
Neuroprotective effects of insulin-like growth factor-2 in 6-hydroxydopamine-induced cellular and mouse models of Parkinson’s disease
2022-11-11 10:28HaiYingZhangYongChengJiangJunRuiLiJiaNanYanXinJueWangJiaBingShenKaiFuKeXiaoSuGu
中国神经再生研究(英文版) 2023年5期
Hai-Ying Zhang , Yong-Cheng Jiang , Jun-Rui Li, Jia-Nan Yan Xin-Jue Wang Jia-Bing Shen Kai-Fu Ke , Xiao-Su Gu
Abstract Skin-derived precursor Schwann cells have been reported to play a protective role in the central nervous system.The neuroprotective effects of skin-derived precursor Schwann cells may be attributable to the release of growth factors that nourish host cells.In this study, we first established a cellular model of Parkinson’s disease using 6-hydroxydopamine.When SH-SY5Y cells were pretreated with conditioned medium from skin-derived precursor Schwann cells, their activity was greatly increased.The addition of insulin-like growth factor-2 neutralizing antibody markedly attenuated the neuroprotective effects of skin-derived precursor Schwann cells.We also found that insulin-like growth factor-2 levels in the peripheral blood were greatly increased in patients with Parkinson’s disease and in a mouse model of Parkinson’s disease.Next, we pretreated cell models of Parkinson’s disease with insulin-like growth factor-2 and administered insulin-like growth factor-2 intranasally to a mouse model of Parkinson’s disease induced by 6-hydroxydopamine and found that the level of tyrosine hydroxylase, a marker of dopamine neurons, was markedly restored, α-synuclein aggregation decreased, and insulin-like growth factor-2 receptor downregulation was alleviated.Finally, in vitro experiments showed that insulin-like growth factor-2 activated the phosphatidylinositol 3 kinase (PI3K)/AKT pathway.These findings suggest that the neuroprotective effects of skin-derived precursor Schwann cells on the central nervous system were achieved through insulinlike growth factor-2, and that insulin-like growth factor-2 may play a neuroprotective role through the insulin-like growth factor-2 receptor/PI3K/AKT pathway.Therefore, insulin-like growth factor-2 may be an useful target for Parkinson’s disease treatment.
Key Words: 6-hydroxydopamine; alpha-synuclein; insulin-like growth factor-2 receptor; insulin-like growth factor-2; neurodegeneration; neuroprotection;Parkinson’s disease; skin-derived precursor Schwann cells
Introduction
Parkinson’s disease (PD) is the most common movement disorder and the second most common age-associated neurodegenerative disorder after Alzheimer’s disease (Bloem et al., 2021; Tolosa et al., 2021).Patients with PD suffer from loss of neuromelanin-containing dopaminergic neurons in the substantia nigra pars compacta (Acharya and Kim, 2021).Once 50–80% of these neurons disappear, the patient’s physical activities of daily living are significantly affected.In recent years, PD research has mainly focused on the mechanism of action of the main component of Lewy bodies, α-synuclein(α-syn) (Wong and Krainc, 2017; Mahul-Mellier et al., 2020; Zheng et al.,2021).The oligomers or aggregates that form when α-syn is overexpressed have toxic effects on cells and can cause mitochondrial dysfunction (Braak et al., 2003).However, the specific pathogenesis of PD is still unclear.
Skin-derived precursor (SKP) cells are a pluripotent source of adult stem cells that are capable of self-renewal and, when cultured appropriately, can differentiate into SKP Schwann cells (SKP-SCs) (Toma et al., 2005; Hunt et al.,2009; Liu et al., 2013).Previous studies have shown that SKP-SCs derived from rat skin precursor cells reduce 6-hydroxydopamine (6-OHDA)-induced dopaminergic neuron damage (Chen et al., 2020; Ma et al., 2021; Yan et al.,2022).However, there is little evidence that SKP-SCs develop into functional neurons or form connections with neurons.The improved neuronal function seen in the presence of SKP-SCs is probably due to growth factors that they release, providing nourishment to host cells.In previous studies, we performed proteomic analysis of SKP-SCs conditioned medium (SKP-SCs-CM)and evaluated the literature to identify possible key growth factors secreted by SKP-SCs, including insulin-like growth factor (IGF)-2 (Chen et al., 2020; Ma et al., 2021; Yan et al., 2022); however, the specific regulatory mechanism by which IGF-2 promotes neuronal growth has not yet been fully elucidated.
IGFs such as IGF-1 and IGF-2, similar to insulin, are evolutionarily conserved peptides (Maki, 2010) that promote bone and muscle growth and differentiation (Agrogiannis et al., 2014).One study showed that IGF signal transduction is critical to neural development (O’Kusky and Ye, 2012).IGF-2 is widely expressed throughout the brain (Beletskiy et al., 2021).It has been found to be neuroprotective in Huntington’s disease (García-Huerta et al.,2020), Alzheimer’s disease (Mellott et al., 2014), amyotrophic lateral sclerosis(Allodi et al., 2016), and neuropsychiatric diseases (Steinmetz et al., 2018).However, there have been few studies regarding the role of IGF-2 in PD.IGF-2 is a soluble factor that exerts its effects by binding to its receptor, IGF-2 receptor (IGF-2R), which appears to be essential for growth, development,and lysosomal enzyme transport (El-Shewy and Luttrell, 2009).IGF-2R may also participate in the activation or removal of extracellular ligands, theactivation of specific transduction proteins, the formation of lysosomes, and the separation and transport of lysosomal enzymes (O’Kusky and Ye, 2012).Studies have shown that, in Huntington’s disease, IGF-2 reduces the load of polyglutamine79 in cells by binding to IGF-2R, thereby exerting a protective effect (García-Huerta et al., 2020).
Based on the evidence described above, the purposes of this study were to verify whether IGF-2 is a key component of neuroprotective effect of SKP-SCs-CM and to further investigate the neuroprotective effect of IGF-2 and related pathways in PD.
Methods
Cell culture and drug treatments
Human neuroblastoma cells (SH-SY5Y, CRL-2266, RRID: CVCL_0019, American Type Culture Collection, Manassas, VA, USA) expressing tyrosine hydroxylase(TH) were used in thein vitro
experiments in this study.The cells were cultured in Dulbecco’s modified Eagle medium/F12 (Hyclone, Logan, UT, USA)containing 10% fetal bovine serum, 1% penicillin, and 1% streptomycin in a 5%COatmosphere.To promote differentiation, SH-SY5Y cells were cultured with 10 μM all-trans-retinoic acid (Sigma-Aldrich, St.Louis, MO, USA) for 7 days,with medium changes every other day (Cheung et al., 2009; Ma et al., 2021).Cells were pretreated for 12 hours with IGF-2 (Peprotech, Rocky Hill, NJ, USA)or SKP-SCs conditioned medium (SKP-SCs-CM), then exposed for 24 hours to 6-OHDA (Sigma-Aldrich).In the IGF-2 inhibition experiment, 0.1 μg/μL of anti-IGF-2 antibody (rabbit, Cat# A2086, RRID: AB_2764106, ABclonal, Wuhan,China) was added to the SKP-SCs-CM, which was then pre-incubated at 37°C for 30 minutes (Jeon et al., 2017; Son et al., 2019).A flowchart of the study design is shown in Figure 1.
