
距离母竹不同位置长叶苦竹的生长发育特征
2022-11-16 04:13吴艳萍李传哲赵福泽王福升丁雨龙刘国华
江苏农业科学 2022年20期
吴艳萍,李传哲,赵福泽,王福升,丁雨龙,刘国华
(1.南京林业大学,江苏南京 210037;2.江苏徐淮地区淮阴农业科学研究所,江苏淮安 223001;3.桐梓县林业局,贵州遵义 563299)
竹类植物是自然界生长迅速、更新较快的物种之一,也是最具开发潜力、效益最高的可再生林业资源之一[1]。竹子具有显著的经济效益,广泛应用于造纸、制浆、建筑、农业、手工业、家具业、观赏、环保、食品等多个领域[2]。研究竹类植物的生长发育规律,对于合理开发其价值具有相当重要的意义[3-4]。竹子主要分为3种类型:丛生型、散生型、混生型。国内竹类植物生长发育规律相关研究主要以几种常见散生竹和从生竹为研究对象,集中在竹笋-幼竹高生长、生物量分配、竹林结构、竹林培育等几个方面[4]。其中对材用竹的研究偏重于竹材形成方面,如秆形形成、节间生长、材质生长等。
通过研究竹子的秆形建成规律,可以了解竹子的材性、生态习性甚至生理习性,不仅可为其生长规律相关研究提供理论依据,对其竹林的经营管理也大有裨益[5-7]。目前对一些常见散生竹和丛生竹的秆形形态研究已有大量报道。如毛竹[Phyllostachysedulis(Carrière) J.Houz.]节间长度由基部到梢部先增大后减小,近中央节间长度最长,且两端的变化几乎一致,而节间壁厚呈递减趋势[8]。与毛竹相比,同样胸径的大木竹(Bambusawenchouensis)全高、鲜质量大于毛竹,竹壁厚随杆高增大而减小的速度较毛竹快,节间长度随高度的变化趋势呈二次曲线[9]。淡竹(Phyllostachysglauca)平均壁厚及外径由基部到梢部也呈递减趋势[10]。缅甸竹(Bambusaburmanica)的基径和胸径,秆质量和胸径,壁厚和高度均存在较高的相关性[11]。因此,相对于散生竹和丛生竹,目前对混生竹秆形规律的相关研究较少。
长叶苦竹(Pleioblastuschinovar.hisauchiiMakino) 别称狭叶青苦竹,为青苦竹[Pleioblastuschino(Franch.et Savat.) Makino]变种,为大明竹属植物[12]。竹秆多色,且壁厚,有分枝的一侧节间基部微微凹陷,没有分枝的竹秆节间为圆筒形或扁圆筒形。枝叶茂密,密集丛生,是一类良好的混生型观赏竹种,具有重要的园林应用价值[13]。本研究以南京林业大学白马基地造林密度4 m×4 m,造林株数8丛,五年生长叶苦竹试验林为研究对象,调查了竹林的发笋时间、发笋期、退笋率,测量了竹笋-幼竹发育过程后中的高度、地径等及距离母竹不同位置竹株的秆形形态指标及生物量,研究林缘及林中的竹笋-幼竹高生长规律及距离母竹不同位置竹株的秆形形态建成规律和生物量分配,旨在为日后改善混生型竹林的管理经营提供科学的理论依据。
1 材料与方法
1.1 时间与地点
试验时间为2019年4月至2020年12月,试验地点为江苏省南京市溧水区南京林业大学白马科研基地竹类植物种质资源圃(119°11′7″E,31°36′51″N)。
1.2 材料与方法
2016年3月,将8丛(蔸)二年生长叶苦竹母竹从育苗池移植于土壤配比为黄心土-草炭-珍珠岩=60-25-15,基质厚度为45 cm的土层中,株行距为4 m×4 m,母竹宿土直径为40 cm×40 cm,每丛株数为3株。2019年新造林郁闭(图1)。
