
菊叶薯蓣DcPMK基因克隆及互作蛋白筛选
2022-10-12 06:40王宏鹏李一丹谭晓宇陈成彬张力鹏
植物研究 2022年5期
王宏鹏 李一丹 汪 耀 谭晓宇 陈成彬 张力鹏
(南开大学生命科学学院,天津 300071)
菊叶薯蓣()原产于墨西哥,是薯蓣科(Dioscoreaceae)多年生藤本植物,其根茎中重要成分薯蓣皂素具有多种生物学活性,在抗肿瘤、保护心血管和抗菌消炎等方面发挥重要作用,是合成蛋白同化激素、性激素和肾上腺皮质激素最理想的前体原料,具有较高药用价值,被誉为“药用黄金”。研究表明,菊叶薯蓣终年无休眠期和枯萎期,新块茎在老块茎上延伸或增长,并且单产和亩产产量远高于国内薯蓣品种如盾叶薯蓣()和穿龙薯蓣()等。因此,在我国制药工业所用野生薯蓣资源濒临枯竭的情况下,对菊叶薯蓣的开发和研究有利于缓解薯蓣皂素紧缺的问题。此外,近年来薯蓣属植物的研究主要集中于其有效成分薯蓣皂素提取技术改良、药理作用和栽培技术等方面,关于其皂素的生物合成途径及其调控机制研究报道较少。
薯蓣皂素属于三萜类化合物衍生而成的甾醇类化合物。在高等植物中,萜类物质主要由甲羟戊酸(MVA)途径和2--甲基--赤藻糖醇-4-磷酸(MEP)途径合成,其中MVA 途径为某些倍半萜、甾醇的合成提供了前体。磷酸甲羟戊酸激酶(phosphomevalonate kinase,PMK)是植物MVA 途径中的一个重要限速酶,催化甲羟戊酸-5-磷酸和ATP 形成甲羟戊酸-5-焦磷酸和ADP 的可逆反应,形成异戊二烯亚基用于合成甾醇、二十二醇和泛醌等各种必需化合物。目前,已在多种植物中克隆出基因,如蓖麻()、麻疯树()、木薯()、拟南芥()、橡胶树()、丹 参()和 盾 叶 薯 蓣等。王霞报道了菊叶薯蓣中基因,通过转录组测序获得基因信息,并采用生物信息学对其定位、结构和功能进行了分析。
本研究从菊叶薯蓣中成功克隆了基因,对其氨基酸序列,组织特异性和水杨酸诱导表达模式进行分析,并构建的酵母双杂交诱饵载体筛选互作蛋白,以期进一步了解菊叶薯蓣基因,为深入研究菊叶薯蓣萜类物质的合成积累提供一定理论基础。
1 材料与方法
1.1 试验材料
供试材料为菊叶薯蓣,2019 年7 月采集于云南西双版纳(2 年生)和广东湛江(4 年生),经南开大学宋文芹教授鉴定为薯蓣科药用植物菊叶薯蓣,均为墨西哥引进种。取部分2年生根茎材料经液氮速冻后,保存于-80 ℃用于转录组测序;其余2 年生和4 年生根茎材料种植于南开大学生命科学学院资源圃中生长繁殖并进行后续试验。
1.2 试验方法
总RNA 提取采用EasteprSuper 总RNA 提取试剂盒(Promega 公司,上海)进行柱式法提取。以NanoDrop1000(Gene Company,China)检测总RNA的纯度和浓度。cDNA 第一条链的合成利用M-MLV 反转录酶(TaKaRa Japan)进行,产物用无核酸酶无菌ddHO稀释10倍后,-20 ℃保存。
基因序列来源于南开大学生命科学学院已完成的转录组数据库,基于拼接的unigene 注释信息,以phosphomevalonate kinase/PMK为关键词对数据库进行检索,选取具有完整开放阅读框(ORF)的基因序列,设计特异性引物(见表1)。以2年生菊叶薯蓣叶片的cDNA为模板,PCR 反应体系包含2.0 μL cDNA,12.5 μL Prime-STAR Primix(2×)(TaKaRa Japan),各0.8 μL 上下游引物,用ddHO 补足至25.0 μL。反应程序为98 ℃预变性2 min;98 ℃变性10 s,55 ℃退火15 s,72 ℃延伸30 s,30 个循环;最后72 ℃延伸5 min,4 ℃保存。PCR 产物用1%琼脂糖凝胶电泳检测,用PCR 产物纯化试剂盒纯化后与pMD19-T载体连接,转化大肠杆菌DH5α,在氨苄抗性平板上进行筛选,经菌液PCR检测后挑选阳性克隆测序。

