
虎毛色多样性的遗传演化
2022-11-22 08:12陆道炜罗述金
生物学通报 2022年1期
陆道炜 徐 霄 罗述金
(北京大学生命科学学院 北京 100871)
体表毛色和斑纹是哺乳动物最显著的形态学特征之一,发挥着隐蔽、警戒、交流及热调节等重要的功能,对物种的生存和繁衍具有重要意义。与其他随机产生的遗传突变一样,影响毛色表型的突变也会在自然界中不断产生,同时受到自然选择的作用,绝大多数属于同一物种的野生动物个体维持着统一的毛色和斑纹。并非所有的同种野生动物都毛色统一,一些哺乳动物(例如,美洲黑熊、棕熊、亚洲金猫、细腰猫等)具有一定的物种内毛色和斑纹的多样性。新出现的变异型如果不影响(甚至增强)物种的适合度,其基因频率则有可能在野生种群中得以保持,甚至逐渐演化形成稳定的种内多样性。
豹属(Panthera)包括虎、狮、豹、美洲豹、雪豹等5 种大型猫科动物,其中虎(Panthera tigris)是唯一具条纹而不是斑点的物种。虎仅分布于亚洲,现存6 个亚种:东北虎、华南虎、苏门答腊虎、孟加拉虎、印支虎、马来虎。对比分明的黄底色黑条纹的野生型是所有虎亚种标志性的毛色表型,但主要分布于印度次大陆的孟加拉虎,除野生型黄虎外,还存在4 种毛色多态型:白底黑纹的白虎(white)、金底棕纹的金虎(golden)、几乎全白的雪虎(snow)和因黑色条纹区域扩张和融合而表现出形成类似于黑化表型的拟黑化虎(pseudomela‐nistic,也称黑虎)(图1)[1]。这些非典型的色型是最早产生于野生种群的遗传变异型,不属于独立物种,也不是独立亚种。在某些色型被引入圈养之后,由于人们的偏好而大量近交繁育,导致了圈养个体中普遍存在着健康问题和生理缺陷,因此,其遗传起源和保育价值长期存在疑问和争议。本文将简要介绍目前已知的非典型虎色型的历史记录,以及近期关于虎毛色多样性的遗传机制的研究进展,并探讨这些遗传变异对物种生存和适应的潜在影响及其对虎的保护和圈养繁殖的科学意义。
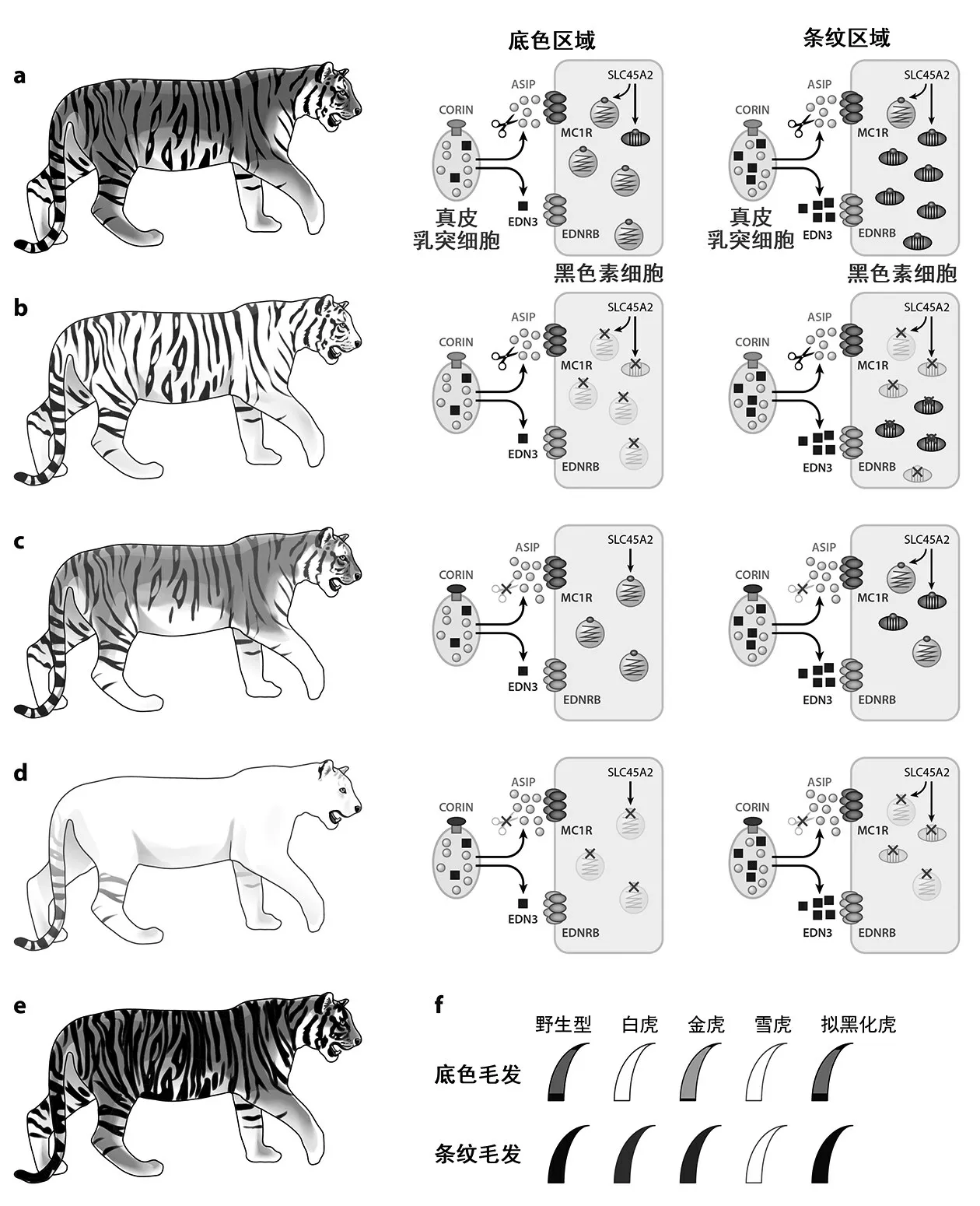
图1 虎的毛色多样性和遗传机制(改自Luo 等,2019)[1]
1 虎毛色多样性的历史记录
在虎的4 种毛色变异型中,白虎出现频率最高,也是可信记载中历史最悠久的。我国自古以来,白虎被奉若神明。在东亚,白虎被认为是吉祥的象征,神灵的化身。
野生白虎的历史最早可追溯至古印度莫卧尔王朝阿克巴(Akbar)大帝统治时期(公元1556—1605年),其编年史的一幅彩绘插画展现了皇帝和侍卫在1561年途经现印度中央邦时遭遇并猎杀4 虎的情形,其中2 只毛色明显偏浅清晰可辨,是最早的白虎记录。在19世纪至20世纪的前半叶,白虎的记录层出不穷。根据孟买博物学协会期刊(The Journal of Bombay Natural History Society)记载,在1907年至1933年间印度共有至少17 次猎杀白虎的记录,包括现阿萨姆邦(Assam)、奥里萨邦(Odisha)、中央邦(Madhya Pradesh)、比哈尔邦(Bihar)、北方邦(Uttar Pradesh)、贾坎德邦(Jharkhand)和恰蒂斯加尔邦(Chhattisgarh)等地。据估计在20世纪30年代,印度东北部的比哈尔邦分布约40 只白虎,而当时印度野生虎种群总数量约为40 000 只。表明白虎不是一个物种,甚至也不是独特的亚种,而是孟加拉虎中自然存在的一种稀少的变异型。然而,由于人类猎杀及栖息地的丧失和破碎化,野生虎种群在上世纪急剧下降,最后一次确切记录是1958年在比哈尔邦猎杀的一只雌性白虎,此后再没有过野外的报道。