自2019年4月至8月,记录试验林发笋时间、出笋数、退笋数等信息。同时于发笋初期,在试验林林中和林缘分别随机选择20株健康无虫害竹笋(5株作为标准样,15株作为备选竹)用挂牌和插牌进行标记,从竹笋露出土面2~3 cm开始,每3 d进行1次竹笋-幼竹的株高、枝下高和地径的测量,直至连续3次测量株高不再发生变化,视作株高生长结束。
2020年12月,沿着母竹去鞭方向,将整条鞭分成5等均分,分别为距离母竹0、2、4、6、8 m,并在每个位置范围内选取3株标准株作为重复,测量株高、枝下高、地径、节数、节长、节径及各构件生物量等指标。高度和长度用卷尺测量,粗度用游标卡尺测量,各构件生物量用电子天平测量。
1.3 数据分析
采用Microsoft Excel 2016和OriginPro 9.0对试验数据进行基本处理,采用SPSS 20.0软件进行单因素方差分析,LSD法进行多重比较。试验数据均为“平均值±标准差”,显著性水平设为P<0.05,极显著设为P<0.01。
2 结果与分析
2.1 长叶苦竹发笋及竹笋-幼竹高生长
长叶苦竹的发笋期为5月4日至6月27日,发笋期为52 d。发笋数量为6.93个/m2,成竹率为76.23%,退笋率为23.77%。林中和林缘的发笋起始时间不一致,林中发笋晚于林缘14 d,且林中的成竹高度高于林缘成竹高度(图2),以长叶苦竹的发笋时间为自变量,竹笋高度为因变量,利用Logistic模型进行拟合,林中和林缘竹株的竹笋-幼竹高生长规律均符合该模型,拟合系数均大于0.99,竹笋-幼竹生长速率均为“S”曲线,符合正态分布。如表1所示,林缘和林中长叶苦竹的生长进度不同,林中竹株抽枝展叶晚于林缘1个月。待竹笋-幼竹高生长结束后,林中植株的地径、株高、枝下高均显著高于林缘,分别是林缘的1.46、2.45、49.22倍。
2.2 距离母竹不同位置长叶苦竹的秆形规律
2.2.1 距离母竹不同位置的每节节长生长 由图3可见,距离母竹不同位置长叶苦竹的每节节长也都呈现规律变化。由鞭基部到顶部,距离母竹不同位置的长叶苦竹每节节长都呈现先增大后减小的趋势。距离母竹越远,竹秆每节平均节长和最长节长呈现递减趋势。母竹周围的最长节长为42.43 cm,距离母竹8 m的最长节长最短,为15.25 m,距离母竹2、4 m竹株的最长节长差异不显著。距离母竹6、8 m 竹秆的平均节长差异也不显著。距离母竹不同位置的最长节位数分布在第5~8节。
2.2.2 距离母竹不同位置的每节节径生长 如图4所示,距离母竹不同位置竹株的每节节径也呈规律性变化。长叶苦竹由秆基部到顶部,每节节径都是呈递减趋势。随着距离母竹越远,相对应的每节节径越小(图4-A)。不同位置的平均节径也是呈梯度变化,距离母竹越远,平均节径也越小。距离母竹最近的平均节径最大,为8.63 mm,距离母竹 8 m 的平均节径最小,为5.53 mm,它们之间具有显著性差异。母竹附近和距离母竹2 m竹株的平均节径差异不显著(图4-B)。

表1 不同生长位置长叶苦竹生长状况
2.2.3 距离母竹不同位置因子的相关性分析 如图5-A所示,长叶苦竹每节节长和每节节径存在一定相关性。为了确定长叶苦竹每节节长和每节节径的关系,利用对数、幂函数、线性、多项式等方程分别进行回归拟合。结果表明,线性拟合较好,距离母竹不同位置竹株每节节长和每节节径呈正相关关系。拟合系数最高为距离4 m时,系数达0.940 8。同样对距离母竹不同位置长叶苦竹的地径和株高,枝下高和株高的相关性进行回归拟合,线性拟合较好,地径和株高拟合相关性r2系数为0.815 7(图5-B)。枝下高和株高拟合系数较高,达0.900 3(图5-C)。
