部脂肪、皮下脂肪、肾周脂肪和背最长肌组织,用锡纸包裹置于液氮带回实验室,-80℃保存,用于后续基因扩增及组织表达分析。
1.2 主要试剂 Trizol 试剂、PrimeScriptRT reagent kit反转录试剂盒、PrimeSTARHS(Premix)高保真酶、TB Green Premix Ex TaqII qPCR 定量试剂盒、限制性内切酶、基因组DNA 提取试剂均购自TaKaRa 公司;琼脂糖、DH5感受态细胞、II 型胶原酶、LB 固体/液体培养基均购自索莱宝公司;无内毒素质粒提取试剂盒、胶回收试剂盒均购自OMEGA 公司;无缝克隆试剂盒购自诺唯赞公司;双抗(青霉素-链霉素混合液,Penicillin-Streptomycin Solution)、胎牛血清和高糖DMEM 培养液购自Biological Industries 公司;过表达载体pHBLV-CMVIE-ZsGreen-T2A-puro 购自汉恒生物科技(上海)有限公司。
1.3 总RNA 提取及cDNA 合成 采用Trizol 法提取4种组织的总RNA,用核酸蛋白测定仪(NanoDrop)检测总RNA 浓度和纯度(1.8<A260/280<2.0),保存于-80℃冰箱中备用。根据PrimeScriptRT reagent kit反转录试剂盒获得cDNA,保存于-20℃。
1.4 广灵大尾羊基因的克隆 根据NCBI 中收录的绵羊基因(XM_004014584.4)序列信息,利用Primer Primier 5.0 软件设计基 因CDS 区 的特异性引物(表1),由上海生工生物工程股份有限公司合成。以脂肪组织cDNA 为模板,通过PCR 扩增出基因CDS 区全长,经2% 琼脂糖凝胶电泳后切割目的条带,并纯化回收。用I 和I 快切酶使载体线性化,按无缝克隆试剂盒说明书连接载体和目的片段。连接产物转化感受态细胞,涂板后12 h 挑选单个阳性克隆菌落过夜摇菌,并将菌液送至北京六合华大基因有限公司进行测序。

表1 引物设计
1.5 广灵大尾羊基因CDS 区生物信息学分析 通过Jalview、MEGA X 软件对广灵大尾羊基因CDS 区进行氨基酸多序列比对和遗传进化树构建;利用生物信息学软件预测基因编码蛋白的特性(表2)。
1.6 广灵大尾羊基因启动子核心区、CpG 岛及转录因子预测分析 从NCBI 中获取绵羊基因(NC_040264.1)转录起始位点上游2 000 bp 及下游100 bp 作为启动子序列。利用BDGP 预测核心启动子区;利用Methprimer 预测启动子区CpG 岛;利用JASPAR、Animal TFDB、ALGGEN 和ChEA3 软件预测与基因启动子潜在结合的转录因子(预测网站见表2)。