目前,唯一有明确来源的圈养白虎是1951年捕获于雷瓦土邦的雄性白虎莫罕(Mohan)。莫罕与黄虎交配所得子一代均为黄虎,随后与其中一只雌性后代回交,于1958年首次得到人工繁育的白虎,并通过进一步的回交和近交建立了以白虎为主的家系。莫罕的后代被引进到印度、美国、英国并逐渐传播到世界各地,成为现存大多数圈养白虎的共同祖先。为了快速获得大量白虎,人们通常采取高度近亲繁殖的策略,导致很多白虎饱受病理缺陷的困扰,诸如流产、死胎、免疫系统缺陷、脊椎侧弯、髋关节发育不良等,白虎的存在与繁育因此长期遭到质疑和反对。一些反对者认为白虎是动物园或马戏团违背自然、通过人工繁殖制造出的白化病畸形,不值得保护。白虎之争,旷日持久。解决这一争议的关键在于圈养白虎常见的生理缺陷究竟是源于白虎毛色这一性状本身,亦或仅是近亲繁殖导致的恶果,解决这一问题就需要探讨其遗传机制。
与通体白色、虹膜浅红的白化病个体不同,白虎的体表遍布黑条纹,虹膜呈蓝色,表明白虎并非白化虎,其细胞内仍有合成真黑色素的功能,但决定黄底色的褐黑色素缺失从而呈现白底色。历史上有很多野生白虎成年个体被捕捉的记录,也说明白虎可在野外生存至成年并繁衍后代,其变异的毛色型没有严重影响其生存,推测圈养白虎的遗传缺陷更可能是近交的结果。一项对美国辛辛那提(Cincinnati)动物园的52 只白虎的观察也表明,如果消除近亲繁殖的影响,白虎和其他圈养动物在生理健康状况上并没有明显差异。广州长隆野生动物世界的繁育记录也表明,在引入了多个来源的白虎个体并扩大繁育种群数量后,近亲繁殖程度得到有效减缓,近年出生的白虎也大多没有斜视等缺陷。这说明只要没有近交,培育健康的白虎是可能的。
与白虎相比,金虎更为罕见(或许与其特征相对野生型差别不明显而容易被忽视有关)。在1929年前后,印度东北部阿萨姆地区曾记录到2 只“红虎”,体表整体呈棕黄色,条纹处不是黄虎的深褐色或白虎的黑色,而是比背景色深的红棕色,研究者根据当时拍摄的黑白照片中颜色的反差推断其为金虎。2014年后在印度东北部开展的野外调查中,再次发现了金虎。圈养种群中的金虎最早可追溯至20世纪80年代,目前,圈养金虎不超过30 只。
在圈养繁育种群中出现金虎的同时,还出现了全身几乎不带有任何条纹的雪虎。印度东北部的原住民将雪虎奉为神灵,然而,近代以来西方殖民者在该地区开发茶园和狩猎,迅速地将稀少的雪虎猎杀殆尽。20世纪80年代,美国辛辛那提动物园的白虎家系中出现雪虎。
