
簕杜鹃种质资源表型性状的遗传多样性分析
2022-10-17 05:47沈甲诚焦越佳张可可何穗华金雨晴
热带作物学报 2022年9期
沈甲诚,焦越佳,张可可,何穗华,金雨晴
簕杜鹃种质资源表型性状的遗传多样性分析
沈甲诚,焦越佳,张可可,何穗华,金雨晴*
深圳市农科集团有限公司,广东深圳 518000
为深入研究簕杜鹃种质资源的遗传多样性和后期簕杜鹃种质资源的创制与利用提供参考,本研究对国内外收集的100份簕杜鹃种质资源的9个数量性状和19个质量性状进行测定与评价,利用相关性分析、主成分分析和聚类分析等方法,探讨其种质资源表型性状的遗传多样性。结果表明,在测定的19个质量性状中共检测到78个变异类型,28个表型性状的遗传多样性指数在0.61~2.06之间,9个数量性状的变异系数为12.38%~37.68%,以花序梗长度的变异系数最大(37.68%)。相关性分析结果表明性状之间关系复杂,大部分性状之间呈现显著相关性,其中叶片大小与苞片大小呈极显著正相关。通过主成分分析将簕杜鹃28个表型性状转化成9个主成分因子,累计贡献率达到70.42%,其中叶片长度、叶片宽度、苞片长度、苞片宽度、花冠直径、花被管长度、叶节间长度、花被管形状、株型等是造成簕杜鹃种质资源表型性状差异的主要因素。聚类分析可将100份簕杜鹃种质资源分为6类,其中第一类群包含7份簕杜鹃种质资源,该类群品种多为重瓣品种,具有较高的观赏价值,但后期苞片宿存,影响观赏性。第二类包含16份簕杜鹃种质资源,叶片大小中等,形状多为椭圆形,苞片形状以卵形为主。第三类包含10份簕杜鹃品种,该类群簕杜鹃品种苞片先端性状为渐尖,基部形状为心形,叶面多稍微上捧。第四类包含18份材料,该类群品种叶片中等,叶柄较短,苞片形态为阔椭圆形。第五类群包含8份簕杜鹃品种,该类群簕杜鹃品种苞片形状特异,叶柄短小,叶节间短缩,其中“塔类”系列簕杜鹃品种,整体形似宝塔,观赏价值较高。第六类包含41份材料,其中在欧式距离为17时,可将第六类群分为4个亚群,第1亚群仅包含1个品种,第2亚群簕杜鹃品种叶片多含有次色,第3亚群叶片较大,第4亚群叶片主要以卵形为主,叶面光滑无毛,苞片主要以卵形为主,花被管性状以纤细和中部收缩为主。本研究明确了不同簕杜鹃种质资源的农艺性状的特异性和遗传多样性,筛选具有特异性状的簕杜鹃种质资源,为簕杜鹃新品种的选育提供理论基础。