表2 生物信息学分析软件
1.7 广灵大尾羊前体脂肪细胞的分离和培养 采集广灵大尾羊尾部脂肪组织在75% 酒精中浸泡15 s,转移到装有PBS 缓冲液的培养皿中,用眼科镊和眼科剪分离出肉眼可见的血管。将脂肪组织剪成小块,转移到15 mL离心管,加入等体积2 mg/mL II 型胶原酶,37℃ 200 rpm条件下消化30 min,随后加入等体积完全培养基(DMEM高糖培养基中加10%胎牛血清和1%青霉素-链霉素混合液)终止消化。室温下1 000 rpm 离心10 min。挑去上层漂浮的脂肪,缓慢倒掉上清,尽可能吸干净残余液体,加入完全培养基重悬沉淀,用200 目和400 目细胞筛过滤重悬液,将细胞接种到10 cm 培养皿中,添加完全培养基至每皿10 mL。轻摇培养皿使细胞均匀分布有利于细胞贴壁,后置于37℃的5% CO的细胞培养箱中培养。6 h 后观察细胞是否贴壁,如果贴壁则更换新的完全培养基,此后每2 d 更换一次培养基,每次换液前用PBS 洗涤,至细胞汇合度达到约80%进行传代、冻存或后续实验。
1.8 实时荧光定量PCR(Real-time quantitative PCR,qPCR)
1.8.1 广灵大尾羊基因在4 种不同组织中的表达利用NCBI 中Primer-BLAST 设计基因的特异性定量引物(表1),以-为对照,qPCR 检测mRNA 在广灵大尾羊尾脂、皮下脂肪、肾周脂肪和背最长肌组织中的表达量。反应体系如下:cDNA(50 ng/μL)2 μL,上下游引物(10 μmol/L)各0.4 μL,TB Green Premix Ex Taq II 5 μL,ddHO 补足至10 μL。反应程序为:95℃5 min;95℃10 s,60℃15 s,72℃20 s,设置39 个循环;95℃5 s,65℃1 min,升温速率5℃/s,从65℃递增到97℃,反应至40℃结束。
1.8.2 广灵大尾羊基因和部分转录因子在前体脂肪细胞分化过程中的表达 待前体脂肪细胞汇合度达到80%时,更换分化诱导培养基,并记做分化第0 d,此后每2 d 更换一次培养基,分化3~4 d 后显微镜观察有可见脂滴生成。分别收集分化第0、2、4、6、8、10 天的细胞,提取总RNA 并反转录成cDNA。利用Primer-BLAST 设计基因特异性定量引物(表1),以-作对照。以各时期的细胞cDNA 为模板进行qPCR 反应,反应体系及程序同1.8.1。
1.9 数据处理及分析 qPCR 结果以-为内参基因,用2来计算mRNA 相对表达量,对所得数据进行显著性分析。<0.05 表示差异显著,<0.01 表示差异极显著。用GraphPad Prism 7.0 绘制柱状图。
2 结果
2.1 广灵大尾羊基因CDS 区的克隆 成功克隆出广灵大尾羊基因的CDS 区,以2%琼脂糖凝胶电泳检测扩增产物,结果见图1,可见一条约为900 bp的条带(包含部分非编码区),与理论扩增目的条带长度一致。
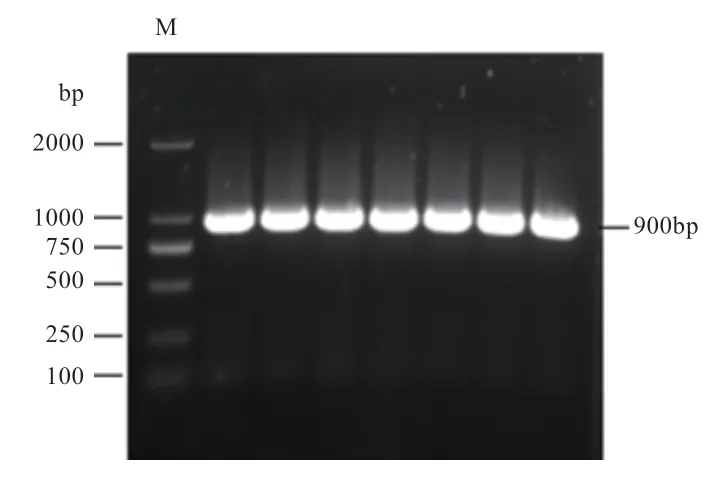
图1 广灵大尾羊FITM2 基因CDS 区扩增
2.2 广灵大尾羊基因CDS 区生物信息学分析
2.2.1 广灵大尾羊基因序列分析和蛋白理化性质分析 利用NCBI 中的ORF Finder 对广灵大尾羊基因的开放阅读框进行定位分析。结果显示基因位于13 号染色体,包含一个长852 bp 的开放阅读框。通过在线软件Protparam 预测到广灵大尾羊基因CDS 区编码蛋白质有283 个氨基酸,分子式为CHNOS,相对分子质量为32 162.64,理论等电点为9.59,该蛋白含有20 种氨基酸,含量最高的为亮氨酸(Leu),达到13.8%,含量最低的为天冬氨酸(Asp)仅为1.1%。带负电荷的残基总数(Asp+Glu):16,带正电荷的残基总数(Arg+Lys):31。预测到该蛋白的消光系数(Mcm,=280 nm)为78 880,不稳定系数为49.51,说明这种蛋白不稳定,容易降解或变性。脂溶指数为94.73,总平均亲水性(GRAVY)为0.134,表明该蛋白为疏水性蛋白。该蛋白在哺乳动物生物体外的网织红细胞中的半衰期为30 h。通过Protscale 对蛋白的疏水性进行分析(图2),其中正值表示疏水性,负值表示亲水性,值越大则性能越强,而该图中大部分位点所对应的值都大于0,表明该蛋白为疏水性蛋白。