虎的毛皮不仅与底色和条纹的色素合成变异相关,条纹的分布模式发生突变也导致其表型多样性,黑虎便是这样的代表。不同于豹和美洲豹中出现的黑化(melanism)现象,并未发现真正的虎黑化个体,只有由于黑色条纹扩张和融合而导致的黑虎,称为拟黑化(pseudo-melanism)。关于黑虎明确的记载仅出现在孟加拉虎中,但20世纪以前印度及周边地区关于黑虎的记载大多为黑豹的误称。近几年位于印度东部奥里萨邦的西密里波(Similipal)国家公园发现了较多野生黑虎。这些虎的体型比同年龄的野生型黄虎稍小,加之该种群与印度其他地区的虎种群隔离程度严重,研究者怀疑该表型的出现是隔离小种群内部近交和遗传漂变的结果。目前,野生黑虎基本集中在该公园,数量呈增长的趋势。在印度的3个圈养种群也有黑虎的记录,但其数量和比例与白虎和金虎相比都更低。
2 虎毛色多样性的遗传机制
哺乳动物的皮肤和毛发的颜色由毛囊中的黑色素细胞(melanocyte)分泌的黑色素(melanin)决定。黑色素有2 种,其中,真黑色素(eumelanin)形成深棕色或黑色,褐黑色素(pheomelanin)形成黄色或红色。这2 种色素的含量、比例和分布决定了哺乳动物毛色和肤色的多样性[2]。野生型黄虎的毛发包括2 种毛色模式:橘黄色毛发为两端真黑色素、中间插入褐黑色素条带的agouti 毛发,而条纹处则为仅包含真黑色素的纯黑毛发[3]。白虎和金虎分别为底色和条纹2 种毛色模式合成通路异常所形成的表型;而雪虎则是真黑色素和褐黑色素的合成同时受到了抑制。拟黑化虎的变异不涉及色素合成通路的改变,而是表皮色素分布的分区出现异常从而导致条纹区域扩张。
基于经典遗传学中的家系分析,研究人员基本确定这几种变异类型都是单基因隐性突变导致。白虎和金虎由2 个独立的隐性突变位点分别决定,而雪虎是同时携带了这2 种突变的纯合子。黑虎则是由另一个独立的与斑纹形成相关的遗传位点所致。近10年来,随着基因组学领域技术的发展,科研人员已破译了目前已知所有的虎毛色多样性的遗传机制,以下将对导致白虎、金虎、雪虎、黑虎等4 种变异表型的基因和突变进行介绍。
2.1 白虎:Slc45a2基因突变对于褐黑色素合成的影响 白虎体表的褐黑色素合成被阻断,表现为白底色和棕黑条纹[4]。白虎体表缺失褐黑色素,其真黑色素的合成通路仍然存在,体现在蓝色虹膜及深色条纹。本研究团队基于限制性酶切位点关联DNA测序(whole genome sequencing,RAD-seq)和全基因组测序(restriction-site-associated sequenc‐ing,WGS),对一个包含了16 只圈养个体的白虎家系开展全基因组关联分析(genome wide associa‐tion study,GWAS),并在130 只没有亲缘关系的虎样本集中进行了验证[5]。表明一个发生在转运蛋白SLC45A2 上的氨基酸改变(A447V,第447 位由丙氨酸变为缬氨酸)形成了白虎表型。Slc45a2是一个已知的参与色素合成的基因,编码一个12 次跨膜的离子转运蛋白,并且特异性地表达在黑色素细胞中负责合成黑色素的黑色素小体(mela‐nosome)。该基因上的一些突变在人类、低地大猩猩、多伯曼短毛猎犬中造成白化病(oculocutaneous albinism),另一些变异则与小鼠、马、鸡、鱼中色素减少的性状相关[6]。Slc45a2基因在欧洲人群历史上曾经历过强烈的自然选择,与其他人群相比形成显著的种群差异,也是与欧洲人浅肤色相关的重要遗传因素之一[7-8]。