簕杜鹃;表型性状;遗传多样性分析
簕杜鹃(spp.)是紫茉莉科(Nyctaginaceae)叶子花属()植物[1],又名三角梅、叶子花、宝巾、九重葛等,主要分布于南美洲的巴西、秘鲁、阿根廷等国,属于热带与亚热带花卉植物。簕杜鹃全世界约有原种14~18种[2],其中光簕杜鹃()、毛簕杜鹃()、秘鲁簕杜鹃()、巴特簕杜鹃(×)、巴克簕杜鹃(×)和拉塔拉簕杜鹃(×)具有较高的园艺观赏价值[3]。
簕杜鹃在我国已有140余年的栽培历史,20世纪80年代开始,簕杜鹃因其苞片色彩艳丽,花期长等优点,广泛应用于园林绿化和盆栽观赏,并成为中国近20个城市的市花[4]。近年来,我国在簕杜鹃种质资源的收集、保存及品种选育等方面取得了一定的进展。随着簕杜鹃的广泛栽培和应用,国内簕杜鹃存在严重的品种命名不统一、不规范、植物学归属不清楚等现象,同时也存在着品种分类等级混乱、品种资源总体遗传变异情况不明等问题[3],这不仅制约了我国簕杜鹃品种的国际交流,也严重影响着我国簕杜鹃品种的推广与保护,因此加强簕杜鹃种质资源的收集保存并开展簕杜鹃种质资源遗传多样性的评价是十分必要的。表型性状包含质量性状和数量性状,基于表型性状的分析是研究植物种质资源遗传多样性最有效和最基础的方法,也是植物优良品种选育的基础。了解和掌握种质资源的多样性水平是种质资源鉴定、分类、新品种选育及应用方面突破的关键[5]。目前,基于表型性状的遗传多样性研究,已在月季[6]、荷花[7]、睡莲[8]、百合[9]等观赏植物中广泛应用。近年来,簕杜鹃属植物的研究主要集中在引种栽培、繁殖方式、观赏性评价、花期调控等方面[10-14],关于其种质资源遗传多样性的研究却鲜有报道[15],分析簕杜鹃属种质资源表型遗传多样性,不仅有利于簕杜鹃属种质资源的保存,而且对观赏性较强的簕杜鹃属植物新品种选育及优异种质的挖掘具有重要的指导意义。因此,本研究以100份簕杜鹃属种质资源为研究材料,对其28个表型性状进行观测统计,并基于表型性状进行遗传多样性分析,旨在揭示簕杜鹃属种质资源表型变异规律,为开展簕杜鹃属植物品种资源收集、分类、种质资源利用及品种改良等提供材料和理论依据。