图2 FITM2 蛋白疏水性分析
2.2.2 广灵大尾羊FITM2 蛋白的亚细胞定位及跨膜结构分析 通过PSORT II 软件进行FITM2 蛋白亚细胞定位分析时发现,该蛋白分别位于内质网、质膜、空泡、线粒体、高尔基体中,其比例分别为33.3%、33.3%、11.1%、11.1%、11.1%。通过在线软件TMpred 预测FITM2 蛋白的跨膜结构,发现FITM2 编码蛋白存在6个跨膜结构域(图3)。

图3 FITM2 蛋白跨膜结构域预测
2.2.3 广灵大尾羊FITM2 蛋白的信号肽、磷酸化预测利用SignalP 5.0 在线软件对广灵大尾羊FITM2 蛋白序列进行预测分析发现,该蛋白N 端不存在信号肽,是非分泌蛋白。磷酸化位点由NetPhos 2.0 在线软件预测,对潜在磷酸化位点的3 种氨基酸即丝氨酸(Ser)、苏氨酸(Thr)、酪氨酸(Tyr)进行预测,若分值大于0.5,则被视为潜在位点。预测结果发现有13 个丝氨酸位点、10 个苏氨酸位点、3 个酪氨酸位点超过了阈值,表明这26 个位点可能成为蛋白激酶磷酸化的位点(图4)。
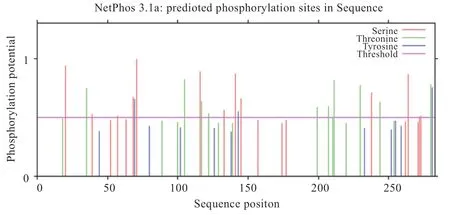
图4 FITM2 蛋白磷酸化位点预测
2.2.4 FITM2 蛋白二级结构和三级结构预测 通过SOPMA 在线软件分析预测,结果表明FITM2 蛋白的二级结构主要由-螺旋、无规则卷曲、延伸链和-转角组成。其中-螺旋占55.48%,无规则卷曲占30.04%,延伸链占11.66%,-转角占2.83%(图5)。通过I-TASSER 在线软件预测FITM2 蛋白三级结构,结果见图6。

图5 FITM2 蛋白二级结构预测

图6 FITM2 蛋白三级结构预测
2.2.5 广灵大尾羊基因序列的多序列比对及遗传进化树分析 利用Jalview、MEGA X 软件对绵羊(XP_004014633.2)、山羊(XP_005688629.2)、人(NP_001073941.1)、小鼠(NP_775573.1)、大鼠(NP_001 101269.1)、猪(NP_001121932.1)、牛(NP_001096565.1)等7 个物种的基因序列进行氨基酸多序列比对(图7)和遗传进化树分析(图8)。从多序列比对看,很多氨基酸位点保守不变,说明基因编码区在各物种中保守性较好。从遗传进化树看,绵羊与山羊亲缘关系最近,与牛、猪和人亲缘关系次之,与小鼠和大鼠的亲缘关系最远。

图7 氨基酸多序列比对结果

图8 不同物种FITM2 蛋白序列进化树分析
2.3 广灵大尾羊基因启动子生物信息学分析 选取基因转录起始位点ATG 上游2 000 bp 及下游100 bp作为基因的启动子。通过BDGP 在线软件预测到基因有3 个核心启动子区,分别位于基因上游-1 624 bp~-1 575 bp,-1 597 bp~-1 548 bp,-167 bp~-118 bp(图9A)。通 过Methprimer 软件预测出在基因上-206 bp~+39 bp 的位置上有1 个245 bp 的CpG 岛(图9B)。通过启动子在线分析软件预测发现该启动子序列包含有真核生物启动子典型的TATA-box 和CAAT-box,并预测到大量可能存在的转录因子。

图9 启动子核心区和CpG 岛预测
2.4基因启动子转录因子的预测 通过在线软件JASPAR、Animal TFDB、ALGGEN 和ChEA3 对启动子上转录因子进行预测,分别预测到633、584、312、65 个转录因子,使用Venn 在线软件取4 个软件预测结果的共同部分,得到9 个转录因子(图10A),。结合在本课题组之前脂肪组织二代测序数据库研究结果发现,表达量较高的有,并在脂肪细胞生长过程中发挥转录调节作用。部分转录因子在部分启动子上结合位点的相对位置如图10B 所示。