表1 引物名称及序列Table 1 Primer and their sequences
通过生物信息学在线网站对获得的基因氨基酸序列进行分析。利用blastx(https://blast.ncbi.nlm.nih.gov/Blastx)进行序列验证;利用ORF Finder(http://www. bioinformatics. org/sms2/orf_find.html)获 得 编 码 框;利 用blastp(https://blast.ncbi.nlm.nih.gov/Blastp)寻找同源基因;利用DNAMAN 计算与其同源基因的同源性比值;利用MEGA 7.0构建N-J进化树;利用在线软件
TMHMM sever v.2.0(http://www.cbs.dtu.dk/services/TMHMM/)和WoLF PSORT(https://wolfpsort.hgc.jp/)分析跨膜结构域和亚细胞位置;利用ExPASy(https://web.expasy.org/protparam/)分析蛋白质的理化性质;利用NetPhos3.1 Server(http://www.cbs.dtu.dk/services/NetPhos/)分析蛋白质的磷酸化位点。
组织特异性分析,以同一株2年生菊叶薯蓣的花、幼叶、老叶、幼茎、老茎、根茎和根为材料。水杨酸诱导以分别处于同一生长状态的2年生和4年生菊叶薯蓣为材料。水杨酸溶于无水乙醇后配置成不同浓度水溶液:20、50、100、200 μmol·L。以水为对照,不同处理条件都为3个平行,每3 d浇1次,每次浇1 L。40 d后取上述未经水杨酸处理的一株2年生菊叶薯蓣花、幼叶、老叶、幼茎、老茎、根茎、根7个器官,及水杨酸处理后的2年生和4年生菊叶薯蓣叶片,经无菌水洗净和液氮速冻过夜,-80 ℃超低温保存用以提取RNA,另外收集相应部分材料按蒋美红的方法进行薯蓣皂素含量检测。
利用Bio-RAD 公司的iQ5 完成实时定量PCR(qRT-PCR)。反应体系为20 μL,包含10 ng cDNA,10 μL SYBRGreen(TaKaRa Japan),上下游引物各0.8 μL,以为内参基因,每个样品包含3个生物学重复。
酵母双杂交拟南芥文库的筛选方法依Clontech 公司Matchmaker Gold Yeast Two-Hybrid System User Manual说明书进行。根据基因的全长,以及pGBKT7 载体图谱设计引物(见表1),利用无缝克隆技术连入诱饵载体pGBKT7载体,并采用LiTE/PEG 法转化Y2H Gold 菌株。经菌液PCR 验证阳性菌并进行自激活和毒性验证,以排除基因本身对杂交结果的影响。然后进行文库筛选,将诱饵载体与文库混合后涂布于SD/-Trp-Leu-His 培养基进行初筛,7 d 后挑选阳性克隆滴于SD/-Trp-Leu+X-α-Gal 培养基,随后挑选蓝斑菌落摇菌提取质粒。利用T7 和3′BD/3′AD 进行扩增和测序,根据测序结果确定互作基因。最后,将比对后的阳性克隆,阳性对照pGBKT7-53+pGADT7-T、阴性对照pGBKT7-+pGADT7菌液分别培养至OD=0.5,取6 μL 分别滴于SD/-Trp-Leu、SD/-Trp-Leu-His 和SD/-Trp-Leu+X-α-Gal 培养基,30 ℃培养3 d后拍照。