1 材料与方法
1.1 材料
供试的100份簕杜鹃种质资源均由深圳市农科集团有限公司收集并保存于深圳市农科集团有限公司簕杜鹃种质资源圃内。具体信息见表1。
1.2 方法
试验于2021年5—8月进行观察和测量。结合簕杜鹃观赏特点并参考《植物新品种特异性、一致性、稳定性测试指南叶子花属LY/T 3206—2020》,以苞片、叶片、枝条为主要性状选取方向,筛选出28个主要性状指标(表2),其中9个数量性状利用直尺或游标卡尺进行测量,19个质量性状采用目测法并进行赋值。每份簕杜鹃种质资源观测3株,每株重复3次。

表1 100份簕杜鹃种质材料

续表1 100份簕杜鹃种质材料
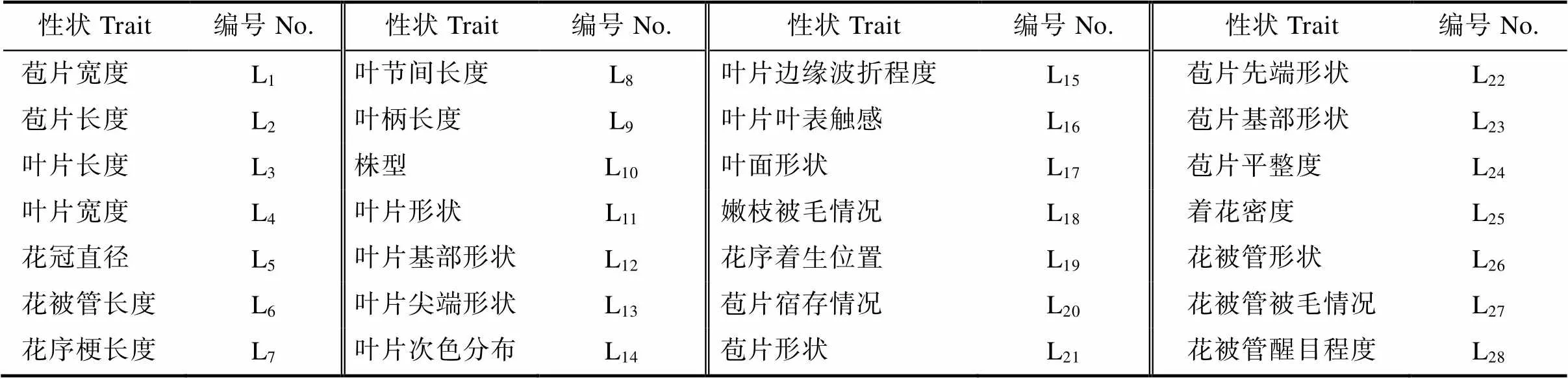
表2 簕杜鹃28个表型性状
1.3 数据处理


式中,为自然对数,P为某性状第级别包含材料份数占总材料数的百分比。
运用IBM SPSS Statistics 19.0软件进行相关性分析、主成分分析和聚类分析,在主成分分析中根据特征值大于1的原则提取主成分个数。
2 结果与分析
2.1 簕杜鹃种质资源的变异及表型多样性分析
参试的100份簕杜鹃种质资源中,9个数量性状的变异系数在12.38%~37.68%之间(表3),遗传多样性指数(′)在1.98~2.06之间。其中,花序梗长度的变异系数最大,为37.68%,花冠直径的变异系数最小,为12.38%。花序梗长度的遗传多样性指数(′)最大,为2.06,叶片长度和花被管长度的遗传多样性指数(′)最小,均为1.98(表3)。由此可见,参试的100份簕杜鹃品种具有比较丰富的遗传多样性。
通过进一步对19个质量性状的频率和遗传多样性指数(′)进行统计,结果表明19个质量性状的遗传多样性指数的变化范围介于0.61~ 1.47之间(表4),其中叶片形状的′最高,为1.47,嫩枝被毛情况的′最低,仅为0.61。在19个质量性状中有8个性状的遗传多样性指数均> 1.00,说明簕杜鹃种质资源质量性状表现出较高的遗传多样性。