Figure 1|Flow chart of in vitro experiments.
Preparation of conditioned media
SKP-SCs isolated from the back skin of newborn green fluorescent protein–transgenic Sprague-Dawley rats were supplied by Nantong University (Wang et al., 2016; Zhu et al., 2018; Chen et al., 2020; Ma et al., 2021; Yan et al.,2022).The cells were cultured in Dulbecco’s Modified Eagle Medium/F12(3:1, Gibco, Billings, MT, USA) containing 1% fetal bovine serum (Gibco), 2%N2 (Gibco), 5 μM forskolin (Gibco), 50 ng/mL heregulin-1β (Gibco), and 1%penicillin/streptomycin (Beyotime, Shanghai, China).High-purity cultures were obtained by serial passage.Cells at passages 18–20 were used in this study.
SKP-SCs were cultured at a concentration of 4 × 10cells/cmfor 48 hours,after which the conditioned medium was collected by centrifugation at low speed (4000 ×g
) for 10–15 minutes and filtered using a 0.22-μm syringe filter.The filtered SKP-SCs-CM was then concentrated by ultrafiltration (Amicon Ultra-15, Millipore, Billerica, MA, USA) and stored at –80°C until use.Animals
All experimental procedures were approved by the Animal Care and Use Committee of Nantong University (approval No.S20200420-802) on April 20, 2020, and were performed in accordance with Guide for the Care and Use of Laboratory Animals (8edition).C57BL/6J mice (specific pathogenfree, 25–30 g, 10–12 weeks, adult, male) were reared under the following conditions: room temperature 22–24°C, relative humidity 60–65%, and photoperiod 12/12 hours light/darkness.Standard cages were used, and each cage contained no more than five mice.The mice were allowed to adapt to the environment for at least 1 week before the experiments were performed.After acclimatization, the mice were randomly allocated to the following groups: sham, IGF-2 + sham, 6-OHDA, and IGF-2 + 6-OHDA.There were eight mice in each experimental group.
6-OHDA-induced lesions
The 6-OHDA-induced mouse model of PD was established as previously described (Heuer et al., 2012; Ma et al., 2021).The mice were treated with desipramine (25 mg/kg intraperitoneal, Sigma-Aldrich) 30 minutes before injection with 6-OHDA (Sigma-Aldrich) or vehicle to protect noradrenergic neurons.After anesthetization via intraperitoneal injection of 1.25%tribromoethanol (Aibei Biotechnology, Nanjing, China), the mice were restrained on a stereotaxic apparatus (Rayward Life Technologies, Shenzhen,China).6-OHDA (8.0 μg/2 μL, dissolved in NaCl at 0.9%, supplemented with 0.02% ascorbic acid) was injected into the right striatum with a syringe pump at a rate of 0.5 μL/min in the following locations (relative to the bregma)(Heuer et al., 2012; Ma et al., 2021): anterior-posterior = +0.5 mm, mediallateral = –2.0 mm, and dorsal-ventral = –3.0 mm.Control mice were injected with 2 μL of a 0.02% ascorbic acid solution.
IGF-2 administration
Starting from 3 days before surgery, mice in the IGF-2 + 6-OHDA group received IGF-2 250 ng/d (Novoprotein, Rocky Hill, NJ, USA) for 10 consecutive days.The specific methods of intranasal administration and drug concentrations used were described previously (Hanson et al., 2013; Ouchi et al., 2013; Mao et al., 2016; Yang et al., 2020).Briefly, the mouse’s head was immobilized with one hand and the mouse was placed in the supine position;then, a 2.5-μL pipette was used to introduce the IGF-2 solution alternately into both nostrils with the other hand.IGF-2 was dissolved in phosphatebuffered saline (PBS), and the total volume (25 μL) was delivered in 2.5-μL increments, alternating nostrils, every 5 minutes.
Apomorphine-induced rotation test
This test is commonly used to evaluate the progression of 6-OHDA-induced injury (Iancu et al., 2005; Gu et al., 2016).Briefly, apomorphine (Sigma-Aldrich,0.5 mg/kg body weight) was injected into the neck subcutaneously 28 days after injury.As soon as the mouse has been injected, it is placed in a beaker with a 18.1-cm diameter and 5-L capacity.The number and direction of 360°rotations that the mouse makes in 30 minutes is recorded.When mice rotate to the uninjured side (i.e., the left side), in most cases they use the left hind limb as the fulcrum; the body then rotates to the left, and some mice may even turn over or perform a few small circular movements.Greater than or equal to seven rotations per minute was regarded as a successfulin vivo
successful model of PD.Mouse serum and brain tissue collection
The mice were anesthetized with 1.25% tribromoethanol, after which tail vein blood was collected, and the serum was separated by centrifugation.The brain was isolated from each mouse, and midbrain tissue was harvested for western blot assay.For immunofluorescence studies, the deeply anesthetized mice were fixed in a supine position on a foam board, the chest cavity was opened (fully exposing the heart to enable the apex and the right atrial appendage to be identified), the infusion needle was inserted into the left ventricle first, cut the right atrial appendage and saline was infused slowly to clear the blood in the blood vessels.After the liver turned yellow, the infusion of normal saline was replaced with 4% paraformaldehyde.The sign of successful perfusion with 4% paraformaldehyde is that the mouse tail vibrates and tilts, and the whole body becomes rigid (Gage et al., 2012).After the end of perfusion, the whole brain was removed and placed in 4%paraformaldehyde, post-fixed for 24 hours, then placed in 20% and 30%sucrose for gradient dehydration.After dehydration, the brain tissue was embedded in optimal cutting temperature compound and frozen at –20°C for at least 20 minutes.After freezing, 8–10-μm serial sections were taken, and the tissue sections were mounted on clean slides.