2 结果与分析
2.1 菊叶薯蓣DcPMK基因的克隆
因菊叶薯蓣缺少基因组和参考基因组信息,本研究对菊叶薯蓣进行转录组测序。通过拼接技术获得1 条长度1 536 bp 注释为的unigene 序列。利用NCBI ORF Finder 分析其含有一个完整的ORF,与NCBI 数据库中圆山药(,登录号为120259136)同源基因比对进一步验证其为完整的基因(序列一致性达91.10%)。随后提取2 年生菊叶薯蓣叶片总RNA,经NanoDrop1000检测,所提取RNA的浓度在200 ng·L之上,/在2.0~2.2,/大于1.8。以总RNA反转录成的cDNA为模板,利用特异性引物对其全长进行扩增,扩增产物与预期大小相符(见图1)。将PCR产物与pMD19-T载体连接,转化大肠杆菌DH5α,经测序得到菊叶薯蓣基因大小为1 536 bp,编码511个氨基酸,序列信息已提交到NCBI GenBank,登录号为MZ171241。
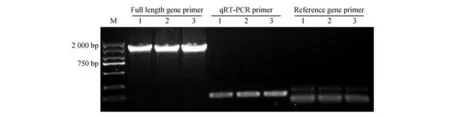
图1 DcPMK基因全长引物、定量引物和内参引物PCR扩增电泳结果Fig.1 PCR amplification electrophoresis results of gene DcPMK
2.2 菊叶薯蓣DcPMK基因生物信息学分析
ExPASy 预测结果显示,DcPMK 蛋白分子式为CHNOS,相对分子质量为54 831.32 顿,等电点为5.63,为酸性蛋白质。不稳定系数为39.35,说明DcPMK 蛋白较稳定。溶脂系数为90.7,说明其为明显的脂溶性蛋白。总平均亲水性为-0.092,说明具有一定的亲水性。WoLF PSORT和TMHMM sever v.2.0 预测结果显示,DcPMK 蛋白位于细胞质膜,含有跨膜结构域。NetPhos 3.1 Server预测结果显示,DcPMK 蛋白含有39个Ser位点、12个Thr位点和5个Tyr位点,说明可能受到蛋白磷酸激酶的调控。
将基因编码蛋白质的氨基酸序列提交到NCBI 的蛋白质序列数据库进行Blastp 搜索,结果显示,与圆山药的PMK 蛋白序列相似性最高,达88.85%。其次为海枣(,74.46%)、油棕(,73.39%)和菠萝(,72.85%)。并且发现含有一个与ATP 结合的位点Gly-X-Gly-XX-Ala。说明PMK蛋白氨基酸序列在系统进化上有高度的保守性。此外,构建的N-J 进化树(见图2),结果显示明显与菠萝、海枣和油棕等单子叶植物处于同一分支上,遗传距离较近,表明基因的进化与物种的进化关系是比较一致的。

图2 DcPMK基因的N-J进化树分支上的数值表示支长的具体数值Fig.2 N-J phylogenetic tree of DcPMKThe numerical value on the branch indicates specific values for branch length
2.3 菊叶薯蓣不同组织中薯蓣皂素含量及DcPMK基因表达模式分析