表3 100份簕杜鹃种质资源数量性状表型多样性分析
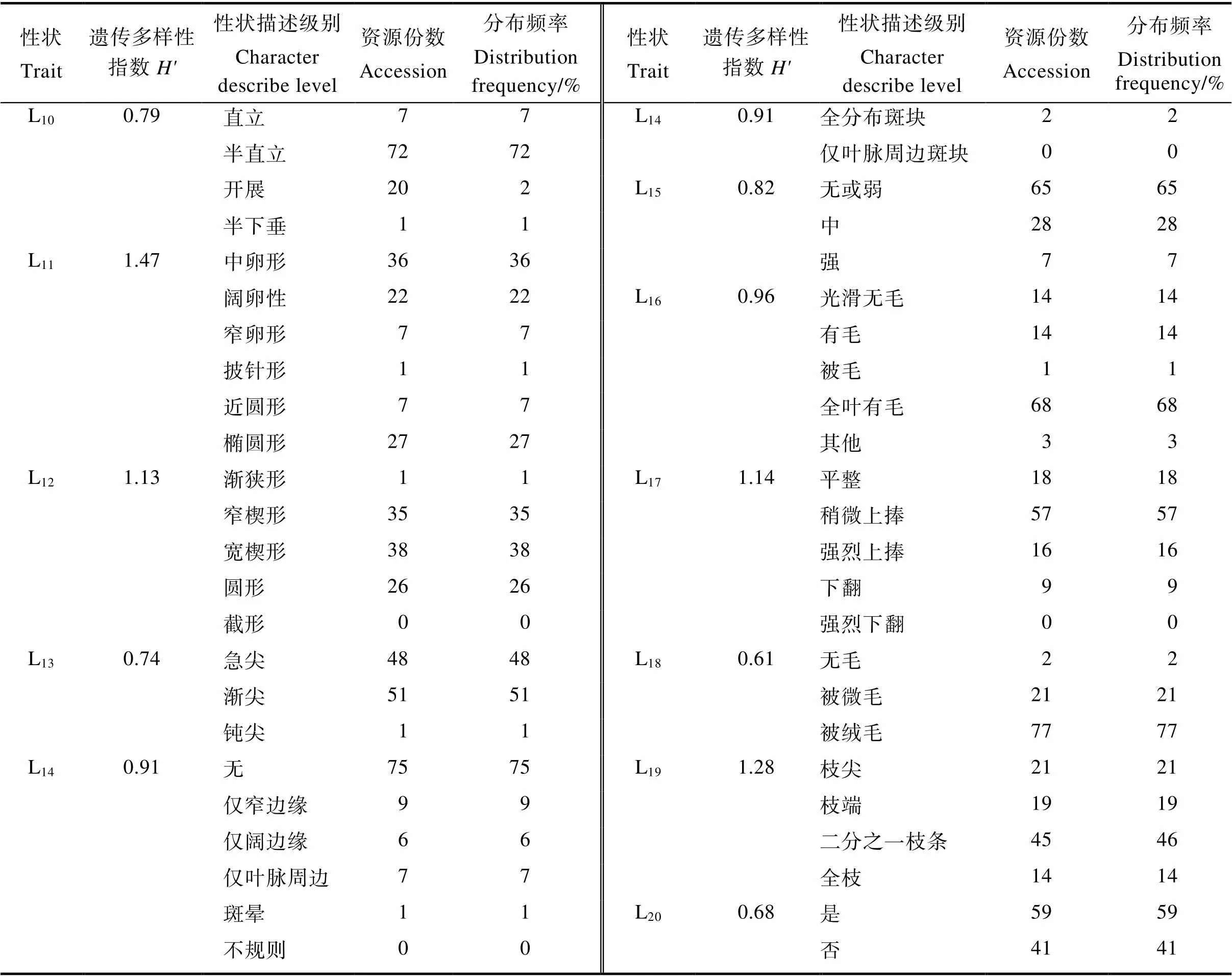
表4 簕杜鹃种质资源质量性状多样性分析

续表4 簕杜鹃种质资源质量性状多样性分析
2.2 簕杜鹃种质资源性状的相关性分析
通过对100份簕杜鹃种质资源的28个主要表型性状进行相关性分析发现,簕杜鹃种质资源的大部分表型性状之间存在显著相关性(表5)。基于花类性状和叶类性状对簕杜鹃种质资源的表型性状分别进行分析。叶类性状有11个,其中L3、L4与L8互为极显著正相关;L4、L8与L9互为极显著正相关;L3与L9互为显著正相关;L3与L11互为显著正相关。花类相关性状有15个,其中L1、L2、L5、L6和L7互为极显著正相关;L23与L2、L1呈极显著正相关;L25与L2、L1呈极显著负相关。在叶类性状与花类相关性状中,L3、L4、L2与L1存在极显著正相关,表明簕杜鹃的叶片大小与苞片大小存在密切联系。
2.3 表型性状的主成分分析
利用SPSS 19.0软件对簕杜鹃种质资源的28个主要表型性状进行主成分分析,基于特征值大于1的原则,筛选出前9个主成分,累计贡献率为70.42%,包含了28个表型性状的部分信息,表明这9个主成分可代表28个表型性状的基本特征(表6)。
第1主成分的贡献率为19.94%,特征值为5.58,该成分中影响较大的是L2、L1、L3、L4、L5、L6和L7等,其特征向量均在0.60以上,主要是与叶类性状大小和花类性状大小相关的数量性状。第2主成分的贡献率为13.33%,特征值为3.73,该成分中影响较大的是L11、L12、L16、L20、L22、L24和L25。第3主成分的贡献率为7.64%,特征值为2.14,该成分中影响较大的是L15、L26和L27等。第4主成分的贡献率为6.59%,特征值为1.85,该成分中影响较大的是L10、L19和L28。第5主成分的贡献率为5.31%,特征值为1.48,该成分中影响较大的是叶片次色分布和叶面。其余4个主成分的贡献率分别为5.10%、4.34%、4.21%和3.95%,特征值分别为1.42、1.22、1.18和1.11。
