
广西普通野生稻渗入系耐低氮和氮素利用率的鉴定评价
2022-11-11 02:41张宗琼农保选杨行海蒋显斌李丹婷夏秀忠
热带作物学报 2022年10期
荘 洁,张宗琼,农保选,杨行海,蒋显斌,李丹婷,夏秀忠
广西普通野生稻渗入系耐低氮和氮素利用率的鉴定评价
荘 洁,张宗琼*,农保选,杨行海,蒋显斌,李丹婷,夏秀忠**
广西壮族自治区农业科学院水稻研究所/广西水稻遗传育种重点实验室,广西南宁 530007
以广西普通野生稻Y11为供体,籼型恢复系GH998为受体构建的166份野生稻渗入系BC3F5为研究对象,设置不施氮(LN)和正常施氮(NN)2种处理,调查株高、有效穗和穗长等12个表型性状,结合耐低氮(NDT)性状、氮素利用率(NUE)性状进行综合评价,以筛选出可利用的氮高效利用育种材料。结果表明:渗入系所有表型均值NN处理均高于LN处理,受施氮影响较大的性状为茎叶含氮量和谷物含氮量,增幅分别为56.89%和37.89%;影响较小的性状为穗长和千粒重,增幅分别为0.50%和2.76%;除穗长外,11个性状在2个施氮处理间呈极显著差异,而且穗总粒数、穗实粒数呈正态分布。渗入系NDT性状的相对生物产量(RBY)与相对生物含氮量(RBN)均值居于双亲之间,且RBN为正态分布;相对谷物产量(GRY)和相对谷物含氮量(RGN)均值高于双亲且趋近GH998,均为正态分布;说明渗入系的耐低氮能力与野生稻的高生物产量和高谷物含氮量特性相关。NUE性状均值处于双亲之间且趋近于GH998,说明氮素高效利用能力主要来源于栽培稻;NDT和NUE性状之间呈显著负相关,因此提高水稻耐低氮能力会降低氮高效利用率。NDT性状的主要影响因子是谷物含氮量和茎叶干重;NUE性状主要影响因子是有效穗、单株产量和谷物产量等。因此,依据研究结果对渗入系进行综合评价筛选,获得3个耐低氮、12个高氮素利用率和3个氮高效且耐低氮的株系,为水稻氮高效育种提供理论依据和育种材料。
普通野生稻;栽培稻;渗入系;耐低氮;氮素利用率
中国是全球水稻总产量和消费量最大的国家[1]。对于水稻生产而言,氮素的影响仅次于水,是水稻生产成本的主要构成部分,而中国的水稻氮肥消耗量占全球水稻氮肥总消耗量的37%,占全国氮肥总消耗量的24%左右[2]。在世界主要稻区中,中国的稻田氮肥利用率相当低,为30%~ 35%[3]。提高氮肥利用率是目前水稻生产亟需解决的突出问题,而水稻自身氮素利用效率的挖掘和利用是提高氮肥利用率的重要生物学途径[4-6]。
普通野生稻(Griff.)作为亚洲栽培稻的祖先种,具有丰富的遗传多样性和多种优良特性,是栽培稻遗传改良的宝贵资源[7-9]。广西作为亚洲栽培稻的起源中心之一[10],普通野生稻资源类型多样[11],遗传多样性丰富[12],但在营养高效利用率方面的研究鲜见报道。从普通野生稻中发掘和利用栽培稻中已经丢失或削弱的基因,加速利用远缘野生种的有利基因,扩大现有栽培稻品种的遗传基础,已引起水稻育种学家的普遍关注。ZHOU等[13]研究结果表明包括东乡野生稻等34个中国野生稻群体的剑叶叶片氮素含量与土壤氮素含量呈显著负相关,说明野生稻具有氮素高效利用基因[13]。许多学者对东乡野生稻耐低氮性进行了大量试验和研究,并定位多个耐低氮相关的QTL[14-16]。LIU等[17]发现调控水稻分蘖基因,可提高氮素利用率20%~30%,该基因在大量野生稻中普遍存在,但在栽培稻中大量丢失。因此,挖掘野生稻氮素利用基因,对培育耐低氮品种及提高水稻氮素利用率具有重要的意义。
本研究以广西普通野生稻Y11与籼型广谱恢复系GH998构建的BC3F5渗入系为研究对象,在2个供氮水平下进行鉴定评价,通过表型、NDT和NUE性状进行综合评价,分析水稻主要表型性状与氮素利用的相关性等,筛选耐低氮和氮高效利用的水稻材料。以期深入了解普通野生稻渗入系在耐低氮、氮素高效利用方面的表现,为水稻氮素高效利用育种提供育种材料,并为挖掘普通野生稻耐低氮基因提供科学依据。