作为质子转运蛋白,SLC45A2 在维持黑色素小体的内环境pH 稳态上起着重要的调节作用[9]。蛋白质结构预测表明,A447V 氨基酸变化可能影响SLC45A2 的转运通道,造成黑色素小体内pH 环境失衡[5]。由此推断,白虎的真黑色素和褐黑色素生成都将受到显著影响,而这与白虎体表仍有明显黑色条纹的性状不符。本研究团队通过探究猫科动物斑纹模式形成的遗传机制发现,体表深色的条纹或斑点区域(例如,家猫的条纹、猎豹的斑点)受另一独立细胞通路调控,通过Edn3的高表达促进特定区域亦即条纹或斑点区的真黑色素合成[10]。这一色素分布模式的维持机制是贯穿于整个猫科动物演化的保守通路,据此推测,白虎可能借助于这一不受Slc45a2突变影响的黑色素合成机制维持了条纹处的真黑素色合成,进而表现出白色底色上棕黑色条纹[11]。
Slc45a2的多态性(polymorphism)与人的肤色及其他一些动物的毛色有关。但除了极少的突变与人类白化病相关,更多的突变表现为人、马、鹌鹑、鸡等物种在该基因自然存在的多态性,这些多态性使毛色或肤色变浅,对其他生理途径或器官功能的影响甚微。因此,从白虎形成的遗传机理分析,白虎不是白化病,也不是先天的遗传缺陷或畸形。从遗传多样性分析,白虎具有保护和研究的价值。这一结论结束了长期以来动物学界关于白虎是否是一种遗传缺陷的争论,为虎的遗传多样性保护及白虎的繁殖策略等提供了科学的佐证[1]。
野生白虎的记录一直零星但持续地出现,说明白虎隐性基因在整个孟加拉虎的野生种群中,以一个较小的比例稳定地传递。尽管携带这一变异的基因频率较低,但这种多态性已在野生虎种群中延续了至少数百年,应当被视为自然的虎遗传多样性的一部分。白虎这一变异没有对生存和繁衍产生显著的负面影响。可能是因为虎及其有蹄类(鹿和野猪等)猎物在视觉上与人类的三原色色觉存在差别,或许在捕食者和被捕食者动物眼中白虎与黄虎并无太大差别的缘故。
由于已发现导致白虎毛色的基因,通过基因鉴定的方法可甄别2 只携带隐性“白虎基因”的孟加拉黄虎。依据经典孟德尔遗传定律,它们交配后可产生的后代中有1/4 的几率是白虎。
2.2 金虎和雪虎:Corin基因突变对于黑色素合成的影响 金虎是由于真黑色素合成通路受到抑制,形成淡金黄色的底色上覆红棕色(而非黑色)条纹。雪虎的2 种黑色素的含量都极低,除了尾部和肢端几乎通体白色。金虎体表背景处的agouti 毛发中褐黑色素条带占比更大,该变异位点被称为“宽带(wideband)”位点。雪虎的表型则是来自于白虎的white位点和金虎的wideband位点复合隐性纯合所致。white位点即为发生在Slc45a2基因上的突变,而wideband位点并不位于任何一个已报道的毛色决定基因(MC1R、ASIP、TYR、TYRP1)上。本研究团队选择圈养虎家系在wideband位点有性状分离的19 只个体(包括1 只金虎,8 只雪虎、10 只白虎)进行限制性酶切位点关联DNA 测序、全基因组测序及全基因组关联分析,并在38 只金虎与雪虎(wideband位点突变型纯合)及159 只野生型黄虎和白虎(至少携带一个wideband位点野生型基因型)的样本中进行了验证分析,将wideband遗传位点定位到Corin基因第587 位氨基酸[3]。野生型的组氨酸在金虎中突变为酪氨酸(H587Y),导致了金虎的浅黄底色;该氨基酸变异与发生在Slc45a2基因的A447V 突变均为纯合时,则表现为条纹和色素沉积几乎完全消失的雪虎表型。