2.4 簕杜鹃种质资源表型性状的聚类分析
利用Q型聚类分析对100份簕杜鹃种质资源的28个表型性状进行聚类分析,在欧氏距离为20时,可将100份材料分为6大类(图1),不同类群簕杜鹃种质在形态表现上具有一定差异。
第一类群包含7份种质资源,该类群品种多为重瓣品种,具有苞片宿存,无真花,苞片较小等特点,此外重瓣品种具有较高的观赏价值。第二类群包含16份簕杜鹃种质资源,该类群簕杜鹃叶片形状主要为椭圆形,苞片形状多为卵形且上部外翻。第三类群包含10份簕杜鹃种质资源,该类群簕杜鹃株型主要为半直立习性,叶片形状主要为椭圆形,叶片基部性状为窄楔形,叶面多光滑无毛,苞片形状多为卵形,真花较大,其花冠直径均值达到8.23 mm。第四类群包含18份簕杜鹃种质资源,该类群簕杜鹃苞片形状多为阔椭圆形,苞片较大。第五类群包含8份簕杜鹃种质资源,该类群簕杜鹃品种苞片形状特异,叶柄短小,叶节间短缩,其中“塔紫(小精灵)”、“塔橙(卡苏米)”和“塔黄(莎莎娜)”3个品种,叶片密生于枝节,呈宝塔状,花苞密集着生于枝顶上,形似一个瓶刷,极具观赏价值。‘橙灯笼’和‘粉灯笼’2个品种苞片形状特异,形似灯笼。第六类群包含41份簕杜鹃种质资源,在欧式距离为17时,可将第六类群分为4个亚群。其中第一亚群仅含有1份簕杜鹃种质资源。第二亚群包含8份材料,该亚群簕杜鹃叶片多含有次色,其中‘金心橙白’‘金心水红’和‘金心橙’3个品种,成熟叶主脉周边有金黄色斑块。第三亚群包含6个品种,该亚群簕杜鹃品种叶片较大,叶片长和宽的均值分别达到8.03 cm和5.57 cm。第四亚群含有26个品种,该亚群簕杜鹃品种,叶片主要以卵形为主,叶面光滑无毛,苞片主要以卵形为主,花被管性状以纤细和中部收缩为主。

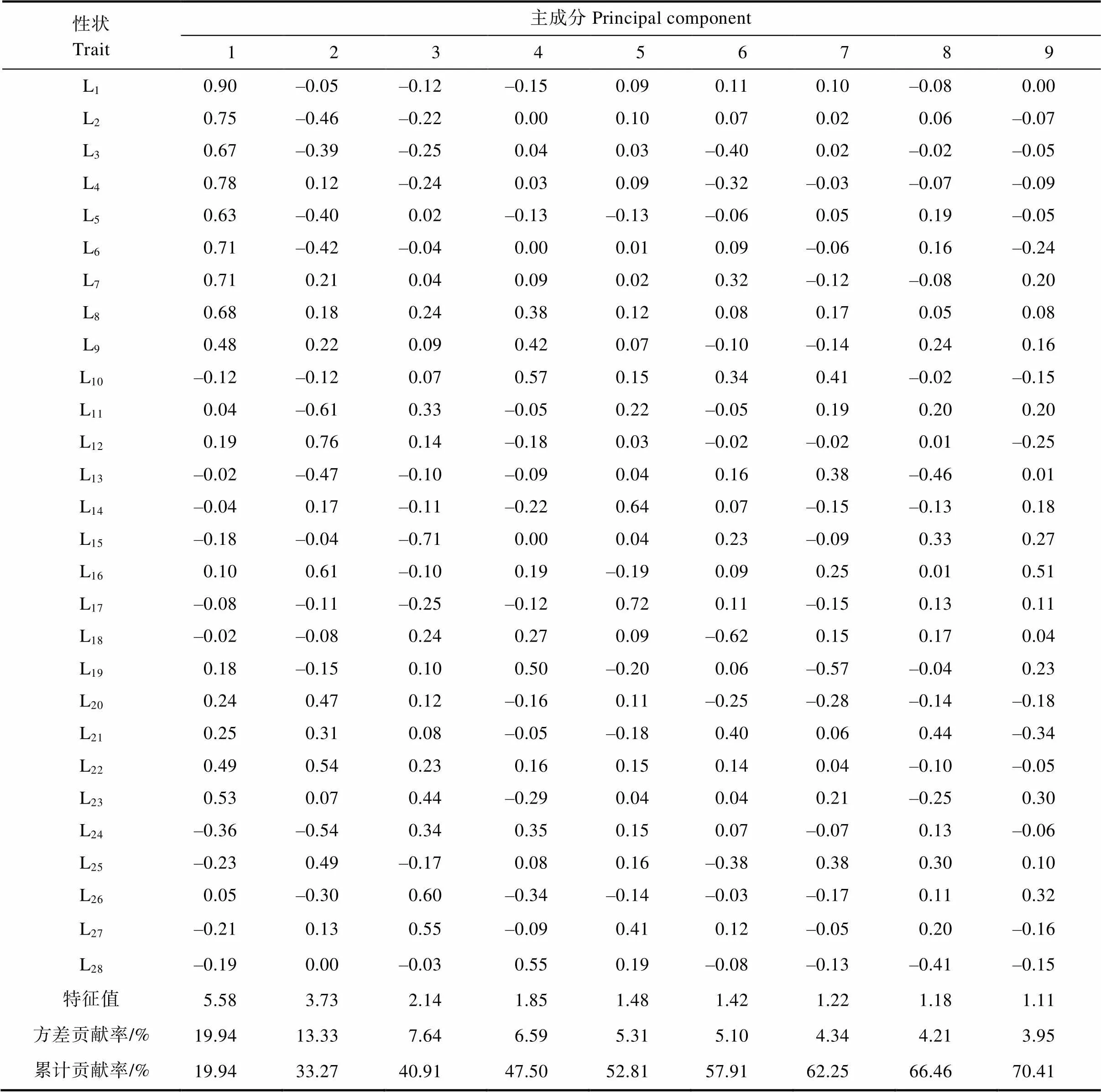
表6 100份簕杜鹃种质资源表型性状主成分分析
轴的数字为表1中对应的品种序号。
Numbers are the serial number of the corresponding varieties in Tab.1.