1 材料与方法
1.1 材料
研究材料来源于由广西普通野生稻Y11与籼型广谱恢复系GH998构建的166个BC3F5渗入系株系,种植于广西农业科学院水稻研究所试验田。试验田为水稻土,pH为6.48,有机质含量17.05 g/kg,全氮含量0.12%,全磷含量0.11%,全钾含量1.78%,速效氮含量90.50 mg/kg,速效磷含量34.00 mg/kg,速效钾含量198.50 mg/kg。
1.2 方法
1.2.1 试验设计 试验设LN(尿素0 kg/hm2)和NN(尿素326.1 kg/hm2)处理,在秧苗移栽后第5、25天分别施尿素228.3、97.8 kg/hm2;KCl 116.7、50 kg/hm2和P2O5833.3 kg/hm2全部作为基肥。小区面积20 m2,行距23.1 cm,株距13.2 cm,设置3次重复。水稻植株成熟后,取种植小区中间部分长势一致、有效穗接近平均有效穗值的代表性植株12株,沿地面割取,茎叶和籽粒分开于105℃烘箱杀青30 min,然后在75℃条件下烘4 d至恒重,使用电子天平(分辨率0.001 g)称重,粉碎。调查12个表型性状:株高(cm)、有效穗数、穗长(cm)、穗总粒数、穗实粒数、千粒重(g)、结实率(%)、单株产量(g)、谷物干重(t/hm2)、茎叶干重(t/hm2)、谷物含氮量(kg/hm2)和茎叶含氮量(kg/hm2)。
1.2.2 全氮含量的测定 采用植物中氮、磷、钾的测定(NY/T 2017—2011)方法测定谷物全氮浓度和茎叶全氮浓度,分别在LN和NN处理下测定谷物干重、茎叶干重、谷物含氮量和茎叶含氮量。
1.2.3 NDT和NUE性状计算 计算公式参考WEI等[18]的研究方法。公式如下:
(1)NDT性状。
相对谷物干重(RGY)=0/F×100%
相对生物产量(RBY)=0/F×100%
相对谷物氮含量(RGN)=0/F×100%
相对生物氮含量(RBN)=0/F×100%
(2)NUE性状。
氮反应(NR, kg/hm2)=F–0
产量反应(GR, t/hm2)=F–0
氮素生理利用率(PE, t/kg)=(F–0)/ (F–0)
氮素农学利用率(AE, t/kg)=(F–0)/N
氮素吸收利用率(RE)=(F–0)/N×100%
式中,0为不施氮谷物干重,t/hm2;F为正常施氮谷物干重,t/hm2;0为不施氮生物产量,t/hm2;F为正常施氮生物产量,t/hm2;0为不施氮谷物氮含量,kg/hm2;F为正常施氮谷物氮含量,kg/hm2;0为不施氮植株总含氮量,kg/hm2;F为正常施氮植株总含氮量,kg/hm2;N为氮肥施用量,kg/hm2。
1.3 数据处理
以研究材料表型鉴定和全氮测定结果为基础数据,利用Microsoft Excel 2016软件进行数据整理并计算平均值、标准差和变异系数等。再利用IBM SPSS Statistics 24软件进行方差分析、Pearson相关系数分析和正态检验(Shapiro- Wilk)。最后利用Origin pro 2019b软件进行表型分布频率图和韦恩图的绘制。
2 结果与分析
2.1 表型性状的统计分析