利用HPLC对菊叶薯蓣花、叶、茎、根茎和根中的薯蓣皂素含量进行测定,并与相应组织中的基因表达量进行比对分析,研究不同组织中薯蓣皂素含量及其相关基因表达量的关系。结果显示不同组织中的薯蓣皂素含量差异显著,根茎中的皂素含量高达3.07%,花和幼叶中次之,为1.70%左右,茎和根中最低,为0.40%左右。相反,基因在菊叶薯蓣不同组织中的表达水平差别不大,相比于根茎组织,在老茎中表达最高,花和幼叶中次之(见图3)。
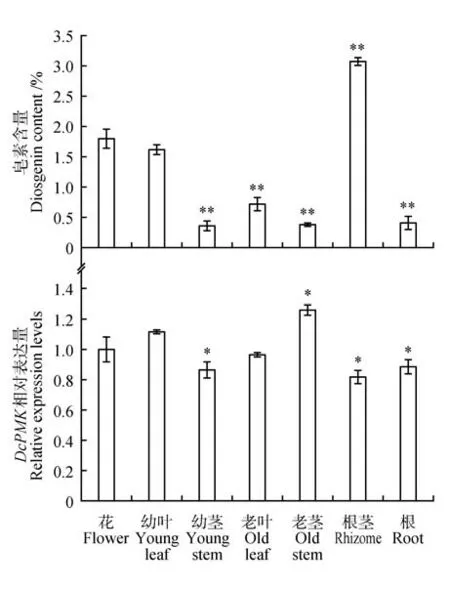
图3 菊叶薯蓣不同组织中薯蓣皂素含量变化和DcPMK表达模式分析星号代表以花器官作为对照,其他器官的显著性分析;*表示显著性差异(P<0.05),**表示极显著性差异(P<0.01)Fig.3 Accumulation of diosgenin and relative expression level of DcPMK in different tissues of D.compositeStars represented significance analyses used as other organs,with flower organs as controls;*P<0.05,**P<0.01 vs control
2.4 水杨酸诱导下薯蓣皂素含量及DcPMK 基因的表达模式分析
利用HPLC 和qRT-PCR 研究水杨酸对薯蓣皂素含量及其相关基因表达量的影响。结果如图4A 所示,不同浓度的水杨酸处理40 d 后,2年生菊叶薯蓣叶片中皂素含量均提高,空白对照中薯蓣皂素含量为1.05%,而浓度为20、50、100、200 μmol·L水杨酸处理后其叶片中皂素含量达1.30%~1.60%。相应地基因也表现出上调趋势,尤其是20 μmol·L水杨酸处理后表达量较对照组提高2.7 倍左右;此外,如图4B 所示,随着水杨酸浓度的提高,4 年生菊叶薯蓣叶片中皂素含量也明显增加,100 μmol·L水杨酸处理后其叶片中皂素含量高达1.56%,较对照组CK 提高了6 倍,与对应叶片中基因表达量的变化趋势吻合:100 μmol·L水杨酸处理后其表达量最高为对照CK的1.4倍。
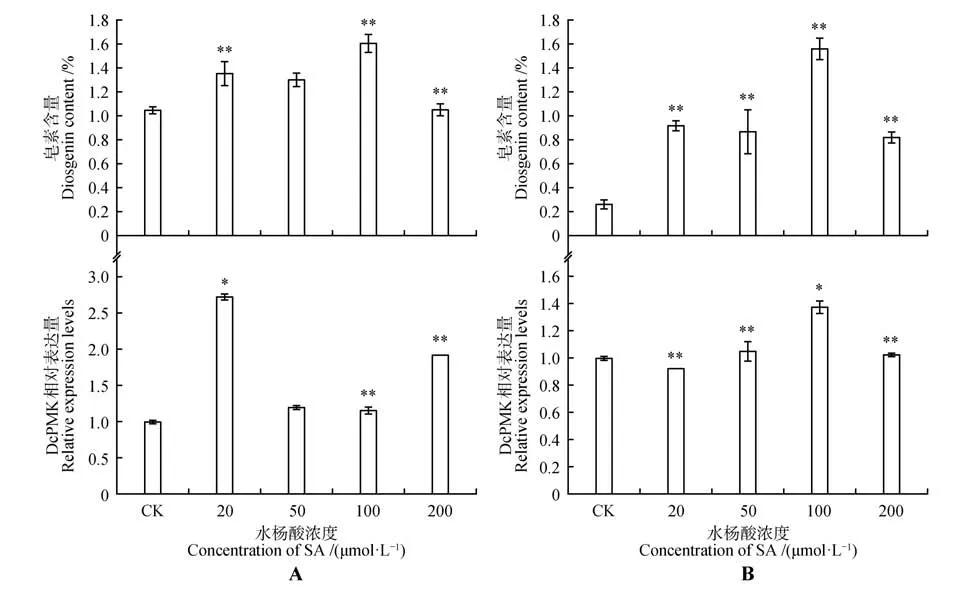
图4 水杨酸诱导下菊叶薯蓣皂素含量变化和DcPMK表达模式分析A.2年生菊叶薯蓣叶片;B.4年生菊叶薯蓣叶片;*表示显著性差异(P<0.05),**表示极显著性差异(P<0.01)Fig.4 Accumulation of diosgenin and relative expression level of DcPMK under plant hormones SA treatment conditionsA.Leaf of 2-year-old D.composite;B.Leaf of 4-year-old D.composite;*P<0.05,**P<0.01 vs control
2.5 DcPMK互作蛋白的筛选
为了进一步研究基因的功能,对基因进行克隆,并筛选拟南芥酵母文库。首先验证其自激活活性和细胞毒性(见图5),转化pGBKT7-DcPMK 的酵母细胞在SD/-Trp 培养基上能正常生长,但在SD/-Trp-His培养基上无法生长,阴性对照pGBKT7 在SD/-Trp-His 培养基上无法生长,阳性对照在SD/-Trp-His培养基上正常生长,表明DcPMK蛋白没有自主激活活性和细胞毒性。