Human serum collection
Participants signed informed consent forms, and the Affiliated Hospital of Nantong University approved the study (approval No.2020-K047, July 9, 2020).The samples came from 19 patients diagnosed with moderate to advanced PD and 17 healthy controls.All PD patients met the clinical PD diagnostic criteria published by MDS in 2015 (Postuma et al., 2015).Exclusion criteria included the following: 1) patients with secondary Parkinson’s syndrome with a clear etiology (such as brain tumor, history of cerebrovascular disease, infection, trauma, non-PD dementia, poisoning, and other neurodegenerative diseases); 2) patients with definite mental illness;3) patients with serious cardiovascular, cerebrovascular, liver, or kidney diseases; and 4) patients with serious diseases such as blood system diseases,systemic infectious diseases, and malignant tumors.Control subjects were healthy individuals who were matched for age and sex with the PD group and underwent routine physical examinations at the hospital (Table 1).4 mL of fasting cubital median venous blood was taken from each subject in the morning and centrifuged at 3000 ×g
for 10 minutes to separate the serum,which was stored in aliquots at –80°C for later testing.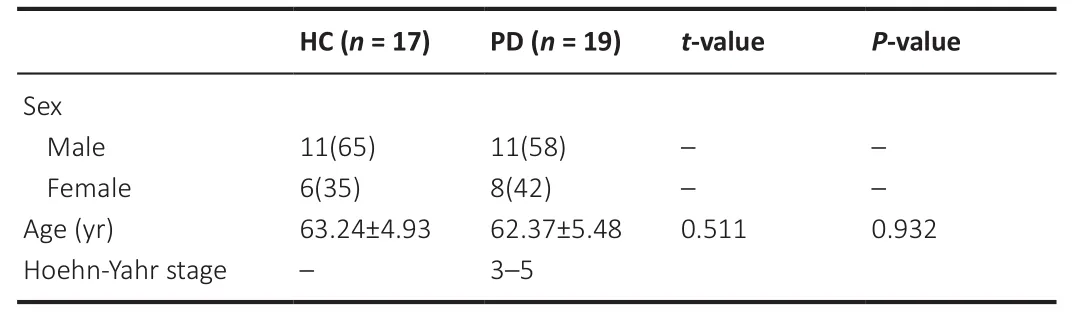
Table 1 |Comparison of general clinical data between the HC and PD groups
Enzyme-linked immunosorbent assay
To detect IGF-2 levels and explore possible changes in IGF-2 in PD, IGF-2 levels in human serum, mouse serum, and mouse midbrain tissue were determined using an enzyme-linked immunosorbent assay kit (Hengyuan, Shanghai,China) according to the manufacturer’s instructions.The absorbance at 450 nm was recorded by a microplate reader (BioTek, Covina, CA, USA).
Cell counting kit-8 assay
A cell counting kit-8 (CCK-8, Dojindo, Kumamoto, Japan) was used according to the manufacturer’s instructions to detect cell viability.First, SH-SY5Y cellswere counted and diluted to the experimental concentration (5 × 10/100 μL per well), then seeded uniformly in 96-well plates.When the cells reached 60–70% confluency, the stimulating agent(s) and/or drug(s) were added for the appropriate amount of time, after which 10 μL of CCK-8 solution was added to each well.The 96-well plate was then reincubated for 2 hours, and the optical absorbance at 450 nm was determined by a microplate reader(BioTek).
5-Ethynyl-2"-deoxyuridine assay
The 5-ethynyl-2′-deoxyuridine (EdU) assay (Beyotime) was carried out to determine cell proliferation.For this experiment, we seeded SH-SY5Y cells into a sterile, glass-bottomed 24-well plate.When the cells reached 60–70%confluency, the stimulating agent(s) and/or drug(s) were added.The 2× EdU working solution prepared in advance was preheated in an incubator and added to a 24-well plate in the same volume as the original medium, and the plates were incubated for another 2 hours.The culture medium with aspirated with a pipette, and cell fixative was added to fix the cells at room temperature.The prepared Click reaction mixture was added, and the cells were then protected from light and incubated at room temperature for 30 minutes.Nuclei were stained and cells were mounted with Hoechst 33342–containing anti-fluorescence quenching mounting solution.The cells were viewed and photographed using a fluorescence microscope (Olympus, Tokyo,Japan).The percentage of proliferating (EdU-positive) cells in the total cell population was determined after different treatments.
TdT-mediated dUTP nick-end labeling assay
We carried out this experiment strictly in accordance with the TdT-mediated dUTP nick-end labeling (TUNEL) assay apoptosis detection kit instructions(Beyotime) to assess apoptosis after various drug treatments.SH-SY5Y cells grown on glass slides were fixed with 4% paraformaldehyde at room temperature for 30 minutes, then permeabilized at room temperature for 5 minutes.Following washing with PBS, we added TUNEL detection solution to the cells, shielded the slides from light, and incubated then at 37°C for 1 hour.Finally, a fluorescence microscope was used to observe the cells.The percentage of apoptotic (TUNEL-positive) cells in the total cell population after the specified treatment was determined.
Western blot assay
We used Western blot assay to detect the protein levels of TH (a dopaminergic neuron marker), α-syn, IGF-2, IGF-2R, phosphorylated phosphatidylinositol 3 kinase (p-PI3K), and phosphorylated protein kinase B(p-AKT) after exposure of the cells to various stimuli.An appropriate amount of tissue or cell lysate solution was added to SH-SY5Y cells and mouse midbrain tissue, the supernatant was obtained by centrifugation.Next, 5×loading buffer was added to the supernatant, and the solution was boiled.The boiled solutions were aliquoted into different Eppendorf tubes to avoid sample degradation caused by repeated freezing and thawing.Samples were run on sodium dodecyl sulfate-polyacrylamide gel electrophoresis gels (stacking gel: 80 V, separating gel: 120 V) and transferred to specific membranes (0.2 or 0.45 μm, Merck Millipore, Burlington, MA, USA) (transfer conditions: 250 mA, 60–180 minutes, on ice).Next, blocking was completed by immersion in 5% nonfat dry milk for 2 hours at room temperature.Then,the membranes were incubated with primary antibody overnight at 4°C,followed by incubations with secondary antibody (1:5000, Proteintech,Wuhan, China) for 2 hours at room temperature.Immunoreactive bands were visualized using a chemiluminescence reagent (Absin, Shanghai, China) and an Odyssey Infrared Imaging System (Li-Cor Bioscience, Lincoln, NE, USA).The following primary antibodies were used: β-actin (mouse, 1:1000, Cat#ab8226, RRID: AB_306371, Abcam, Cambridge Science Park, Cambridge,UK), IGF-2 (rabbit, 1:1000, Cat# A2086, RRID: AB_2764106, ABclonal), IGF-2R (rabbit, 1:1000, Cat# A13022, RRID:AB_2759869, ABclonal), TH (rabbit,1:5000, Cat# 25859-1-AP, RRID: AB_2716568, Proteintech), α-syn (rabbit,1:1000, Cat# ab51253, RRID: AB_869973, Abcam), p-PI3K (rabbit, 1:1000,Cat# ab182651, RRID: AB_2756407, Abcam), p-AKT (rabbit, 1:1000, Cat#ab38449, RRID: AB_722678, Abcam), total PI3K (rabbit, 1:1000, Cat#ab182651, RRID: AB_2756407, Abcam), and total AKT (rabbit, 1:1000, Cat#ab8850, RRID: AB_2112117, Abcam).The following secondary antibodies were used: goat anti-mouse IgG (1:5000, Cat# SA00001-1, RRID: AB_2722565,Proteintech) and goat anti-rabbit IgG (1:5000, Cat# SA00001-2, RRID:AB_2722564, Proteintech).The membranes were visualized with an enhanced chemiluminescence system (Beyotime).Images were analyzed using ImageJ v1.6.0 (National Institutes of Health, Bethesda, MA, USA) (Schneider et al.,2012).