双亲和渗入系在LN和NN处理下的表型存在较大的遗传差异,各性状的变化趋势和幅度不一致,如表1。Y11在正常施氮条件下,株高、有效穗、茎叶干重、谷物干重和谷物含氮量表现升高,穗长、穗总粒数等7个性状表现下降。谷物干重和谷物含量氮量增幅最高分别为74.47%和69.49%,降幅最大为穗实粒数–43.85%和单株产量–39.52%,降幅最小为穗长和茎叶含氮量–1.64%和–2.07%。GH998在正常施氮条件下所有表型均表现增加,但增加的幅度不同,其中增幅最大为单株产量和茎叶含氮量,分别为57.14%和48.02%;最小为千粒重和穗长,分别为3.43%和2.48%。渗入系所有表型均值NN均高于LN处理,受施氮影响最大的性状为茎叶含氮量和谷物含氮量,增幅分别56.89%和37.89%;影响最小的性状为穗长和千粒重,增幅分别为0.50%和2.76%。渗入系12个性状在2个处理间的表型均值变化趋势与栽培稻GH998一致;株高、有效穗等5个性状与野生稻变化趋势一致,穗总粒数和千粒重等7个性状与野生稻变化趋势相反。
在LN和NN处理条件下,166份渗入系12个表型的变化规律,如图1。在2个处理间的表型存在较大遗传差异,表型平均值在NN均高于LN处理。除穗长外,株高、有效穗等11个性状在处理间为极显著差异。正态检验(Shapiro-Wilk)结果表明,穗总粒数、穗实粒数和千粒重在LN和NN处理下均呈正态分布;穗长、千粒重、结实率和谷物含氮量在LN处理下,数量呈现正态分布;其余性状为非正态分布。
2.2 NDT和NUE性状分析
双亲和渗入系在NDT性状和NUE性状中表现出不同的遗传差异,如表2。NDT性状中,GH998的RGY、RGN显著高于Y11;Y11的RBY、RBN显著高于GH998。渗入系的RBY与RBN均值居于双亲之间,GRY和RGN均值高于双亲且趋近GH998;渗入系RBY(17.52)的变异系数大于RGY(10.06)、RGN(25.43)的变异系数大于RBN(17.71),说明渗入系的生物产量和谷物氮含量更容易受施氮的影响。说明野生稻的NDT特性主要体现在RBY和RBN,栽培稻NUE主要体现在RGY和RGN,渗入系在遗传了野生稻的高生物产量特性,同时遗传了栽培稻的高产特性,从而在稳定生物产量的同时提高了谷物干重。NDT性状中的RGY、RGN和RBN为正态分布,RBY非正态分布。
NUE性状中,GH998的NR、GR、AE和RE显著高于Y11,PE在双亲之间表现不显著。渗入系所有NUE性状均为非正态分布,平均值均处于双亲之间且趋近于GH998。说明渗入系的高氮素利用率主要来源于栽培稻亲本,其吸收氮的能力,谷物产量和氮转化产量能力均偏向栽培稻。
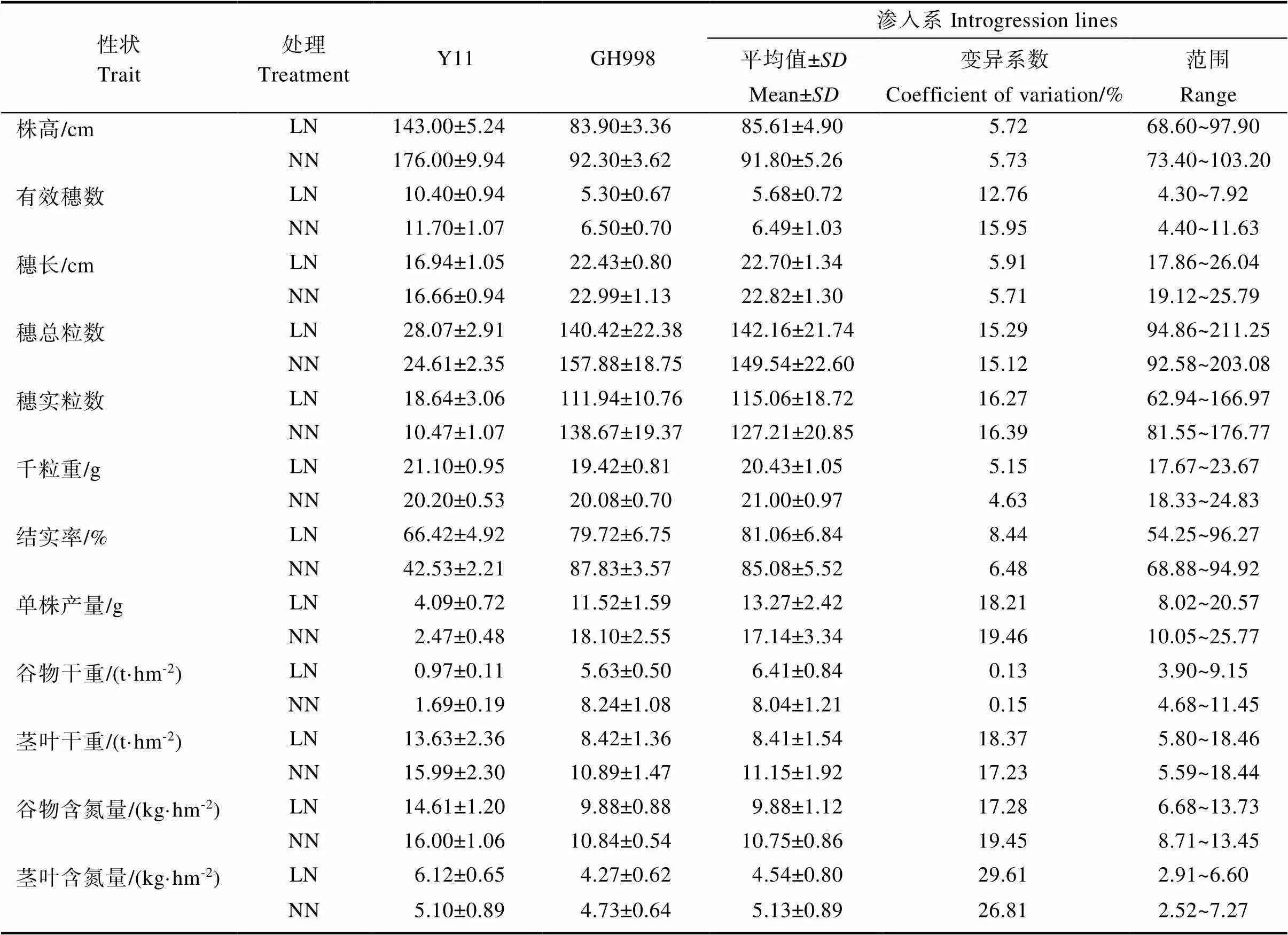
表1 亲本和渗入系在LN和NN处理下的12个表型性状表现

ns表示无显著差异;**表示差异极显著(P<0.01)。
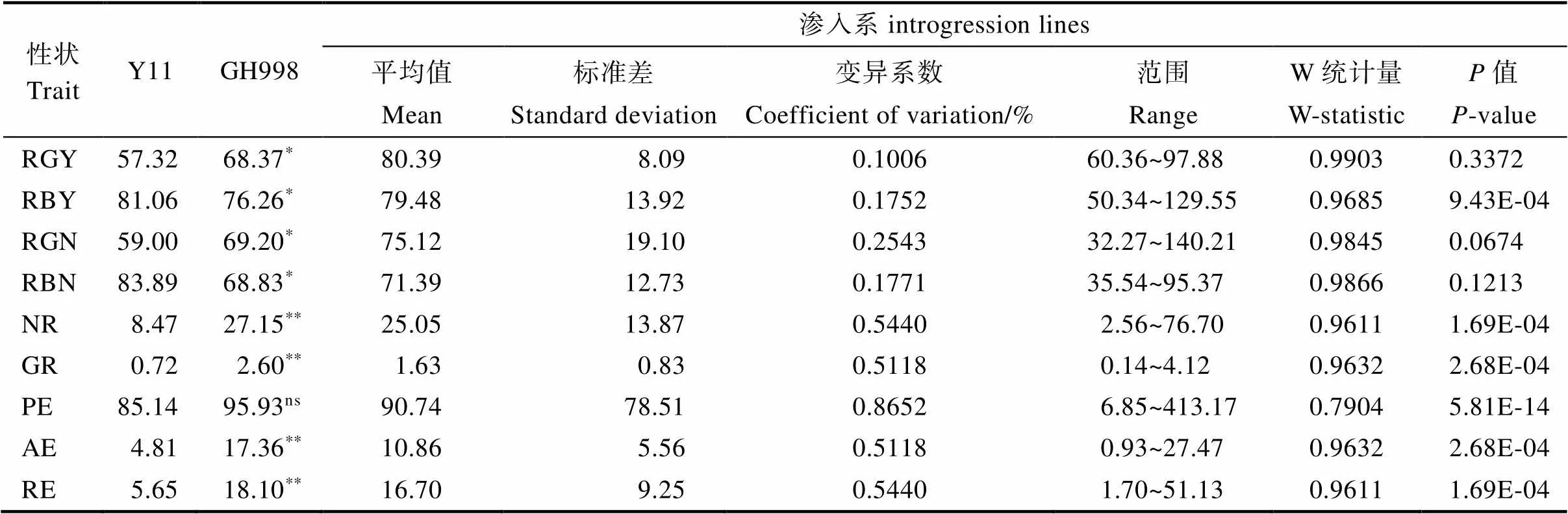
表2 双亲及渗入系NDT和NUE性状的表现
注:*表示差异显著(<0.05),**表示差异极显著(<0.01)。
Note:*indicate significant difference (<0.05),**indicate extremely significant difference (<0.01).
NDT和NUE性状之间相关分析结果见表3。NDT性状之间均呈显著正相关;NUE性状之间除PE与NR、RE呈负相关之外,均呈正相关;NDT和NUE性状之间除PE与RBY、RGN和RBN呈正相关之外,均呈显著负相关。说明提高水稻的氮素利用率会降低耐低氮能力。
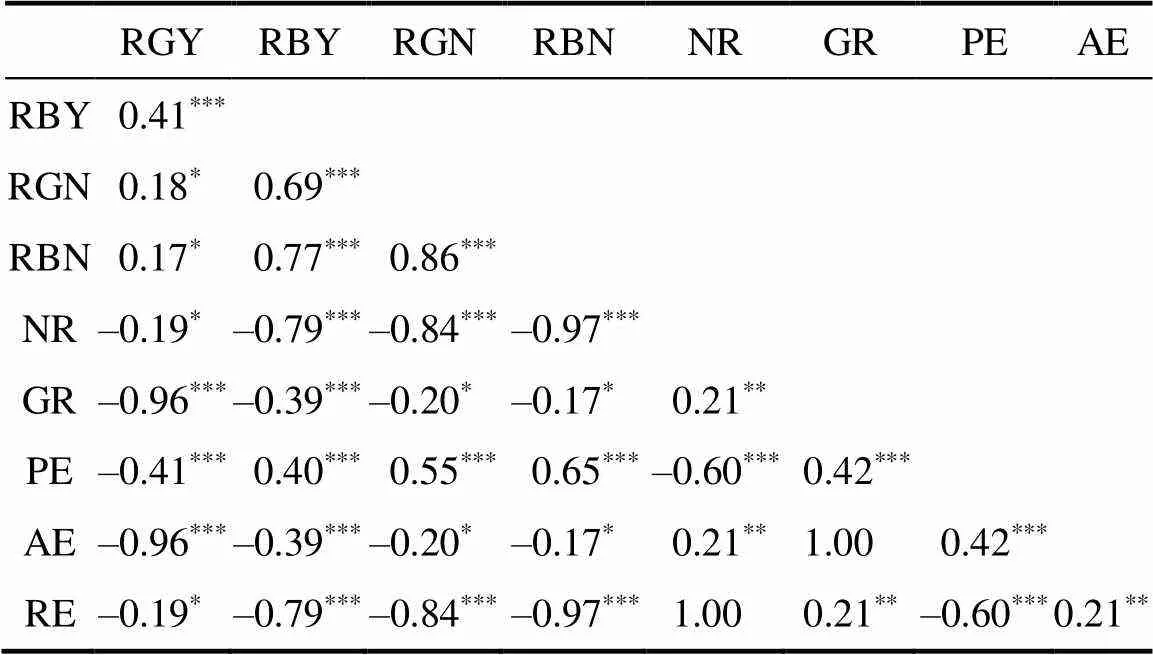
表3 NDT和NUE性状的相关分析
注:*表示显著相关(<0.05);**表示极显著相关(<0.01),***表示极显著相关(<0.001)。
Note:*indicates significant correlation (<0.05);**indicates extremely significant correlation (<0.01),***indicates extremely significant correlation (<0.001).