图5 DcPMK酵母自激活和毒性验证Fig.5 Transcription activity and toxicity detection of DcPMK
将诱饵载体与文库混合后涂布于SD/-Trp-Leu-His培养基进行初筛,7 d后挑选阳性克隆滴于SD/-Trp-Leu+X-α-Gal 培养基上,培养一段时间后,共筛选出48个蓝斑菌落摇菌提取质粒,利用T7和3/BD/3/AD 分别进行PCR 反应,以验证阳性菌株(见图6),结果均能扩增出基因的条带,同时大部分拟南芥互作基因的长度在1 000 bp左右,符合拟南芥cDNA文库的基因大小,说明文库筛选有效。然后将这48 个阳性克隆测序并比对,最终筛选到了27个与互作明显的蛋白,其中出现2 次以上的互作蛋白有6 个。根据拟南芥文库数据库收录的基因功能描述可知,可能通过与这些蛋白发生相互作用,参与蓝光反应(AT2G47590)、生长发育(AT1G67730、AT3G4908 0、AT2G33793)、光合作用(AT2G33800)等生物学进程,对冷害(AT2G33800)、盐害(AT5G67500、AT5G15090)、干 旱(AT3G10910)、病 原 菌(AT4G00660)等非生物与生物胁迫环境中的植物体进行保护,并响应植物激素SA(AT5G16710)等的积累(见表2)。为了验证蛋白质之间的相互作用,滴板验证如图7 所示,27 个阳性克隆,阳性对照pGBKT7-53+pGADT7-T 均能在SD/-Trp-Leu 和SD/-Trp-Leu-His 培养基上正常生长,并在SD/-Trp-Leu+X-α-Gal 培养基上显蓝斑,阴性对照pGBKT7-+pGADT7 在SD/-Trp-Leu 培养基上正常生长,在SD/-Trp-Leu-His 培养基上无法生长,在SD/-Trp-Leu+X--gal培养基上未显蓝斑。表明与这27 个蛋白之间存在相互作用。说明可能通过蛋白质与蛋白质之间的相互作用参与调控菊叶薯蓣的生长发育、胁迫响应等代谢途径。

表2 DcPMK基因互作基因信息Table 2 The interaction genes function information of DcPMK
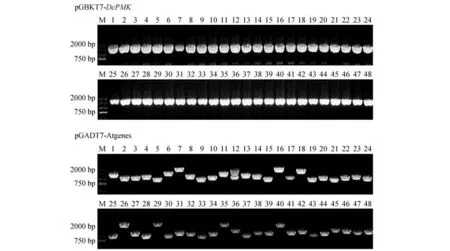
图6 DcPMK阳性酵母菌菌液PCRFig.6 PCR detection of positive yeast of DcPMK

图7 DcPMK互作蛋白验证Fig.7 Validation of interaction proteins for DcPMK
2 讨论
萜类化合物作为菊叶薯蓣重要的药效物质基础之一,具有显著的药理活性,在植物体内主要通过MVA 代谢途径合成,磷酸甲羟戊酸激酶(PMK)是植物MVA 代谢途径中的第二个关键酶,该酶在植物中的含量可以影响萜类物质的合成代谢。本研究克隆了基因,其编码蛋白位于细胞质膜,含有跨膜结构域,与李莉等报道的合成萜类物质发生在细胞质不符,但与王霞研究菊叶薯蓣中PMK 的结果相符。推测可能是菊叶薯蓣中PMK 存在特殊性。通过多序列比对发现DcPMK蛋白与其他物种相似度高达70%以上,具有PMK蛋白家族保守的1 个与催化反应密切相关的ATP结合位点Gly-X-Gly-XX-Ala。且与芦笋、菠萝和海枣等单子叶植物的PMK 蛋白处于同一分支上。其原因可能是它们均属于亚热带及热带植物,遗传距离较近。但具体原因有待进一步分析验证。