Immunofluorescence
Immunofluorescence was used to detect the location of proteins expressed by the target genes or co-expression of proteins expressed by two target genes.We sequentially fixed SKP-SCs with ice-cold 4% paraformaldehyde and blocked them with QuickBlockblocking buffer (Beyotime).The cells were then incubated with the following primary antibodies for 2 hours at room temperature or overnight at 4°C: S100 (rabbit, 1:200, Cat#ab52642, RRID:AB_882426,Abcam), P75 (rabbit, 1:200, Cat#ab52987, RRID: AB_881682,Abcam), glial fibrillary acidic protein (GFAP; rabbit, 1:100, Cat#3670T, RRID:AB_561049, CST, Danvers, MA, USA), IGF-2 (rabbit, 1:200, Cat# A2086, RRID:AB_2764106, ABclonal), TH (mouse, 1:50, Cat#sc-25269, RRID: AB_628422,Santa Cruz Biotechnology, Santa Cruz, CA, USA), and β3 tubulin (Tuj1; mouse,1:50, Cat#sc-80005, RRID: AB_2210816, Santa Cruz Biotechnology).Then,the cells were incubated with immunofluorescent secondary antibody(1:1000, Alexa Fluor®-488: mouse, Cat# A11029, RRID:AB_2534088; Alexa Fluor®-488: rabbit, Cat# A11034, RRID: AB_2576217; Alexa Fluor®-568:rabbit, Cat# A11036, RRID: AB_10563566, Invitrogen, Carlsbad, CA, USA) for 1 hour at 37°C or 2 hours at room temperature.From incubation with the immunofluorescent secondary antibody to final observation, all operations were performed in the dark.After final washes with PBS, a fluorescence quenching sealing tablet containing Hoechst 33342 was used to seal the slides.A fluorescence microscope (Olympus) was used to examine the cells and capture the images.For cellular immunofluorescence, five different fields were observed for each sample, and three samples were analyzed per group.For immunofluorescence analysis of brain tissue samples, three mice were randomly selected from each group, and at least five discontinuous sections were selected from each mouse for observation and analysis.
Ingenuity pathway analysis
Differentially expressed genes from substantia nigra tissues were determined from RNA sequencing (RNA-seq) data, which have been deposited in the China National Genebank DataBase (CNGBdb, https://db.cngb.org/) under Project ID accession number CNP0001036 (Yang et al., 2021).All targets were imported into Microsoft Excel (Microsoft Excel; version 2019; MicrosoftCorporation, Redmond, WA, USA) to remove duplicate values.Ingenuity pathway analysis (IPA) was employed to predict canonical pathways and networks of compounds and targets based on known interactions between different proteins.
Statistical analysis
No statistical methods were used to predetermine sample sizes; however,our sample sizes are similar to those reported in previous publications (Gu et al., 2016; Ma et al., 2021).No animals or data points were excluded from the analysis.The evaluators were blinded to the assignments.Data are presented as mean ± standard error of the mean (SEM).A two-tailed Student’st
-test was used to compare two independent groups, and one-way analysis of variance followed by Tukey’spost hoc
analysis was used to compare the means of the experimental and control groups.Clinical sample data were compared using the independent samplest
-test.Statistical analysis was conducted using GraphPad Prism 8.4.2 (GraphPad Software, San Diego, CA, USA,www.graphpad.com).Values ofP
< 0.05 were considered to be statistically significant.Results
The neuroprotective effect of SKP-SCs is related to IGF-2 release
We recently demonstrated the neuroprotective effect of SKP-SCs (Chen et al., 2020; Ma et al., 2021; Yan et al., 2022).Conditioned medium from SKPSCs can reduce the injury that 6-OHDA causes to SH-SY5Y cells, and direct transplantation of SKP-SCs can reduce 6-OHDA-induced dopaminergic neuron damagein vivo
andin vitro