2.3 NDT、NUE与表型性状之间的相关性分析
NDT、NUE与表型性状相关分析结果见表4,NDT部分性状在LN处理下呈正相关,但在NN处理下呈负相关。如RBY、RGN和RBN在LN处理下与茎叶干重和谷物氮含量表现显著正相关,在NN处理下RBY、RGN和RBN与有效穗、单株产量、茎叶干重和谷物含氮量呈显著负相关;RGY与谷物干重在LN处理下表现正相关,在NN处理下与株高和谷物干重等8个性状表现负相关;但RGN与茎叶含氮量在LN处理下呈负相关,NN处理下呈正相关,与其他性状相反。说明随着施氮量的增加,水稻的耐低氮能力显著下降,其中最大的影响因子是谷物含氮量和茎叶干重。
NUE性状在NN处理下显著相关的性状明显多于LN处理,主要集中在与谷物产量相关的表型性状,如有效穗、谷物干重等。在LN处理下,NR、RE与株高、茎叶干重和谷物含氮量呈负相关外,其余相关均呈正相关;在NN处理下,NR、RE与株高呈负相关,PE与有效穗呈负相关,其余相关均呈正相关。说明随着施氮量的增加,表型影响因子增多,相关性增强,氮素利用率上升,其中变化最大的影响因子的是有效穗、单株产量、谷物干重和茎叶含氮量。
2.4 渗入系NDT和NUE性状的综合评价筛选
在166份渗入系株系中,以NDT指标前20%的株系进行综合评价,其中有3个株系4项NDT性状指标较高,14个株系3项NDT性状指标较高(图2A)。以NUE指标前20%株系进行综合评价,其中有12个株系4项NUE性状指标较高,11个株系3项NUE性状指标较高(图2B)。以NDT性状中与生物产量相关的RBY、RBN;NUE性状中与产量性状相关GR、PE和AE前30%的株系进行综合筛选,获得3个在氮素高效利用且具有耐低氮特性的株系(图2C)。这些耐低氮和氮素利用率较高的株系,为氮素利用高效利用提供了较好研究材料,如表5。
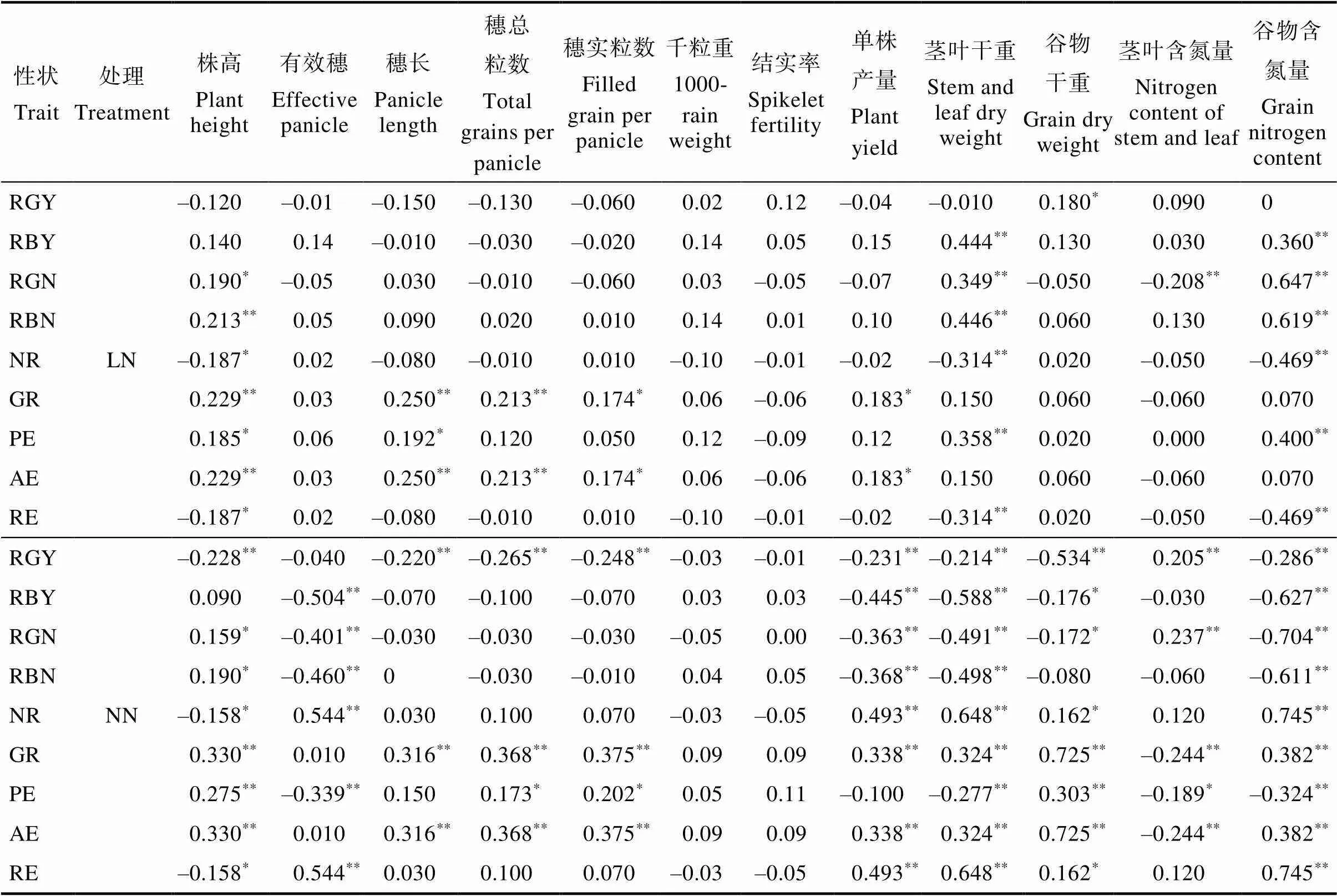
表4 NDT和NUE性状与表型性状之间的相关关系
注:*表示显著相关(<0.05);**表示极显著相关(<0.01)。
Note:*indicates significant correlation (<0.05);**indicates extremely significant correlation (<0.01).