李雅静等发现盾叶薯蓣根茎中皂素含量明显高于叶片,而其皂素生物合成MVA 途径中3 个关键酶基因(、、)在叶片与根茎中表达量相当。此外,转录组分析发现有5个内生真菌仅存在于根茎的转录组中。因此李雅静等结合内生真菌参与次级代谢产物合成的相关报道,推测这可能是盾叶薯蓣根茎中内生真菌干预的结果。本研究中HPLC 结果显示菊叶薯蓣根茎中皂素含量也明显高于其他组织,且基因在菊叶薯蓣根茎、花和叶等组织中的表达量差异不明显,这与上述盾叶薯蓣中的研究结果相符。有报道称水杨酸可以有效提高菊叶薯蓣中薯蓣皂素的产量,并增强MVA 途径中角鲨烯合酶(SQS)、甲基戊二酰辅酶A还原酶(HMGR)和环阿屯醇合成酶(CAS)基因的表达。本研究发现水杨酸也能有效提高2 年生和4 年生菊叶薯蓣叶片中薯蓣皂素的产量,并上调基因的表达,说明与菊叶薯蓣萜类化合物的生物合成途径相关。此外,4 年生菊叶薯蓣于100 μmol·L的水杨酸处理后,其叶片中皂素产量和表达量最高,之后下降,说明该浓度可能是其最适浓度;2年生菊叶薯蓣叶片中皂素产量与表达量结果不太一致,推测可能是由于此时2年生菊叶薯蓣叶片中的表达量已达最高,而皂素仍处在积累阶段。
PMK 是植物MVA 途径中的关键酶之一,控制着萜类物质的合成。另外,袁聪颖等对拟南芥基因的启动子分析发现其包含有光响应、生物钟及逆境胁迫响应等元件。拟南芥文库数据库收录的基因功能描述和相关研究表明,拟南芥中,KCR1、EXPB5、RPS9M、ARA4、MED10A 和ASY4 等均能参与拟南芥的多种生理、生长及发育(如RPS9M 参与拟南芥的种子发育和胚乳发育;ASY4 突变体在减数分裂重组中显示出定量缺陷)等生物学过程。此外,VDAC2/3 编码电压依赖性阴离子通道,参与NaCl 胁迫反应和种子在低温萌发期间的呼吸调节等;RH8的过表达能够显著抑制病毒感染。PHR2 能够促进类黄酮化合物花青素积累和促进开花等。DHAR 依赖的谷胱甘肽氧化作用影响氧化还原驱动的水杨酸积累。因此,本研究推测,可能通过与上述相关蛋白互作,直接参与菊叶薯蓣的细胞伸长、衰老与生殖等生理活动,及冷害、盐害和病原菌等生物和非生物胁迫的抗性。同时,也可能通过合成的萜类化合物间接参与菊叶薯蓣的生长发育、胁迫响应等过程。
本研究成功获得了菊叶薯蓣基因序列,对其序列特征、表达模式进行了分析,结果显示能够在植物多个发育器官表达,水杨酸处理后与薯蓣皂素的积累具有正相关关系,说明是菊叶薯蓣萜类物质合成途径中关键酶基因之一。通过分析筛选出的多个拟南芥互作蛋白可以推论在菊叶薯蓣体内可能参与多种调控途径,广泛的参与其生长发育和胁迫响应等代谢途径。以上研究结果为深入了解菊叶薯蓣萜类化合物生物合成途径及其调控机制打下基础。
猜你喜欢
飞天(2022年5期)2022-05-18
学与玩(2020年11期)2020-12-23
中老年健康(2017年9期)2017-12-13
神州·下旬刊(2017年6期)2017-10-28
健康博览(2017年7期)2017-09-30
中国民族民间医药·上半月(2016年12期)2017-01-11
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
家庭科学·新健康(2014年6期)2014-08-08
恋爱婚姻家庭·养生版(2011年8期)2011-05-14