by regulating autophagy.In our previous studies, we analyzed SKP-SCs-CM by mass spectrometry and identified the cytokine IGF-2 as being potentially related to the therapeutic effects of SKP-SCs.This factor is a member of the insulin polypeptide family, which is expressed at high levels in the brain and plays an instrumental role in brain development.In this study, we cultured SKP-SCs and confirmed their identity by immunofluorescence staining for the Schwann cell markers S100, P75, and GFAP (Figure 2A).Next, we explored whether IGF-2 is involved in the neuroprotection mediated by SKP-SCs-CM.An IGF-2 neutralizing antibody was added to SKP-SCs-CM, which were then pre-incubated at 37°C for 30 minutes.Both the CCK-8 and the EdU results showed that the neuroprotective effect of SKP-SCs-CM was eliminated by pre-incubation with the neutralizing antibody.As shown in Figure 2B, compared with the 6-OHDA injury group, the SKP-SCs-CM pretreatment group exhibited significantly greater cell viability (P
< 0.01).However, SKP-SCs-CM pre-incubated with the anti-IGF-2 neutralizing antibody was unable to protect SH-SY5Y cells from the damage induced by 6-OHDA.The EdU results were consistent with the CCK-8 results (Figure 2C and D).These results suggest that IGF-2 plays a crucial role in SKP-SCs-CM–mediated neuroprotection.IGF-2 levels are increased in PD serum samples
Next, we sought to obtain a more in-depth understanding of the role of IGF-2 in this context.Because IGF-2 is a soluble secretory factor, its serum level is considered to be a biomarker for disease development and prognosis (Lewittand Boyd, 2019).Therefore, we analyzed IGF-2 levels in peripheral serum samples from a mouse model of PD and patients with PD.First, we collected serum from mice in the 6-OHDA and sham groups.Statistical analysis of the enzyme-linked immunosorbent assay results showed that IGF-2 levels were higher in the serum samples of the 6-OHDA group compared with the sham group (P
< 0.01; Figure 3A).Next, we examined the level of IGF-2 in the serum of PD patients.The results were consistent with those from the animal experiments (P
< 0.01,vs
.HC group; Figure 3B).In summary, these results indicate that the level of IGF-2 in the peripheral circulation is increased in PD.IGF-2 promotes SH-SY5Y cell survival and proliferation
To determine whether IGF-2 is responsible for the observed neuroprotection,we pre-treated 6-OHDA-induced SH-SY5Y cells with recombinant IGF-2 at different concentrations (5, 25, 50, 100 ng/mL) and then measured cell viability by CCK-8 assay (Figure 4A).The results showed that IGF-2, when administered in a certain concentration range, inhibited the 6-OHDA-induced cytotoxicity.In particular, 50 ng/mL IGF-2 had a significant protective effect (P
< 0.05,vs
.6-OHDA group).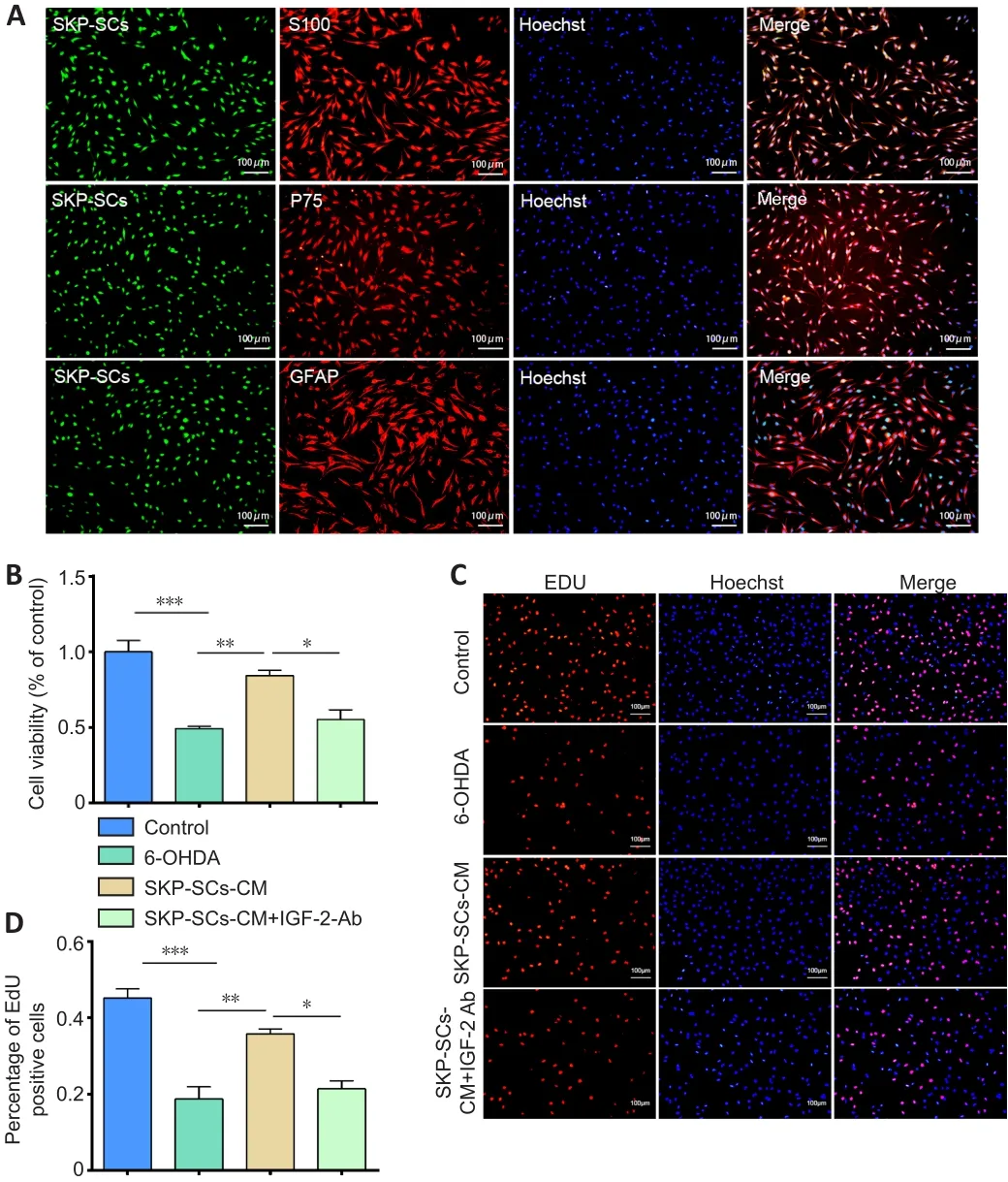
Figure 2| Pretreatment with an anti-IGF-2 neutralizing antibody reduces the neuroprotective effect of SKP-SCs-CM on 6-OHDA-treated SH-SY5Y cells.

Figure 3| IGF-2 levels are increased in serum from PD mice (A) and patients (B).
Based on the CCK-8 results described above, we chose 50 ng/mL IGF-2 as the maximum drug concentration for subsequent experiments.Next, we tested cell proliferation by EdU assay.As illustrated in Figure 4B and C, the number of proliferating cells was significantly reduced after 6-OHDA treatment (P
< 0.001,vs.
control group), but there was no significant decrease in proliferation as a result of treatment with IGF-2.We then performed a TUNEL test to further verify the ability of IGF-2 to rescue 6-OHDA-induced cytotoxicity.Compared with the 6-OHDA group, there were fewer TUNEL-positive cells in the IGF-2 +6-OHDA group (P
< 0.001; Figure 4D and E).Finally, we performed western blot analysis to determine the protein expression levels of TH and α-syn.The results showed that 6-OHDA treatment significantly decreased TH levels and increased α-syn levels (P
< 0.001,vs
.control group).Pretreatment with IGF-2 prevented the reduction in TH levels(P
< 0.01,vs
.6-OHDA group) and reduced α-syn aggregation (P
< 0.001,vs.