图10 预测出与FITM2 基因具有靶标关系的转录因子及部分结合位点
2.5 广灵大尾羊基因在4 种不同组织中的表达量 定量结果显示,基因在4 种组织中都有表达,在脂肪细胞中表达量显著高于背最长肌组织,在浅层脂肪中的表达量显著高于深层脂肪中的表达量(图11A)。如图11B 所示,基因的mRNA 表达量在分化过程中呈上升趋势,表明可能促进前体脂肪细胞分化,并促进脂滴的积累。转录因子的表达趋势与一致,可能正调控基因并促进前体脂肪细胞分化。而在第4天之后呈下降趋势,但与表达趋势不完全相反。
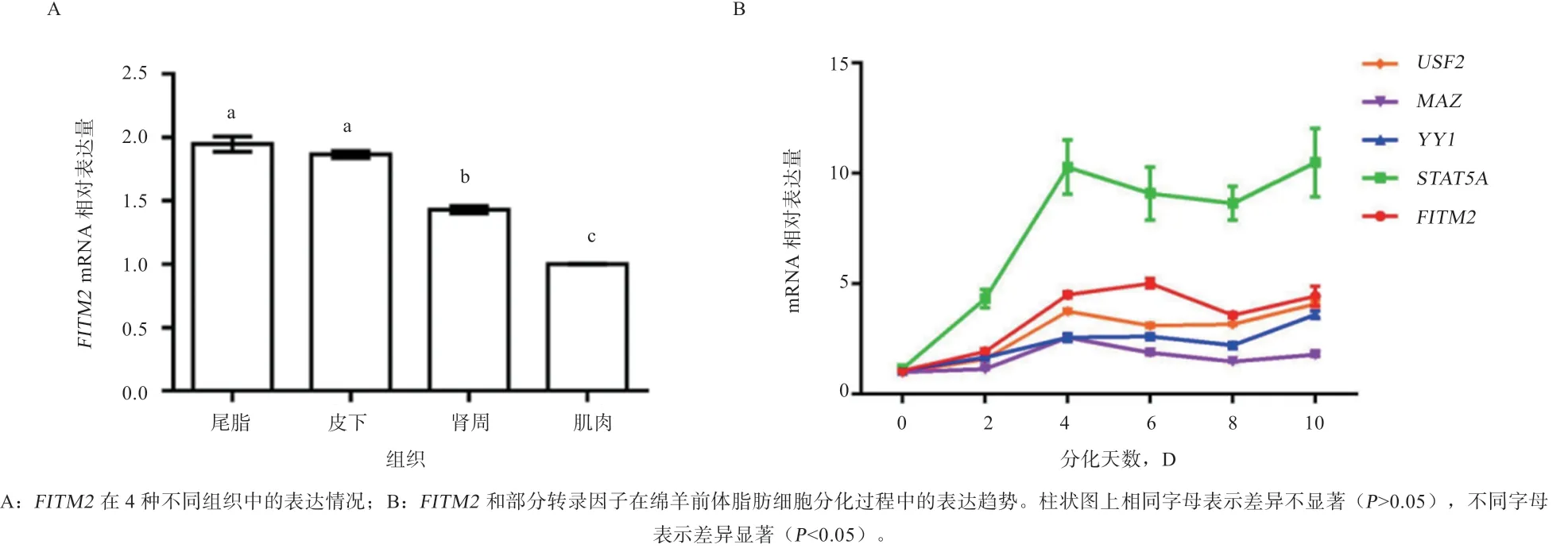
图11 实时荧光定量结果
3 讨 论
是进化上高度保守的基因,在细胞中将甘油三酯打包成脂滴的过程中发挥重要作用。本实验经过基因克隆获得广灵大尾羊基因CDS 区全长852 bp,生物信息学分析发现其编码283 个氨基酸,没有信号肽,与NCBI 上收录的绵羊基因(XM_004014584.4)的编码蛋白一致,但与猪、牛、人和小鼠的序列相比在5' 端多出21 个氨基酸。有研究表明,猪基因编码262 个氨基酸,其中N 端的30 个氨基酸为信号肽,其作用是靶向运输分泌的FITM2 蛋白到胞外。但是广灵大尾羊的FITM2 蛋白没有预测到信号肽,可能与它在N 端多出的21 个氨基酸有关。生物信息学分析广灵大尾羊FITM2 蛋白有6个跨膜结构域,二级结构多为-螺旋和无规则卷曲结构,前人研究发现FITM2 蛋白大部分结构域是由跨膜结构域组成的,而跨膜结构域在本质上是螺旋结构,与本预测结果一致。本研究预测到广灵大尾羊FITM2蛋白结构含有多个潜在的磷酸化位点,这些位点可能成为蛋白激酶磷酸化的位点。研究发现FITM2 蛋白和参与磷脂生物合成的基因之间有强相互作用,在之后的研究中发现FITM2 作为酰基辅酶A 双磷酸酶在维持内质网结构和脂质在脂滴中的正常储存发挥作用。
脂肪细胞中含有大量脂滴,脂滴是细胞内中性脂质的主要储存场所,由极性单磷脂层包裹疏水核心组成,参与细胞内物质的代谢和转运、细胞信号传导及脂类合成代谢等过程,是一个活动旺盛的多功能细胞器。研究表明,基因在哺乳动物组织中普遍表达,尤其在白色和棕色脂肪组织中表达最高。王静对猪基因进行组织表达谱分析发现mRNA在各种组织中表达量都较高。研究发现基因没有参与甘油三酯合成,只是在甘油三酯积累中发挥作用,基因主要表达于脂肪组织,其特点是产生大脂滴。本文对基因在广灵大尾羊尾部、皮下、肾周脂肪和背最长肌组织中的表达量进行分析,发现mRNA 表达量在尾脂和皮下脂肪中不存在显著差异,但和肾周相比差异显著,且在3 种脂肪组织中的含量均显著高于背最长肌组织,表明基因主要在脂肪组织中高表达,并且在绵羊前体脂肪细胞开始分化后,基因含量开始升高,表明基因可能促进绵羊前体脂肪细胞分化,与前人研究一致。