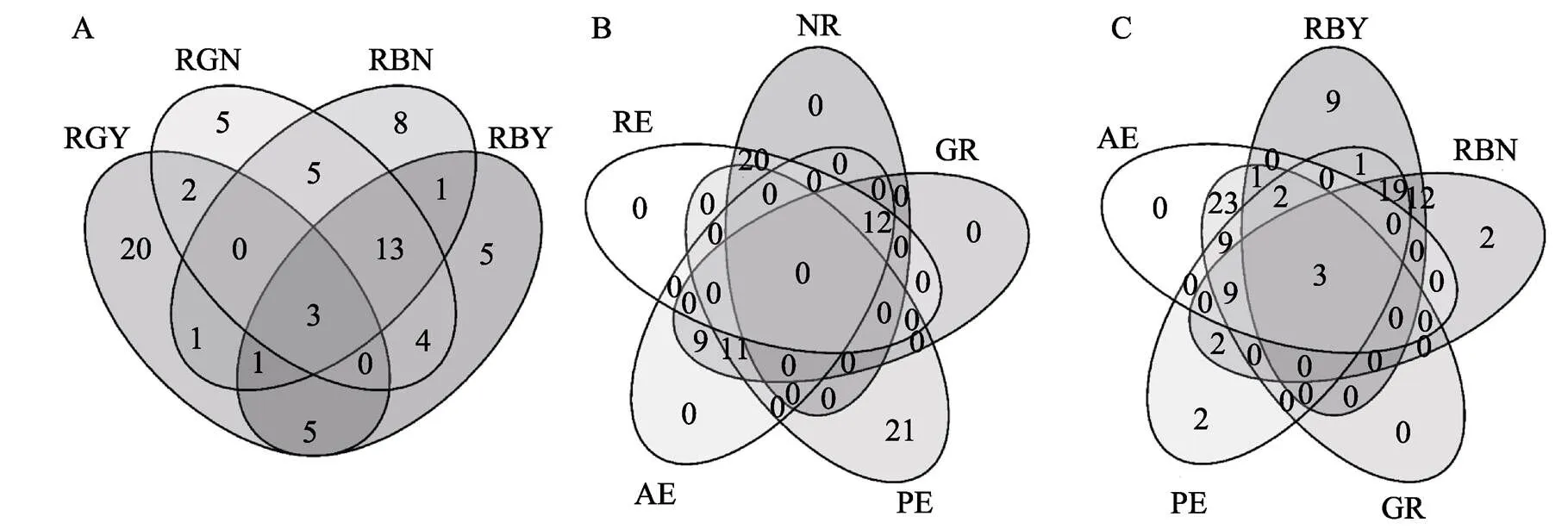
图2 渗入系NDT和NUE性状综合筛选筛选韦恩图
3 讨论
普通野生稻作为亚洲栽培稻的野生祖先种长期生活在野外环境,经过长期的自然选择,保留大量适应自然条件下生长的有利基因,是水稻育种的重要基因来源;而栽培稻经过长期的人为选择,含有符合人类对水稻生产需求的有利基因,是水稻育种的基础基因来源。但水稻氮素利用研究主要集中在栽培稻资源的研究[4, 6, 19-21],野生稻资源研究较少[15, 16, 22],而广西普通野生稻具有非常好的研究价值[10, 23-24]。本研究中,其广西普通野生稻渗入系在处理间变化趋势与栽培稻亲本相同,而东乡野生稻/协青早及BIL群体的穗长和结实率等与双亲变化趋势一致,株高和千粒重变化趋势不同[15],这可能与亲本的选择及后代群体的回交代数不一致有关。如广西普通野生稻Y11在LN和NN处理间穗总粒数、千粒重和结实率等7个性状表现降低,株高、有效穗和茎叶干重5个性状表现升高;而东乡野生稻在正常施氮后株高、千粒重、结实率3个性状表现升高,其他穗实粒数和有效穗等6个性状表现降低[25]。
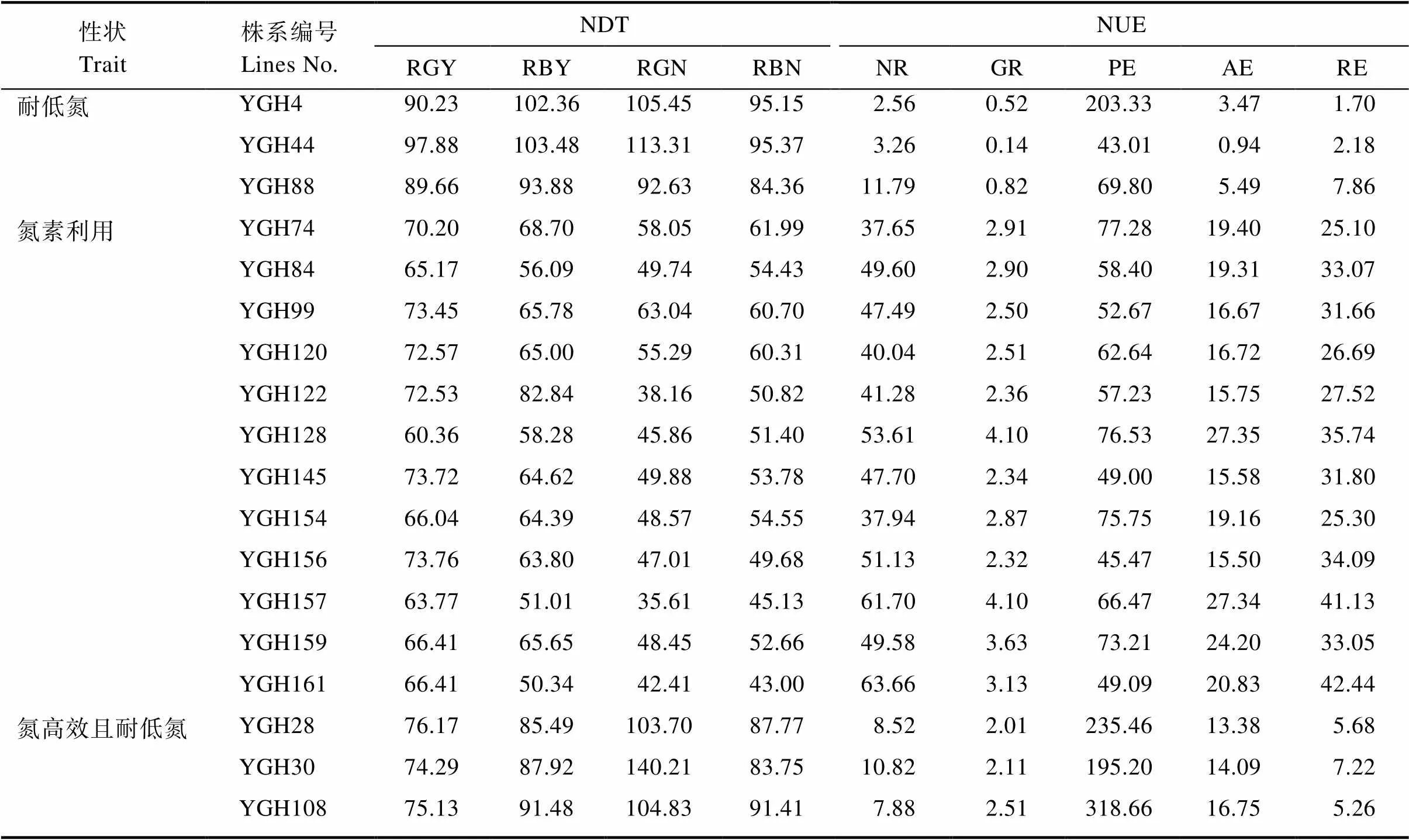
表5 18份渗入系株系的NDT和NUE性状表现
在NDT和NUE指标的选择方面,许多研究学者已经在水稻不同生育期进行了大量的氮素利用研究。曹桂兰等[4]以氮反应指数作为耐低氮指标,认为未施氮或施低氮处理更适合于耐低氮种质的筛选;胡标林等[15]提出株高、总穗粒数和结实率等性状的相对值作为耐低氮评价指标。WEI等[18]利用耐低氮和氮素利用指标进行分析,并进行了QTL定位。本研究利用野生稻和栽培稻构建的渗入系群体,结合表型性状、NDT和NUE性状多项指标进行综合评价,旨在了解三者之间的相关关系,为培育氮高产利用品种提供育种方向和理论基础。
野生稻渗入系与双亲的NDT和NUE性状分析结果表明,(1)渗入系的耐低氮能力主要与野生稻亲本的高生物产量和高谷物含氮量特性相关;渗入系氮素利用率均值接近栽培稻亲本,说明氮素利用率能力主要来源于栽培稻。(2)NDT和NUE在表型性状上主要影响因子并不相同,NDT性状主要影响因子主要与茎叶干重、谷物含氮量等生物产量和谷物含氮性状相关;NUE性状主要与有效穗、穗总粒数等产量性状相关;(3)NDT和NUE性状之间为负相关,说明提高氮素利用率会降低耐低能力,这与BERTRAND等[26]研究结果一致。这是因为植物为了在缺氮条件下生存,往往会改变生理、生化过程和基因表达,如水稻植株通常会通过扩大根系和加深根系来获取土壤中的氮,减少茎叶含氮量,增加种子的含氮量,从而保证本身的繁衍。因此在正常施氮下,具有较高生物产量和氮含量的基因型比具有较低生物产量和氮含量的基因型更容易受到低氮胁迫[27]。
依据表型性状、NDT和NUE性状的相关分析结果,在氮高效利用水稻品种的选育方面,应该选择在低氮条件下,具有较高的生物产量和较低茎叶含氮量的株系,这与阮新民等[22]的研究结果一致。依据这些分析结果,本研究筛选了耐低氮株系、氮素利用率高的株系以及氮素利用率高且耐低氮的株系,这些材料可作为培育氮高效利用品种优异的育种材料。
4 结论
本研究利用广西普通野生稻与栽培稻的渗入系群体,在LN和NN处理下进行分析,对表型性状、NDT和NUE性状进行综合评价,解析各指标之间的相关性,筛选获得耐低氮、氮素高效利用的育种中间材料,为培育氮高效利用品种提供育种材料和品种的筛选方向,在减少水稻氮肥的施用量、降低生产成本、保护环境方面具有重要意义。