6-OHDA group; Figure 4F–H).In summary, IGF-2 has a protective effect in anin vitro
6-OHDA-induced model of PD.The protective effect of IGF-2 on 6-OHDA-induced injury in mice
Next, we conductedin vivo
experiments to verify ourin vitro
results.We treated C57BL/6J mice with 6-OHDA to mimic PD, administered IGF-2 intranasally, and performed an apomorphine-induced rotation experiment to evaluate the success of the model (Figure 5A).Twenty-eight days after 6-OHDA injection, the number of contralateral rotations had increased significantly (P
< 0.001,vs
.sham group), suggesting that 6-OHDA had damaged the dopaminergic system.Treatment with IGF-2 reduced the number of rotations induced by apomorphine (P
< 0.001,vs
.6-OHDA group,Figure 5B).Next, the number of THneurons in the substantia nigra was detected by immunofluorescence.In mice that had sustained injury with 6-OHDA, the number of TH+ neurons was significantly reduced (P
< 0.001,vs
.sham group).Administration of IGF-2 prevent the loss of THneurons (P
<0.05,vs
.6-OHDA group, Figure 5C and D).Finally, we analyzed TH and α-syn protein levels.Consistent with thein vitro
results, TH protein expression was significantly decreased after 6-OHDA treatment (P
< 0.01), while α-syn protein expression was significantly increased (P
< 0.001), and treatment with IGF-2 reversed these effects (Figure 5E–G).These findings suggest that IGF-2 has a neuroprotective effectin vivo
.Changes in IGF-2 and IGF-2R expression in in vitro and in vivo models of PD
To explore the specific mechanism by which IGF-2 exerts its neuroprotective effects, we first detected its localization in mouse midbrains by immunofluorescence double-label staining and showed that it was expressed in TH-positive cells, Tuj1-positive neurons, and GFAP-positive neurons (Figure 6).In addition, we measured IGF-2 protein expression levels.Bothin vivo
(Figure 7A and B) andin vitro
(Figure 7C and D) experimental results indicated that IGF-2 expression was reduced after 6-OHDA treatment (P
< 0.001,vs.
sham group;P
< 0.01,vs
.control group).Considering that IGF-2 generally acts by binding to its receptors to transduce signals (Hawkes et al., 2006), we next asked whether IGF-2R expression changed during PD development.To test this, we performed western blot analysis of IGF-2R protein expression levels(Figure 7E–J) and found that, in thein vivo
experiments, IGF-2R expression was dramatically reduced (P
< 0.01,vs.
sham group; Figure 7E and H).In parallel,we found in thein vitro
experiments that the effect of 6-OHDA treatment on IGF-2R protein expression is time-dependent.As the duration of 6-OHDA injury increased, IGF-2R expression level gradually decreased (P
< 0.05,vs
.control group; Figure 7F and I).Next, we assessed the effect of treatment with IGF-2 on IGF-2R expression.As can be seen from Figure 7G and J, IGF-2 treatment prevented the 6-OHDA-induced down-regulation of IGF-2R in atime-dependent manner (P
< 0.05,vs
.0 h group).The above results suggest that IGF-2 can inhibit 6-OHDA-induced neuronal damage in combination with IGF-2R.Given our previous observation regarding the increase in peripheral serum IGF-2 levels, we speculate that the lack of IGF-2 and its receptor in braintissue may increase negative feedback increase in the periphery.The PI3K/AKT pathway participates in the neuroprotective effect of IGF-2
PI3K/AKT is an important signaling pathway for neuron development, survival,and energy metabolism (Brunet et al., 2001).Therefore, to determine whether the PI3K/AKT pathway is involved in the neuroprotective effects of IGF-2, we assessed PI3K and AKT activity by western blot.Thein vitro
experimental results demonstrated that p-PI3K and p-AKT levels in the 6-OHDA group were significantly lower than those in the control group (P
< 0.001),while IGF-2 pretreatment reversed this effect (Figure 8A–C).To further confirm these results, we detected changes in phosphorylated and total PI3K and AKT protein levels in mouse brain tissue (Figure 8D–F).The results indicated that 6-OHDA inhibits PI3K and AKT activation, while treatment with IGF-2 activated the PI3K/AKT signaling pathway.
Finally, we analyzed the target molecules described above and their interactions using IPA software.The results are shown in Figure 9.Taken together, these results suggest that the PI3K/AKT pathway participates in the neuroprotective effect of IGF-2.

Figure 4|IGF-2 pretreatment reduces the damage to SH-SY5Y cells induced by 6-OHDA.

Figure 5 | Effect of intranasal IGF-2 administration on 6-OHDA-induced injury in mice.
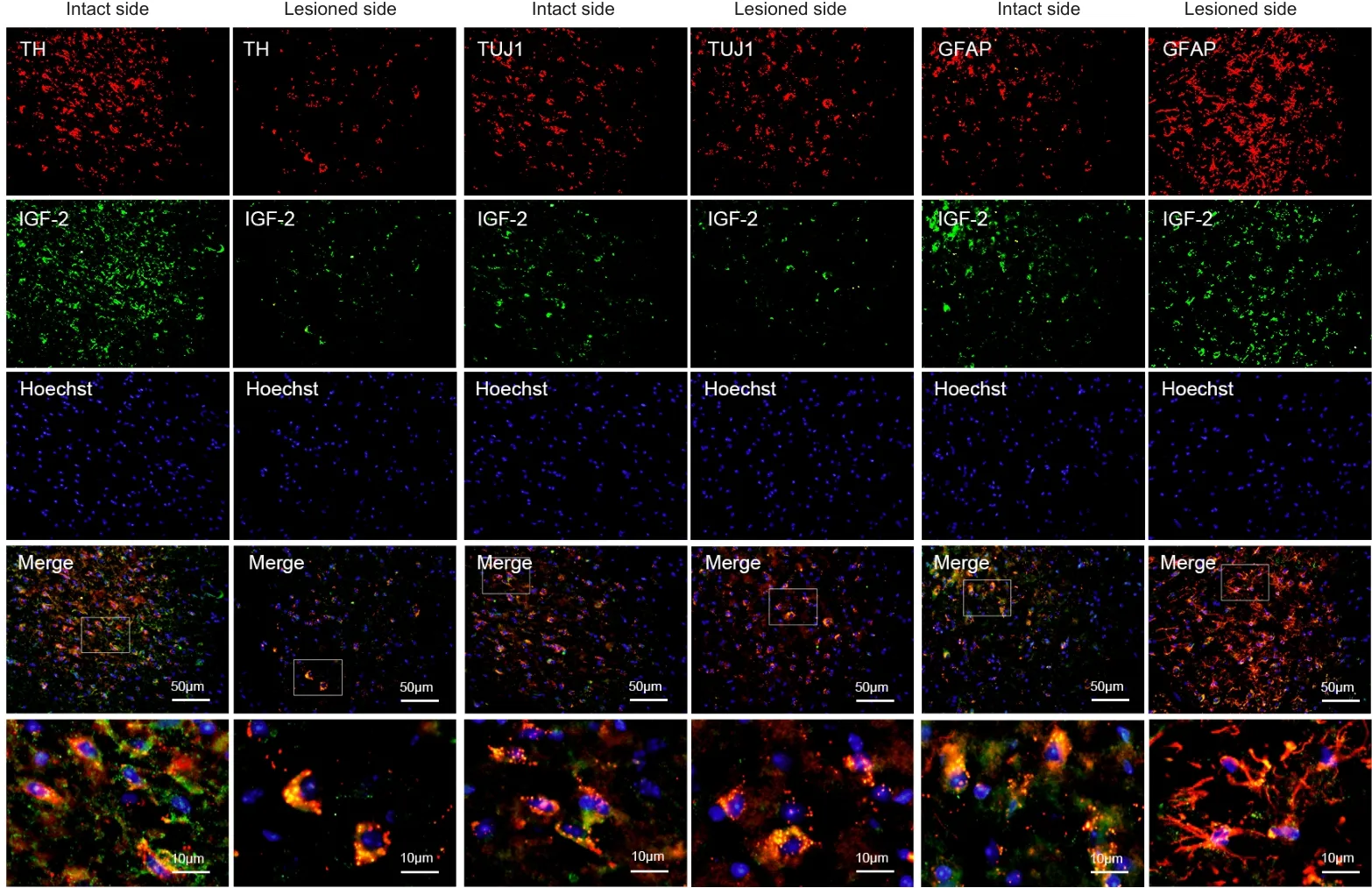
Figure 6|Double immunofluorescence staining for IGF-2 in the midbrain region of a mouse model of PD.