Corin基因编码了一个跨膜的丝氨酸蛋白酶,通过抑制ASIP 通路参与色素形成[12]。哺乳动物agouti 毛发中段的褐黑色素条带源自毛发形成过程ASIP 信号蛋白(agouti signaling protein)的短暂高表达,导致黑色素的合成由真黑色素向褐黑色素的转变[2]。CORIN 蛋白在ASIP 蛋白下游发挥作用,通过降解ASIP 参与调控毛发生长周期中ASIP蛋白的生效时间长度。金虎的H587Y 突变削弱了CORIN 蛋白的功能,干扰了ASIP 蛋白的及时降解,导致其作用时间变长,从而在agouti 毛发中生成更长的褐黑色素条带[3],整体表型则表现出真黑色素显著减少而呈浅金黄色毛发。
在双突变纯合的雪虎中,白虎所携带的突变和金虎所携带的突变效果叠加后造成了黑色素合成过程的整体抑制[3]。发生在Corin基因上的突变本身对于ASIP 活性的调控作用并不足以解释雪虎的体表条纹颜色改变。白虎在条纹处还有Edn3基因的高表达,对应于白虎中特定于条纹区域的黑色素合成,即通过EDN3 蛋白激活的黑色素合成通路可不受ASIP/MC1R 的调控[5]。金虎和雪虎的表型则说明,EDN3 蛋白的这一效果可能在雪虎中被Corin和Slc45a2的双重突变抑制。
2.3 黑虎:Taqpep基因突变对于毛色斑纹形成的影响 虎的拟黑化表型是由于真黑色素的沉积增加而导致了更密的条纹,表现为体表黑色条纹的加粗和部分融合,尤其是在背部和尾部,使得其体表黑色面积增加乃至在部分角度下看起来为全黑色[13]。家猫(Felis catus)中的斑点猫(blotched cat)类型和猎豹(Acinonyx jubatus)中的王猎豹(king cheetah)变异都表现为类似体色,此前的研究将这种条纹与背景毛发的排列模式异常定位于Taqpep基因。Taqpep基因在胚胎发育早期确定深色和浅色区域的预模式(pre-pattern)过程中起着重要作用,其突变会影响黑色素分布的区域,进而导致体表黑色条纹的扩张和融合[10]。
为探究黑虎的毛色型是否也由Taqpep基因突变导致,Sagara 等[14]对8 只圈养虎的家系(包括3 只黑虎)进行全基因组测序,发现所有黑虎都携带有TAQPEP 蛋白序列上第454 位的由组氨酸变为酪氨酸的纯合突变(H454Y)。该位点位于TAQPEP 蛋白与金属离子结合区域上,在其他物种中都是保守的组氨酸。研究人员进一步通过对13 只圈养虎的非损伤性样品(毛发、粪便、唾液等)的研究,验证了该突变仅在黑虎中为纯合,而在西密里波国家公园以外的野生虎种群均未发现Taqpep基因突变型。基于粪便等非损伤性采样,研究者从西密里波国家公园野生虎种群识别出12 只不同个体,其中4 只为突变基因型纯合,6只为杂合,反映了黑虎基因型在该种群的高频率分布。针对印度圈养种群中黑虎个体的溯祖分析也显示该遗传突变最可能源自西密里波种群。对于黑虎突变型的地域性分布而且在特定种群中的高基因频率,研究者主要认为是在近百年来受到隔离的小种群中发生近交及随机漂变积累的结果。此前针对于黑化美洲豹的研究显示,更深的毛色可能有利于捕食者在热带丛林的隐藏[15],因而,拟黑化的毛色也可能具有一定的适应性。