图1 100份簕杜鹃种质资源聚类分析
Fig.1 Cluster analysis of 100germplasm resources
3 讨论
表型性状作为植物遗传多样性研究的考量标准,其变异系数和多样性指数反映了种质间性状的离散程度和多样性,对植物表型性状遗传多样性进行分析,是合理利用植物种质资源和创制新种质的前提和基础[16]。本研究对100份簕杜鹃种质资源的28个表型性状进行遗传多样性分析,结果表明28个表型性状的遗传多样性指数在0.61~2.06之间,9个数量性状的变异系数在12.38%~37.68%之间,表明参试的簕杜鹃种质资源表型变异丰富,类型广泛,这可为簕杜鹃种质创新、新品种选育提供丰富的种质材料,同时也有利于筛选和利用优异种质资源。
主成分分析是利用降维的方法,根据需要选择较少的指标或因子来反映原来的指标信息。前人研究结果表明,利用主成分分析法可以简化表型性状的分类工作[17]。本研究利用主成分分析将100份簕杜鹃种质资源的28个表型性状转化成9个主成分因子,其累计贡献率达到70.42%,可以解释不同簕杜鹃种质资源主要表型性状的绝大部分信息,这与杨珺等[15]的研究结果类似。主成分分析可以将簕杜鹃种质资源复杂的表型性状转化成几个主要的综合指标,转化后的综合指标的使用将提高簕杜鹃种质资源的选育效率。此外,相关性分析结果表明,簕杜鹃种质资源大部分表型性状之间均存在不同程度的相关性,因此今后选育符合不同育种目标的簕杜鹃新品种需综合考虑各性状之间的联系。
聚类分析可将相似的对象聚为一类,各个类群具有不同的形态学特征[18]。本研究利用组内连接法将100份簕杜鹃种质资源划分为6类,初步明确了100份簕杜鹃种质资源的大致类型,可根据育种目标提高育种效率。其中第一类群簕杜鹃多为重瓣品种,具有较高的观赏价值,但后期苞片宿存,影响观赏价值。第5类群簕杜鹃品种苞片形状特异,叶柄短小,叶节间短缩,其中“塔类”系列簕杜鹃品种,整体形似宝塔,观赏价值较高。第六类群包含41份簕杜鹃种质资源,在欧式距离为17时,可将第六类群分为4个亚群。其中第二亚群包含8份材料,该亚群簕杜鹃叶片多含有次色。通过聚类分析结果能充分体现每个种质资源间的遗传变异和亲缘关系,能够为培育不同利用价值的簕杜鹃新品种奠定基础。
熟悉和掌握簕杜鹃种质资源主要表型性状的遗传多样性和亲缘关系,是挖掘和利用簕杜鹃种质资源的基础和前提。本研究通过利用相关性分析、主成分分析和聚类分析等方法,完成了对100份簕杜鹃种质资源的28个表型性状的综合分析,本研究结果将为后期分子标注技术与簕杜鹃表型性状评价相结合,创造优异种质资源,挖掘优异基因提供科学的理论基础。
[1] 麦有专, 刘明虎, 徐诗涛, 温艺超. 不同花色三角梅的嫁接技术[J]. 热带林业, 2006(4): 33-34.
MAI Y Z, LIUM H, XU S T, WEN Y C. Grafting techniques ofwith different colors[J]. Troplcal Forestry, 2006(4):33-34.(in Chinese)
[2] 傅立国. 中国高等植物[M]. 青岛: 青岛出版社, 2003(6): 564-576.
FU L G. Chinese higher plants[M]. Qingdao: Qingdao Publishing House, 2003(6): 564-576. (in Chinese)
[3] 周 群, 黄克福, 丁印龙, 郭惠珠. 中国引栽三角梅属观赏品种的调查与分类鉴定[J]. 江西农业学报, 2011, 23(5): 53-56.
ZHOU Q, HUANG K F, DING Y L, GUO H Z. Investigation and taxonomic identification on introduced ornamental varieties inin China[J]. Acta Agriculture Jiangxi, 2011, 23(5): 53-56. (in Chinese)
[4] 常圣鑫, 杨光穗, 陈金花, 黄素荣. 世界三角梅产业发展历史及趋势[J]. 热带农业科学, 2018, 38(1): 71-77.
CHANG S X, YANG G S, CHEN J H, HUANG S R. Development history and tendency ofindustry all over the world[J]. Chinese Journal of Tropical Agriculture, 2018, 38(1): 71-77. (in Chinese)
[5] STEPANSKY A, KOVALSKI I. ,PERL-TREVES R.. Intraspecific classification of melons (L.) in view of their phenotypic and molecular variation[J]. Plant Systematics and Evolution, 1999(217): 313-332.
[6] 吉乃喆, 华 莹, 赵世伟, 崔荣峰, 周 燕. 藤本月季表型分析与综合评价[J/OL]. 分子植物育种: 1-28[2022- 08-03].http://kns.cnki.net/kcms/detail/46.1068.S.20210517.1612.024.html.
JI N Z, HUA Y, ZHAO S W, CUI R F, ZHOU Y. Phenotype character diversity analysis and comprehensive evaluation of climbing rosearieties[J/OL]. Molecular Plant Breeding: 1-28[2022-08-03].http://kns.cnki.net/kcms/detail/46.1068.S.20210517.1612.024.html. (in Chinese)
[7] 原 鑫, 李文玲, 刘召强, 吴芳芳, 刘艺平, 孔德政. 荷花品种表型性状遗传多样性分析[J]. 江苏农业科学, 2020, 48(16): 188-193.