Figure 7|Changes in IGF-2 and IGF-2R expression in PD models.

Figure 8| PI3K and AKT are activated in SH-SY5Y cells and mice pretreated with IGF-2.
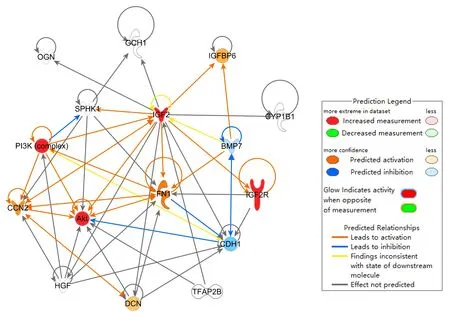
Figure 9| Protein-protein interaction network in PD mice.
Discussion
In recent years, increasing evidence has shown that the protective effect of stem cells is related to the various substances they release (Tran and Damaser,2015; Lin and Du, 2018; Wang et al., 2020).According to our previous studies,SKP-SCs can reduce the damage to dopaminergic neurons induced by 6-OHDA(Chen et al., 2020; Ma et al., 2021; Yan et al., 2022).In this study, we verified that IGF-2 is a key component by which SKP-SCs exert their protective effect and studied the neuroprotective effects of IGF-2 in 6-OHDA-induced cell and mouse models of PD.
SKP-SCs differentiate from SKPs when stimulated by the appropriate growth factors (Hunt et al., 2009), and have proven effective for treating acute and chronic nerve injuries (Khuong et al., 2014).A growing body of evidence has shown that transplanted cells can be therapeutically beneficial as a result of their pleiotropic properties, in that they directly replace damaged or lost cells and secrete paracrine factors, such as IGF-1 (Wang et al., 2014), IGF binding proteins (Jeon et al., 2017; Son et al., 2019) and stromal cell-derived factor-1α (Wang et al., 2010).Our previous studies confirmed that SKP-SCs reduced the damage that 6-OHDA causes to dopaminergic neurons, and, through proteomic analysis of concentrated conditioned medium from SKP-SCs,we identified IGF-2 as a key component that may have a protective effect.The objectives of the current study were to further ascertain whether SKPSCs play an effective therapeutic role by secreting IGF-2 and to determine whether this neurotrophic factor could be used for the treatment of PD.
We found that adding an anti-IGF-2 antibody to SKP-SCs-CM inhibited the neuroprotective effects of SKP-SCs-CMin vitro
.Based on this, we searched the protein interaction database and related literature and discovered that IGF-2 expression decreased significantly in the late disease stage in a 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced mouse model of PD.This prompted us to further investigate whether IGF-2 has a definite protective effect in the context of PD.We first explored possible changes in IGF-2 expression in peripheral serum samples from a mouse model of PD and patients with PD.Surprisingly, IGF-2 expression was increased in the peripheral serum in both cases, which led us to explore this phenomenon in more detail.IGF-2 can traverse the blood-brain barrier and play an endocrine role, and is highly expressed in the nervous system during early fetal development(Beletskiy et al., 2021).Increasing numbers of studies have shown that IGF-2 is a neuroprotective agent.Exogenous low-dose IGF-2 ameliorates negative age-related changes in rats by reducing oxidative damage, increasing antioxidant enzyme activity, and increasing mitochondrial function (Castilla-Cortázar et al., 2011).Intracerebroventricular injection of IGF-2 can rescue partial damage caused by occlusion of large intracerebral arteries and play a critical part in brain recovery after stroke (Vafaee et al., 2020).In amyotrophic lateral sclerosis, IGF-2 can inhibit glycogen synthase kinase through the PI3K/AKT pathway, promote the retention of motor neurons, and induce nerveregeneration (Allodi et al., 2016).Recently, studies have demonstrated the physiological functions of IGF-2 in cognition, memory, and learning.Chen et al.(2011) first demonstrated that IGF-2 administration in rats can significantly improve memory retention and prevent memory loss.Several later studies confirmed that IGF-2 can enhance memory (Stern et al., 2014;Lee et al., 2015; Yu et al., 2020).Taken together, these studies show that IGF-2 can play a therapeutic role in multiple neurological diseases, including neurodegenerative diseases.
However, few studies have investigated the ameliorating effects of IGF-2 on neuronal injury in PD, and even fewer studies have addressed this topic in the 6-OHDA-induced mouse model of PD.Martín-Montañez et al.(2021)described the protective effects of IGF-2 in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine–induced neuronal culture and animal models of PD.Their experiments revolved around oxidative stress and mitochondrial function.Although the experimental models and methods they used and the specific research mechanisms studied were different from ours, the results point to a similar conclusion: that IGF-2 is neuroprotective in PD models.In addition,owing to abundant evidence that autophagy is linked to PD progression(Zhang et al., 2015; Bonam et al., 2021; Lizama and Chu, 2021), Sepúlveda et al.(2022) explored possible changes in the expression of autophagy-related genes and IGF-2 in PD.Their results showed that PD patients had altered levels of IGF-2, as well as autophagy-related genes, and that these changes in expression were correlated (Sepúlveda et al., 2022).The IGF-2 levels they measured in PD patients’ plasma were significantly lower than those detected in the control subjects’ plasma.This contradicts our results showing elevated levels of IGF-2 in the serum of patients and a 6-OHDA–induced mouse model of PD, and indeed results from many previous studies showing that IGF-1 is elevated in PD patients’ serum (Godau et al., 2010, 2011; García-Huerta et al.,2020).Our hypothesis is that these discrepancies are related to variations in geographical, clinical, and ethnic factors.In addition, experimental results can also be affected by sample size and detection technique.