[1] FAO. Status of the World’s Soil Resources (SWSR): Main report. Food and Agriculture Organization of the United Nations and intergovernmental technical panel on soils[M]. Rome, 2015.
[2] 彭少兵, 黄见良, 钟旭华, 杨建昌, 王光火, 邹应斌, 张福锁, 朱庆森, ROLAND B, CHRISTIAN W. 提高中国稻田氮肥利用率的研究策略[J]. 中国农业科学, 2002, 35(9): 1095-1103.
PENG S B, HUANG J L, ZHONG X H, YANG J C, WANG G H, ZOU Y B, ZHANG F S, ZHU Q S, ROLAND B, CHRISTIAN W. Research strategy in improving fertilizer- itrogen use efficiency of irrigated rice in China[J]. Scientia Agricultura Sinica, 2002, 35(9): 1095-1103. (in Chinese)
[3] 朱兆良. 农田中氮肥的损失与对策[J]. 土壤与环境, 2000, 1(9): 1-6.
ZHU Z L. Loss of fertilizer N from plants-soil system and the strategies and techniques for its reduction[J]. Soil and Environmental Sciences, 2000, 1(9): 1-6. (in Chinese)
[4] 曹桂兰, 张媛媛, 朴钟泽, 韩龙植. 水稻不同基因型耐低氮能力差异评价[J]. 植物遗传资源学报, 2006, 7(3): 316-320.
CAO G L, ZHANG Y Y, PIAO Z Z, HAN L Z. Evaluation of tolerance to low N-fertilized level for rice type[J]. Journal of Plant Genetic Resources, 2006, 7(3): 316-320. (in Chinese)
[5] 李 敏, 张洪程, 李国业, 魏海燕, 殷春渊, 马 群, 杨 雄. 水稻氮效率基因型差异及其机理研究进展[J]. 核农学报, 2011, 25(5): 1057-1063.
LI M, ZHANG H C, LI G Y, WEI H Y, YIN C Y, MA Q, YANG X. Genotypic difference of nitrogen use efficiency in rice its morphological and physiological mechanisms[J]. Journal of Nuclear Agricultural Sciences, 2011, 25(5): 1057-1063. (in Chinese)
[6] 江立庚, 戴廷波, 韦善清, 甘秀芹, 徐建云, 曹卫星. 南方水稻氮素吸收与利用效率的基因型差异及评价[J]. 植物生态学报, 2003, 27(4): 466-471.