YUAN X, LI W L, LIU Z Q, WU F F, LIU Y P, KONG D Z. Analysis of genetic diversity of phenotypic traits of lotus cultivars[J]. Jiangsu Agricultural Sciences, 2020, 48(16): 188-193. (in Chinese)
[8] 潘庆龙, 付瑛格, 谷 佳, 盛玉辉, 李清雪, 饶 英, 朱天龙, 周 扬, 史佑海, 赵 莹, 王 健. 海南引种睡莲表型多样性分析及评价[J]. 热带作物学报, 2021, 42(10): 2777-2788.
PAN Q L, FU Y G, GU J, SHENG Y H, LI Q X, RAO Y, ZHU T L, ZHOU Y, SHI Y H, ZHAO Y, WANG J. Analysis of phenotypic diversity ofL. in Hainan province[J]. Chinese Journal of Tropical Crops, 2021, 42(10): 2777-2788. (in Chinese).
[9] 郭方其, 吕 萍, 吴 超, 彭 娟, 秦德辉, 朱文静, 黎 侠, 丁晓瑜. 浙江主栽盆栽百合种质资源表型性状遗传多样性分析[J]. 分子植物育种, 2020, 18(14): 4802-4811.
GUO F Q, LYU P, WU C, PENG J, QIN D H, ZHU W J, LI X, DING X Y. Genetic diversity of main strawberry germplasm resources in Zhejiang province bytraits[J]. Molecular Plant Breeding, 2020, 18(14): 4802-4811. (in Chinese)
[10] 孙利娜, 林 茂, 李进华, 唐 庆, 龙定建, 廖美兰. 广西30份三角梅种质资源观赏价值综合评价[J]. 农业研究与应用, 2016(6): 1-6.
SUN L N, LIN M, LI J H, TANG Q, LONG D J, LIAO M L. Comprehensive evaluation on ornamental value of 30 species ofin Guangxi[J]. Agricultural Research and Application, 2016(6): 1-6. (in Chinese)
[11] 李 瑜, 杨柳慧, 吴 宪, 马洪洋, 冯欣欣. 三角梅繁殖技术与园林应用研究进展[J]. 安徽农业科学, 2018, 46(36): 10-12.
LI Y, YANG L H, WU X, MA H X, FENG X X. Research progress on propagation and landscape application ofspp[J]. Journal of Anhui Agricultural Sciences. 2018, 46(36): 10-12. (in Chinese)
[12] 赵 阳. 三角梅的园林应用及控花养护措施[J]. 科技创新与应用, 2019(35): 128-129.
ZHAO Y.garden application and flower control and maintenance measures[J]. Technology Innovation and Application. 2019(35): 128-129. (in Chinese)
[13] 韩洪波, 吴 英. 基于三角梅外观的种质资源评价[J]. 黑龙江农业科学, 2019(12): 97-101.
HAN H B, WU Y. Evaluation of germplasm resources based on the appearance of bougainvillea[J]. Heilongjiang Agricultural Sciences, 2019(12): 97-101. (in Chinese)
[14] 李旺南. 三角梅落地栽植花期控制技术[J]. 福建热作科技, 2012, 37(3): 46-48.
LI W N. Flowering period control technology forplanting[J]. Fujian Thermal Technology, 2012, 37(3): 46-48. (in Chinese)
[15] 杨 珺, 符瑞侃, 张 珂, 陈 宣, 任军方, 云 勇. 三角梅种质资源形态学多样性和聚类分析研究[C]. 中国观赏园艺研究进展, 2016: 91-94.
YANG J, FU R K, ZHANG K, CHEN X, REN J F, YUN Y. Study on the morphological diversity and cluster analysis ofgermplasm resources[C]. Advances in Ornamental Horticulture of China, 2016(4): 82-85. (in Chinese)
[16] 火 艳, 招雪晴, 黄厚毅, 黄贤斌, 许云方, 祝遵凌, 苑兆和. 观赏石榴表型遗传多样性分析[J]. 浙江农林大学学报, 2020, 37(5): 939-949.
HUO Y, ZHAO X Q, HUANG H Y, HUANG X B, XU Y F, ZHU Z L, YUAN Z H. Phenotypic genetic diversity of ornamental pomegranate cultivars[J]. Journal of Zhejiang A & F University, 2020, 37(5): 939-949. (in Chinese)
[17] 吕 伟, 韩俊梅, 任果香, 文 飞, 王若鹏, 刘文萍. 山西芝麻种质资源SSR遗传多样性及群体结构分析[J]. 核农学报, 2021, 35(7): 1495-1506.