We found that,in vitro
, the neuroprotective effect of IGF-2 is dosedependent, and that treatment with the appropriate concentration of IGF-2 can confer resistance to the cytotoxic and apoptotic effects of 6-OHDA.In addition, intranasal administration of recombinant IGF-2 to a mouse model of PD reversed the 6-OHDA–induced behavioral motor deficits, DA neuron loss,and α-syn aggregation.Intranasal administration has been gradually adopted in recent years because of its many advantages.First, unlike intravenous administration and direct intraventricular injection, it is a non-invasive approach.Second, intranasal delivery delivers the drug along the olfactory and trigeminal nerves, a route that bypasses the blood-brain barrier, allowing direct delivery of exogenous substances into the brain (Lochhead and Thorne,2012; Aly and Waszczak, 2015; Lochhead et al., 2019; Long et al., 2020; Wu et al., 2021).To explore the mechanism underlying the effects described above, we detected IGF-2 localization in the mouse midbrain and found that it was expressed in dopaminergic neurons and surrounding glial cells and neurons.Western blot results suggested that, both in mouse midbrain and in SH-SY5Y cells, IGF-2 expression is reduced after 6-OHDA-induced injury, unlike the increase in IGF-2 expression seen in the peripheral serum of PD patients and the PD mouse model.Because IGF-2 generally binds to its receptor to exert its effects, we assessed the expression of its receptor IGF-2R in the PD mouse model and found that it was reduced compared with that in control mice.Furthermore,in vitro
results illustrated that IGF-2R expression decreased gradually as 6-OHDA injury time increased, and that treatment with IGF-2 rescued this effect.This result is consistent with the study by Martín-Montañez et al.(2021).We speculate that the neuroprotective effect of IGF-2 in PD may be mediated by IGF-2R.However, it remains unclear why IGF-2 expression increases in the peripheral serum but decreases in the central brain tissue.A review of the literature showed that there are many precedents for similar apparent contradictions in neurological diseases.For example,insulin and IGF-1 expression are decreased in the brain and increased in the peripheral serum of Alzheimer’s disease patients, and administering insulin or IGF-1 can improve cognition and regulate β-amyloid (Craft et al., 1998; Hoyer,2004; Horvath et al., 2020; Selles et al., 2020).Similarly, in PD, IGF-1 function is impaired, but IGF-1 expression in the circulation increases (Kao, 2009; Picillo et al., 2013; Fan et al., 2018).We speculate that, during the chronic injury that occurs during PD, IGF-2 and IGF-2R production in the central nervous system decreases, resulting in an increase in negative feedback from IGF-2 in the peripheral serum.However, balance is not completely restored, possibly because the amount of IGF-2 produced in the periphery is insufficient, and the efficiency with which it crosses the blood-brain barrier is inadequate to rescue expression in the brain tissue.At the same time, reduced IGF-2R expression prevents IGF-2 from exerting its effects.Thus, administration of IGF-2 at an early stage of the disease could prevent the extreme deficiency of IGF-2 and IGF-2R in brain tissue.Moreover, intranasal administration bypasses the blood-brain barrier, so, compared with subcutaneous injection or intravenous injection, small doses of exogenous IGF-2 administered intravenously could be a potent promoter of neuronal cell survival.The PI3K/AKT pathway performs an indispensable role in maintaining cell survival and inhibiting apoptosis (Brunet et al., 2001).AKT activates the pathway by promoting the binding of upstream growth factors to their receptors.Our results indicate that this pathway plays a key role in the protective effect of IGF-2 in PD models.After 6-OHDA treatment, p-AKT and p-PI3K levels decreased, and treatment with IGF-2 reversed these changes.This suggests that IGF-2 enhances AKT and PI3K phosphorylation through IGF-2R signaling, thereby activating the anti-apoptotic PI3K/AKT signaling pathway.
Although our results are significant, the study had some limitations.Because we only collected data at a single time point, we cannot draw any conclusions about the effect of IGF-2 treatment over time.In addition, we did not explore whether IGF-binding protein are involved in the IGF-2 signaling changes observed in PD.Therefore, the results should be interpreted with caution.We speculate that IGF-2 may play a neuroprotective effect in PD through IGF-2R/PI3K/AKT signaling.Finally, only male mice were used in our study, as male mice are more resistant and recover better than female mice after injury;indeed, almost all published studies 6-OHDA–induced PD also used male mice.
Despite these limitations, this study clearly demonstrated the effect of IGF-2 on 6-OHDA-induced models PD and characterized changes in IGF-2 expression in PD.Our results suggest that IGF-2 has many effects, such as promoting neuronal survival, inhibiting apoptosis, and degrading toxic protein aggregates.Upregulating IGF-2 in the brain may be an important avenue for therapeutic development in the future.Based on the available literature and the current study, we believe that targeting IGF-2 expression could benefit patients with PD in the future.
Acknowledgments:
We thank Dr.Mei Liu and Dr.Cheng-Bin Xue (both from Key Laboratory of Neuroregeneration of Jiangsu and Ministry of Education,Co-innovation Center of Neuroregeneration of Nantong University) for assistance with the experiments.Graphical Abstract was designed by Figdraw(www.figdraw.com, accessed on May 19, 2022).
Author contributions:
Study design: XSG, KFK; experiment implementation:HYZ, YCJ, JNY; data analysis: JRL, XJW, JBS; reagents/materials/analysis tools support: JBS, KFK; manuscript draft: HYZ, XSG.All authors have read and approved the final manuscript.
Conflicts of interest:
The authors declare that they have no conflicts of interest.
Open access statement:
This is an open access journal, and articles are distributed under the terms of the Creative Commons AttributionNonCommercial-ShareAlike 4.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
Open peer reviewer:
Sanusi Mohammad Bello, King Faisal University, Saudi Arabia.

- 中国神经再生研究(英文版)的其它文章
- The effects and potential of microglial polarization and crosstalk with other cells of the central nervous system in the treatment of Alzheimer’s disease
- Novel therapeutic strategies targeting mitochondria as a gateway in neurodegeneration
- Vimentin as a potential target for diverse nervous system diseases
- Clemastine in remyelination and protection of neurons and skeletal muscle after spinal cord injury
- Artificial nerve graft constructed by coculture of activated Schwann cells and human hair keratin for repair of peripheral nerve defects
- The critical role of the endolysosomal system in cerebral ischemia