JIANG L G, DAI T B, WEI S Q, GAN X Q, XU J Y, CAO W X. Genotypic differences and valuation in nitrogen uptake and utilization efficiency in rice[J]. Acta Phytoecologica Sinica, 2003, 27(4): 466-471. (in Chinese)
[7] 钱 韦, 葛 颂, 洪德元. 采用RAPD和ISSR标记探讨中国疣粒野生稻的遗传多样性[J]. 植物学报, 2000, 42(7): 741-750.
QIAN W, GE S, HONG D Y. Assessment of genetic variation ofdetected by RAPDs and ISSRs[J]. Acta Botanica Sinica, 2000, 42(7): 741-750. (in Chinese)
[8] 王象坤, 孙传清, 才宏伟, 张居中. 中国稻作起源与演化[J]. 科学通报, 1998, 43(22): 2354-2363.
WANG X K, SUN C Q, CAI H W, ZHANG Z Z. Origin and evolution of rice farming in China[J]. Scientific Bulletin, 1998, 43(22): 2354-2363. (in Chinese)
[9] 孙传清, 李自超, 王象坤. 普通野生稻和亚洲栽培稻核心种质遗传多样性的检测研究[J]. 作物学报, 2001, 27(3): 313-318.
SUN C Q, LI Z C, WANG X K. Studies on evaluation of the genetic diversity of core collection of common wild rice (Griff.) and Asian cultivated rice (L.)[J]. Acta Agronomica Sinica, 2001, 27(3): 313-318. (in Chinese)
[10] HUANG X H, KURATA N, WEI X H, WANG Z X, WANG A H, ZHAO Q, ZHAO Y, LIU K Y, LU H Y, LI W J, GUO Y L, LU Y Q, ZHOU C C, FAN D L, WENG Q J, ZHU C R, HUANG T, ZHANG L, WANG Y C, FENG L, FURUUMI H, KUBO T, MIYABAYASHI T, YUAN X P, XU Q, DONG G J, ZHAN Q L, LI C Y, ASAO F, TOYODA A, LU T T, FENG Q, QIAN Q, LI J Y, HAN B. A map of rice genome variation reveals the origin of cultivated rice[J]. Nature, 2012, 490: 497-501.
[11] 吴妙燊. 浅谈广西野生稻的分布[J]. 遗传, 1981, 3(3): 36-37.
WU M S. On the distribution of wild rice in Guangxi[J]. Hereditas, 1981, 3(3): 36-37. (in Chinese)
[12] 黄 娟, 杨庆文, 陈成斌, 梁世春, 张万霞, 乔伟华, 王家祥. 广西普通野生稻的遗传多样性及分布特征[J]. 中国农业科学, 2009, 42(8): 2633-2642.
HUANG J, YANG Q W, CHEN C B, LIANG S C, ZHANG W X, QIAO W H, WANG J X. Genetic diversity and the geographical characteristics of wild rice (Griff) in Guangxi[J]. Scientia Agricultura Sinica, 2009, 42(8): 2633-2642. (in Chinese)
[13] ZHOU W, WANG Z, XING W, LIU G. Plasticity in latitudinal patterns of leaf N and P ofin China[J]. Plant Biology, 2014, 16(5): 917-923.
[14] 汤国平, 陈小荣, 朱昌兰, 彭小松, 贺晓鹏, 傅军如, 边建民, 胡丽芳, 欧阳林娟, 贺浩华. 协青早B//协青早B/东乡野生稻BC1F13群体及若干杂交稻恢复系低氮耐性鉴定[J]. 江西农业大学学报, 2017, 39(2): 214-222.
TANG G P, CHEN X R, ZHU C L, PENG X S, HE X P, FU J R, BIAN J M, HU L F, OUYANG L J, HE H H. Identification low nitrogen tolerance of Xieqingzao B//Xieqingzao B/Dongxiang wild rice BC1F13populations and some restorers of hybricl rice[J]. Acta Agriculturae Universitatis Jiangxiensis, 2017, 39(2): 214-222. (in Chinese)
[15] 胡标林, 李 霞, 万 勇, 邱在辉, 聂元元, 谢建坤. 东乡野生稻BILs群体耐低氮性表型性状指标筛选及其综合评价[J]. 应用生态学报, 2015, 26(8): 2346-2352.
HU B L, LI X, WAN Y, QIU Z H, NIE Y Y, XIE J K. Index screening and comprehensive evaluation of phenotypic traits of low nitrogen tolerance using BILs population derived from Dongxiang wild rice (Griff.)[J]. Chinese Journal of Applied Ecology, 2015, 26(8): 2346-2352. (in Chinese)
[16] 沈雨民, 洪 骏, 熊焕金, 陈明亮, 吴小燕, 邓 伟, 肖叶青. 利用重组自交系定位东乡野生稻在低氮胁迫下相关性状QTL[J]. 分子植物育种, 2019, 17(11): 3615-3623.
SHEN Y M, HONG J, XIONG H J, CHEN M L, WU X Y, DENG W, XIAO Y Q. Identification of related traits QTL stressed with nitrogen deficiency by recombinant inbred lines of Dongxiang wild rice[J]. Molecular Plant Breeding, 2019, 17(11): 3615-3623. (in Chinese)
[17] LIU Y, WANG H, JIANG Z, WANG W, XU R, WANG Q, ZHANG Z, LI A, LIANG Y, OU S, LIU X, CAO S, TONG H, WANG Y, ZHOU F, LIAO H, HU B, CHU C. Genomic basis of geographical adaptation to soil nitrogen in rice[J]. Nature, 2021, 590(7847): 600-605.
[18] WEI D, CUI K, YE G, PAN J, XIANG J, HUANG J, NIE L. QTL mapping for nitrogen-use efficiency and nitrogen-deficiency tolerance traits in rice[J]. Plant and Soil, 2012, 359(1-2): 281-295.
[19] 程建峰, 戴廷波, 曹卫星, 姜 东, 刘宜柏. 不同类型水稻种质氮素营养效率的变异分析[J]. 植物营养与肥料学报, 2007, 13(2): 175-183.
CHENG J F, DAI T B, CAO W X, JIANG D, LIU Y B. Variations of nitrogen nutrition efficiency in different rice germplasm types[J]. Plant Nutrition and Fertilizer Science, 2007, 13(2): 175-183. (in Chinese)
[20] 黎毛毛, 万建林, 黄永兰, 曹桂兰, 陈红萍, 韩龙植. 水稻微核心种质氮素利用率相关性状的鉴定评价及其相关分析[J]. 植物遗传资源学报, 2011, 12(3): 352-361.