LYU W, HAN J M, REN G X, WEN F, WANG R P, LIU W P. Genetic diversity analysis of sesame germplasm resources in Shanxi with SSR Markers[J]. Journal of Nuclear Agricultural Sciences. 2021, 35(7): 1495-1506. (in Chinese)
[18] 彭 枫, 赵孟良, 徐晨曦, 王小丽, 葛晨辉, 王全华, 蔡晓锋. 基于表型性状的菠菜种质资源遗传多样性分析[J]. 分子植物育种, 2021, 19(5): 1698-1708.
PENG F, ZHAO M L, XU C X, WANG X L, GE C H, WANG Q H, CAI X F. Genetic diversity analysis of spinach germplasm resources based on phenotypic traits[J]. Molecular Plant Breeding, 2021, 19(5): 1698-1708. (in Chinese)
Analysis of Genetic Diversity of Phenotypic Traits ofGermplasm Resources
SHEN Jiacheng, JIAO Yuejia, ZHANG Keke, HE Suihua, JIN Yuqing*
Shenzhen Nongke Group Co., Ltd., Shenzhen, Guandong 518000, China
In order to provide a reference for the in-depth study of the genetic diversity and the creation and utilization ofgermplasm resources in the later period, this study analyzed 9 quantitative characters and 19 qualitative characters of 100germplasm resources collected at home and abroad. The determination and evaluation were carried out, and the genetic diversity of the phenotypic traits was discussed by means of correlation analysis, principal component analysis and cluster analysis. The results showed that a total of 78 types of variation were detected in the 19 qualitative traits, the genetic diversity index of 28 phenotypic traits was between 0.61–2.06, and the coefficient of variation of 9 quantitative traits was 12.38%–37.68%. The coefficient of variation of peduncle length was the largest (37.68%). The results of correlation analysis showed that the relationship between the traits was complex, and most of the traits showed significant correlation, among which the size of leaf and bract traits had a very significant positive correlation. Through principal component analysis, 28 phenotypic traits of Rhododendronwere transformed into 9 principal component factors, and the cumulative contribution rate reached 70.42%, includingleaf length, leaf width, bract length, bract width, corolla diameter, perianth tube length, leaf internode length, perianth tube shape, plant type, are the main factors that cause the differences in phenotypic traits of Rhododendron bougainvillea germplasm resources. Cluster analysis could divide the 100 germplasm resources into 6 categories, of which the first group contained 7germplasm resources. Most of the varieties in this group were double-lobed, which have high ornamental value, but the later bracts are of high ornamental value. The second category contained 16 germplasm resources. The leaves were medium in size, mostly oval in shape, and the bracts were mainly oval in shape. The third category included 10 germplasm resources. The bracts of thegroup were acuminate at the apex, heart-shaped at the base, and the leaves were mostly held up slightly. The fourth group contained 18 materials. The leaves of this group were medium, the petiole was short, and the bracts were broad and oval. The fifth group included 8 germplasm resources. The group had a specific shape of bracts, short petioles, and shortened internodes. Among them, the ‘Ta-like’ series of germplasm resources had an overall shape like a pagoda and were of high ornamental value. The sixth group contained 41 materials. When the Euclidean distance was 17, the sixth group could be divided into 4 subgroups. The first subgroup contained only one germplasm resource, and the second subgroup contained mostly secondary color leaves of Rhododendron species. The leaves of the third subgroup were larger, the leaves of the fourth subgroup were mainly ovate, the leaf surface was smooth and glabrous, the bracts were mainly oval, and the tectonic tubes were slender and centrally contracted. The leaves of the group were medium, the tips of the leaves were mostly acuminate and acute, and the bracts were mainly oval. This study clarified the specificity and genetic diversity of agronomic traits of differentgermplasm resources, and selectedgermplasm resources with specific traits to provide a theoretical basis for the selection and breeding of newvarieties.
; phenotypic trait; genetic diversity analysis
S685.99
A
10.3969/j.issn.1000-2561.2022.09.008
2021-12-30;
2022-03-07
深圳市城市管理和综合执法局科研项目(No. 202016)。
沈甲诚(1993—),男,硕士,研究方向:遗传育种。*通信作者(Corresponding author):金雨晴(JIN Yuqing),E-mail:yuqingjin29@163.com。
猜你喜欢
中华医学图书情报杂志(2022年1期)2022-11-18
中国免疫学杂志(2022年17期)2022-11-15
中国现代医生(2022年21期)2022-08-22
中国医药科学(2022年5期)2022-05-05
农村科学实验(2022年2期)2022-03-12
科学导报(2020年25期)2020-04-28
三农资讯半月报(2020年2期)2020-03-09
绿色科技(2019年14期)2019-11-19
江苏农业科学(2019年5期)2019-09-02
植物保护(2019年2期)2019-07-23