2.3 距离母竹不同位置生物量及分配比例
2.3.1 距离母竹不同位置的单株生物量 由图6所示,距离母竹越远,长叶苦竹的地上生物量、地下生物量以及总生物量均呈递减趋势。母竹周围的地上生物量最大,平均为511.93 g,距离母竹 8 m 的地上部分生物量最小,为197.80 g。距离母竹最近的地下部分生物量最大,为51.66 g,距离母竹位置8 m的地下生物量最小,为10.64 g。距离母竹0、2、4 m单株的地下生物量之间没有显著性差异,同样距离母竹6、8 m的单株之间也没显著性差异。母竹周围的总生物量最大,为563.59 g,距离母竹位置8 m的总生物量最小,为208.44 g。距离母竹不同位置的单株总生物量均无显著性差异。
2.3.2 距离母竹不同位置竹株生物量分配 由表2可知,距离母竹不同位置竹株的生物量分配均为秆的生物量最大,根的生物量最小。距离母竹8 m和6 m位置,均遵循秆>枝>叶>蔸>根的规律;距离母竹2.4 m位置,均遵循秆>叶>枝>蔸>根的规律。距离母竹8 m长叶苦竹的生物量分配为秆>枝>叶>蔸>根,秆的生物量最大,为75.57 g;根的生物量最小,为2.97 g;叶和其他构件差异显著,秆和枝以及根和蔸差异不显著。距离母竹6 m的生物量分配为秆>枝>叶>蔸>根,秆的生物量最大,为145.45 g,根的生物量最小,秆和其他构件差异显著。距离母竹4 m的生物量分配为秆>叶>枝>蔸>根,秆的生物量最大,为263.66 g,根的生物量最小,为17.3 g,秆和其他构件差异显著,其他构件之间生物量差异不显著。距离母竹2 m的生物量分配为秆>叶>枝>蔸>根,秆的生物量最大,为344.15 g,根的生物量最小,为19.66 g,秆和其他构件差异显著,其他构件之间生物量差异不显著。距离母竹 0 m 的生物量分配为秆>叶>蔸>枝>根,秆的生物量最大,为427.13 g,根的生物量最小,为 16.82 g,秆和其他构件差异性显著。
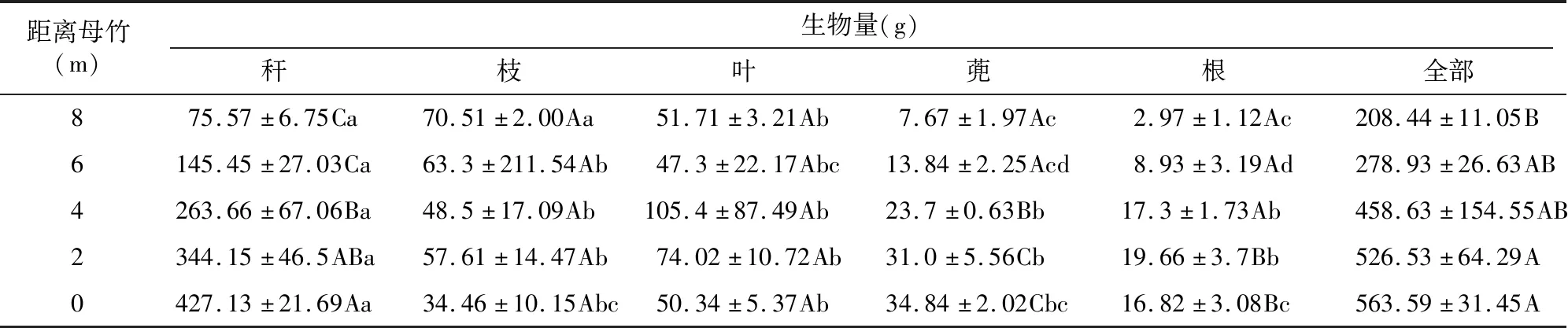
表2 距离母竹不同位置生物积累量及分配比例
2.3.3 距离母竹不同位置竹株生物量和各调查因子的相关分析 由表3可知,长叶苦竹距离母竹不同位置的生物量和其他测量因子的相关性表现为秆生物量与地径、株高、枝下高关系非常密切,相关性都达到了极显著水平。枝生物量与枝下高达到显著水平,与地径和株高没有显著相关性。叶生物量和其他因子没有显著相关性。地上生物量和地径、株高、枝下高的相关性非常密切,都达到了极显著水平。
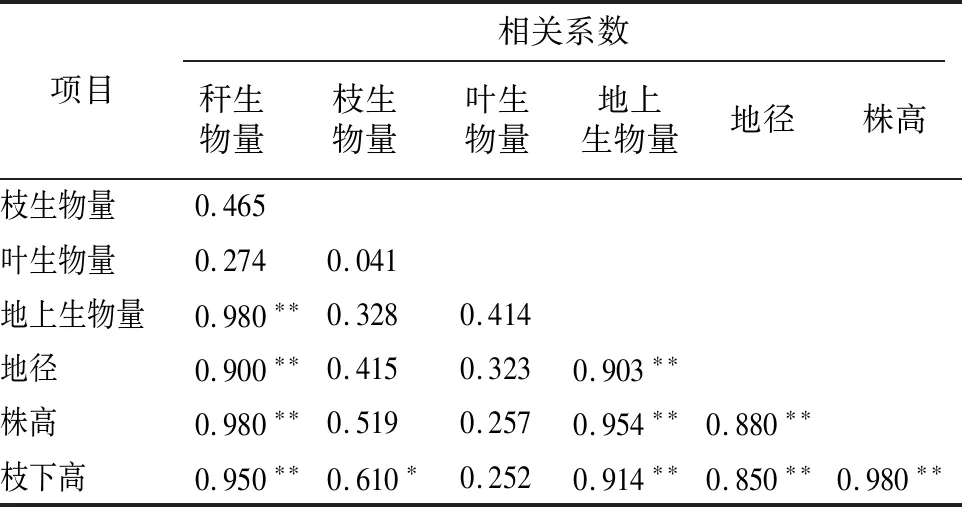
表3 距离母竹不同位置生物量及相关因子相关性分析
3 讨论与结论