3 总结及展望
由于栖息地丧失、猎物减少及偷猎、盗猎,野生虎种群数量已从一个世纪前的大约10 万只锐减至目前的4 000 只左右。与此同时,世界圈养虎数量庞大,保守估计至少上万只。除了野生型黄虎和新近报道的黑虎之外,其他白虎、金虎和雪虎等毛色型的野外记录已极为罕见,虽然人工繁育条件下有相当数量圈养种群,但其遗传起源和保育价值长期存在争议。值得注意的是,这些不同毛色的虎都是物种内的多态性,甚至基本可确定是孟加拉虎亚种内起源的遗传变异,不是独立的物种或亚种,因此,单一的表型不应该成为独立的保护和管理单元。
通过遗传机制和野外记录的分析得知,白虎不是白化病,其Slc45a2基因变异本身对于个体的生存和繁衍没有显著影响,而且在野生虎种群中延续了至少数百年,应当被视为自然的虎遗传多样性的一部分,具有保护的意义和价值。现存圈养白虎所遭受的健康问题是由于严重近交所致,并非因为白虎性状是遗传缺陷。数百年亦或数千年,人们无法准确得知白虎在地球上最早出现的时间。然而,人们知道,在南亚次大陆的广袤原野中,在人类将其捕杀殆尽之前,白虎及其黄虎同胞曾经作为一个种群而一起自由驰骋。人们固然不必盲目地过量繁殖白虎,但也不应妖魔化地排斥白虎。动物园作为生物多样性的“诺亚方舟”,笔者建议应该担负起维系虎的遗传多样性的责任,在圈养种群中排除近交,保持少量但是健康的白虎,将其纳入到规范的虎繁育管理体系中。
金虎和雪虎虽然也有野外的记录,其频率远小于白虎,而且由于Corin基因除了黑色素细胞通路以外还参与了血压调节等重要的生理过程,其突变除了导致毛色变异外,是否还产生其他缺陷性状进而引起个体适合度的下降,需要更深入的研究。近几年在印度东北发现的野生金虎位于高度破碎的栖息地,有可能与小种群中近交加剧及遗传漂变有关,值得密切关注。现存野生黑虎仅分布于印度东部的一个地区,也可能与栖息地片段化及种群隔离有关。
除了对于物种保护的意义,揭示虎的毛色形成遗传机理也为研究哺乳动物黑色素合成通路和适应性演化进行了积极的探索。对于虎毛色变异的研究拓展了人们对哺乳动物毛色遗传机制的认识,例如,金虎决定基因Corin基因是哺乳动物中报道的第2 个wideband遗传位点,由此表明了哺乳动物agouti 毛发类型突变的多重起源,也体现了通过非模式生物研究动物表型多样性的价值和意义。
猜你喜欢
养猪(2021年4期)2021-08-26
今日农业(2021年11期)2021-08-13
今日农业(2020年14期)2020-12-14
小学阅读指南·低年级版(2017年10期)2017-11-20
散文选刊·下半月(2017年9期)2017-09-21
小说月刊(2015年11期)2015-04-23
火花(2015年6期)2015-02-27
小学生作文·小学中高年级适用(2014年3期)2014-08-18
通俗小说报(2009年3期)2009-03-11
通俗小说报(2009年1期)2009-01-12