真核生物基因表达量受多个水平的调控,尤其在转录水平,其中研究最多的是关于启动子的研究,启动子是启动靶基因开始转录的DNA 序列,位于基因5'端上游序列中,可特异性识别和结合RNA 聚合酶并使之活化,从而起始转录。因此,研究启动子有助于了解基因在转录水平的调控机制。本研究预测到3 个核心启动子区,分别位于基因上游-1 624 bp~-1 575 bp、-1 597 bp~-1 548 bp、-167 bp~-118 bp,发挥主要作用的核心区域需要进一步通过构建双荧光素酶报告载体进行双荧光素酶活性分析验证。在启动子的-206 bp~+39 bp的位置上预测到1 个长245 bp 的CpG 岛,表明在此区间内可能会发生甲基化修饰进而影响基因的表达。在该启动子序列中发现真核生物启动子典型的TATA-box 和CAAT-box,并预测到大量可能存在的转录因子,做韦恩图取交集并参照本课题组之前的脂肪组织转录组二代测序数据库,筛选出和4 个在脂肪组织中含量较高的转录因子。因此本文初步研究了这4 种转录因子在前体脂肪细胞分化过程中的表达趋势,结果表明转录因子和可能正调控基因表达并促进前体脂肪细胞分化,在第4 天之后表达量下降,与表达趋势不一致且未完全相反,推测其可能不参与基因的表达调控。然而这些转录因子在成脂过程中能否通过调节基因的表达进而影响前体脂肪细胞分化仍然未知,需进一步研究。
4 结 论
广灵大尾羊基因CDS 区全长852 bp,编码283 个氨基酸。生物信息学分析表明FITM2 定位于内质网,有6 个跨膜域和26 个潜在磷酸化位点,在哺乳动物间高度保守。基因在脂肪组织中表达量高。初步筛选出STAT5A、YY1、USF2 和MAZ 4 个对基因活性可能有影响的转录因子。本实验结果为进一步探究广灵大尾羊基因在脂肪沉积过程中的作用提供理论依据。
猜你喜欢
中国农业科学(2022年16期)2022-09-19
中国农业科学(2022年15期)2022-08-09
电脑报(2020年40期)2020-11-06
赢未来(2019年15期)2019-08-14
电脑知识与技术(2018年19期)2018-11-01
恋爱婚姻家庭·养生版(2018年6期)2018-08-01
数学学习与研究(2018年7期)2018-05-16
课程教育研究·新教师教学(2017年33期)2018-05-07
山东青年(2017年11期)2018-03-29
科学中国人(2016年9期)2016-11-04