Logistic模型广泛应用于竹类植物的笋-幼竹高生长规律的拟合[14-15]。本研究中五年生长叶苦竹林林缘和林中株高生长总时间一致,且高生长均呈现“慢—快—慢”的现象,符合逻辑斯蒂方程,与其他竹类植物如勃氏甜龙竹(Dendrocalamusbrandisii)、椽竹(Bambusatextilisvar.fasca)、梁山慈竹(DendrocalamusfarinosusMunro)、毛竹的高生长规律一致[16-19]。虽然长叶苦竹林中和林缘竹株完成高生长均历经70 d,但林中和林缘竹株的出笋时间不一致,林中竹株发笋时间晚于林缘14 d,且林中的成竹速度和成竹高度均显著高于林缘。植物生长会受到密度制约,不同密度种内竞争不同,影响植物对水分、养分、光照的吸收[20]。木本植物林分密度和其地径存在负相关关系,林分密度越小,其地径越大[21]。长叶苦竹林中密度大于林缘,其枝下高、地径和株高均显著大于林缘,说明混生竹的生长发育受其林分密度的影响较小,主要与竹类植物自身遗传决定的生物学特性有关。距离母竹越近,获得营养越多,越利于新生竹的生长发育。此外,植物抽枝展叶的时间也受生存环境的干扰,尤其受光照、温度、水分等因素的制约。长叶苦竹林缘处的密度小,光照度大,因此早于林中完成抽枝展叶过程。
研究竹类植物的秆形结构,可以了解竹类植物对生态环境的自适应性[22]。植物生长在不同立地条件下,株高、枝下高等生长情况相差较大[23]。随着植物种群密度的增大,植物形态指标如株高、冠幅、枝下高等也会发生可塑性调节来适应密度的改变[24]。本研究中距离母竹不同位置长叶苦竹的株高、枝下高等都随着离母竹位置越远,呈减少趋势。这可能因为竹株离母竹越远,竹林密度越小,离母竹越近,竹林密度越大,与植物在高密度下株高、枝下高等都会增大的研究现象[24-25]相符。相关研究表明,竹类植物的地径、株高和枝下高存在一定的相关性[26]。本研究结果表明,距离母竹不同位置长叶苦竹的地径和株高,枝下高和株高都呈正线性相关关系,即竹类植物的地径越大,株高越高;枝下高越高,株高也越高。此外,本研究中距离母竹不同位置长叶苦竹的每节节长和相对应的节径也存在正线性相关关系,且距离母竹越远,每节节长和节径呈递减趋势。上述结果表明,长叶苦竹在不同位置,其秆形形态建成具有一定的规律,距离母竹较近的长叶苦竹,高度和节茎要高于越远的竹株,与混生竹自身的生物学特性密切相关。
生物量是评价植物生长状况和生态系统功能的重要指标。植物资源最优化理论认为,植物会通过调节各构件生物量的配置来争取更多的光照、水分等资源。竹类植物各构件生物量的分配也是其对资源获取和适应外界环境的重要策略[27-29]。竹类植物各构件的生物量配置一般不同,受竹种和外界生长环境的影响[30]。竹类植物各构件生物量的分配在种群形态可塑性上起了基础性作用[31-32],不同构件生物量配比的关系同样也体现了竹类植物获取资源和适应外界环境的能力。长叶苦竹竹秆是无性繁殖植物,是由地下径和蔸发笋形成群体[33]。本研究中长叶苦竹表现为距离母竹越远,地上生物量、地下生物量及总生物量呈递减趋势,这依然与竹类植物特殊的生物学特性相关,距离母竹越近,越容易从母竹获得营养,其单株生物量优于距离母竹远的。此外,无论距离母竹多远位置,长叶苦竹单株生物量分配均为秆占比最大,根占比最小,这样的生物量分配有利于壮龄竹的形成,从而更有利于长叶苦竹这类混生竹由丛生状态转向散生状态,加速整个竹林的繁盛。
猜你喜欢
四川林业科技(2023年4期)2023-11-13
南方农业(2022年16期)2022-11-22
电脑迷(2021年12期)2021-11-28
广西植物(2021年9期)2021-10-16
现代农业科技(2019年23期)2019-01-14
南风(2018年22期)2018-01-30
福建农业科技(2015年1期)2015-04-17