LI M M, WAN J L, HUANG Y L, CAO G L, CHEN H P, HAN L Z. Evaluation and correlation analysis of the related characters of nitrogen use efficiency for mini core collection of rice (L.) in China[J]. Journal of Plant Genetic Resources, 2011, 12(3): 352-361. (in Chinese)
[21] 朴钟泽, 韩龙植, 高熙宗. 水稻不同基因型氮素利用效率差异[J]. 中国水稻科学, 2003, 17(3): 233-238.
PIAO Z Z, HAN L Z, KOH H J. Variations of nitrogen use efficiency by rice genotype[J]. Chinese Journal of Rice Science, 2003, 17(3): 233-238. (in Chinese)
[22] 阮新民, 施伏芝, 从夕汉, 罗志祥. 基于重组自交系群体水稻氮素利用效率分析和利用[J]. 中国生态农业学报, 2016, 24(6): 780-789.
RUAN X M, SHI F Z, CONG X H, LUO Z X. Analysis of rice nitrogen use efficiency based on recombinant inbred line population[J]. Chinese Journal of Eco-Agriculture, 2016, 24(6): 780-789. (in Chinese)
[23] 潘英华, 徐志健, 梁云涛. 广西普通野生稻群体结构解析与核心种质构建[J]. 植物遗传资源学报, 2018, 19(3): 498-509.
PAN Y H, XU Z J, LIANG Y T. Genetic structure and core collection of common wild rice (Griff.) in Guangxi[J]. Journal of Plant Genetic Resources, 2018, 19(3): 498-509. (in Chinese)
[24] 薛艳霞, 梁燕理, 冯 璇, 黄金艳, 刘 芳, 覃宝祥, 邱永福, 李容柏. 广西普通野生稻遗传多样性中心的确定与核心种质构建[J]. 华南农业大学学报, 2016, 37(5): 24-30.
XUE Y X, LIANG Y L, FENG X, HUANG J Y, LIU F, QIN B X, QIU Y F, LI R B. Center of genetic diversity and core collection of common wild rice,Griff., in Guangxi[J]. Journal of South China Agricultural University, 2016, 37(5): 24-30. (in Chinese)
[25] 吴 婷, 李 霞, 黄得润, 黄凤林, 肖宇龙, 胡标林. 应用东乡野生稻回交重组自交系分析水稻耐低氮产量相关性状QTL[J]. 中国水稻科学, 2020, 34(6): 499-511.
WU T, LI X, HUANG D R, HUANG F L, XIAO Y L, HU B L. QTL analysis for yield traits related to low nitrogen tolerance using backcrossing recombinant inbred lines derived from dongxiang wild rice (Griff.)[J]. Chinese Journal of Rice Science, 2020, 34(6): 499-511. (in Chinese)
[26] BERTRAND H, JACQUES L G, BERTRAND N, ANDRÉ G. The challenge of improving nitrogen use efficiency in crop plants: towards a more central role for genetic variability and quantitative genetics within integrated approaches[J]. Journal of Experimental Botany, 2007, 58(9): 2369-2387.
[27] NOVOA R, LOOMIS R S. Nitrogen and plant production[J]. Plant and Soil, 1981, 58: 177-204.
Identification and Evaluation of Nitrogen-deficiency Tolerance and Nitrogen-use Efficiency for Introgression Lines of Wild Rice (Griff.) in Guangxi
ZHUANG Jie, ZHANG Zongqiong*, NONG Baoxuan, YANG Xinghai, JIANG Xianbin, LI Danting,XIA Xiuzhong**
Rice Research Institute, Guangxi Academy of Agricultural Sciences / Guangxi Key Laboratory of Rice Genetics and Breeding, Nanning, Guangxi 530007, China
In order to utilize the elite genes of wild rice, 166 BC3F5introgression lines constructed from Guangxi wild rice Y11 as the donor andrestorer line GH998 as the recipient were used as the identification materials. Twelve phenotypic traits were studied under low nitrogen (LN) and normal nitrogen (NN) treatments. Combined with nitrogen-deficiency tolerance (NDT) and nitrogen-use efficiency (NUE) traits, a comprehensive evaluation was conducted in order to select available nitrogen-efficient breeding materials. The results showed that the mean value of 12 phenotypes in NN were higher than that of LN. The nitrogen content of stem-leaf and grains were strongly affected by nitrogen fertilizer, improved by 56.89% and 37.89%, respectively. The panicle length and 1000-grain weight were less affected, improved by 0.50% and 2.76%, respectively. Except for spike length, 11 traits showed extremely significant differences between LN and NN. Total grains per panicle and filled grains per panicle were normal distributed. The mean value of relative biomass yield (RBY) and relative grain yield (RBN) of NDT traits in the introgression lines were located between the parents, and RBN was normal distribution. The mean value of relative grain yield (RGY) and relative grain nitrogen (RGN) were higher than those of the two parents and close to GH998. All NDT traits were normal distribution. It indicated that NDT of introgression lines was correlated with high biomass yield and grain nitrogen content of wild rice. The mean value of NUE traits were located between the two parents and close to GH998, which indicating that the high NUE mainly was affected by cultivated rice. Moreover, NDT was significantly negatively correlated with NUE. So, improving NDT level would reduce the level of NUE. The grain nitrogen content, dry weight of stems and leaves were key impact factors of NDT. The panicle number, yield per plant and grain yield were key impact factors of NUE. Finally, based on these studies, 3 lines with high NDT, 12 lines with high NUE and 3 lines as both traits were comprehensively evaluated, which would provide theoretical basis and special germplasm for rice breeding in nitrogen efficient.
common wild rice; cultivated rice; introgression lines; nitrogen-deficiency tolerance; nitrogen-use efficiency
S511
A
10.3969/j.issn.1000-2561.2022.10.013
2022-01-25;
2022-03-02
国家自然科学基金项目(No. 32060476);广西科技计划项目(桂科AB21238009);中央引导地方科技发展专项资金项目(桂科ZY21195034)。
荘 洁(1986—),女,大专,研究实习员,研究方向:水稻种质资源;*同等贡献作者:张宗琼(1978—),女,硕士,副研究员,研究方向:水稻遗传育种。**通信作者(Corresponding author):夏秀忠(XIA Xiuzhong),E-mail: xiaxiuzhong@163.com。
猜你喜欢
今日农业(2022年13期)2022-09-15
今日农业(2021年17期)2021-11-26
江西农业学报(2021年8期)2021-09-08
今日农业(2021年9期)2021-07-28
上海师范大学学报·自然科学版(2020年5期)2020-12-18
军事文摘(2020年20期)2020-11-16
天津农林科技(2020年1期)2020-04-08
植物研究(2020年6期)2020-03-05
燃料化学学报(2019年3期)2019-04-03
分析化学(2018年8期)2018-11-01
