
基于SSR标记的麻栎天然群体遗传多样性分析
2022-11-29 13:51解孝满鲁仪增宗绍宁李文清
南京林业大学学报(自然科学版) 2022年3期
吕 锋,解孝满,,韩 彪,鲁仪增,王 磊,董 昕,王 艳,陆 璐,刘 莉,宗绍宁,李文清,*
(1.山东师范大学生命科学学院,山东 济南 250014;2.山东省林草种质资源中心,山东 济南 250102)
麻栎(Quercusacutissima)属于壳斗科(Fagaceae)栎属(Quercus),最新的栎属分类系统将其分为8组,分别为中间栎组(Sect.Protobalanus)、本都栎组(Sect.Ponticae)、活栎组(Sect.Virentes)、白栎组(Sect.Quercus)、红栎组(Sect.Lobatae)、青冈栎组(Sect.Cyclobalanopsis)、冬青栎组(Sect.Ilex)和麻栎组(Sect.Cerris),共 423 种[1]。其中,麻栎属麻栎组,同组树种还有栓皮栎(Q.variabilis)和小叶栎(Q.chenii)。麻栎在我国分布于东经91°~123°、北纬18°~41°的地区,是东亚暖温带和亚热带地区森林植被中重要的组成树种之一[2]。麻栎为喜光树种,耐干旱瘠薄,具有良好的水土保持功能,其木材硬度大、有弹性、耐腐蚀,为优等家具、乐器等用材。其树皮、壳斗及果实可提取栲胶,也可药用,具有收敛止血、涩肠固脱等功效,树叶具有抗突变活性[3-4]。
近年来,对麻栎种质的相关研究主要集中在组织培养及扦插繁殖技术[5]、苗期生长特性[6]、优良家系的选择[7]和群体遗传多样性[8-9]等方面。对栎属遗传多样性研究也较多,如孟旭[10]利用10对nSSR引物对麻栎33个居群510个个体进行分析,结果显示麻栎的遗传多样性水平并不高[等位基因数(Na)=5.264,有效等位基因数(Ne)=3.504,Shannon多样性指数(I)=1.218,观测杂合度(Ho)=0.283,期望杂合度(He)=0.595,多态性百分率(PPPL)=96.06%]。徐小林等[11]基于16对SSR分子标记分析结果,认为栓皮栎5个天然群体的遗传多样性较丰富。秦英英等[12]利用SSR标记研究发现山西省辽东栎(Q.liaotungensis)群体的遗传多样性较高。王雁红等[13]利用SSR分子标记对24个短柄枹栎群体(Q.serratavar.brevipetiolata)研究发现其遗传多样性不高。李文英等[14]使用等位酶和AFLP 技术研究认为,我国8个蒙古栎 (Q.mongolica)天然群体的遗传多样性水平偏低。其中,由于对辽东栎、短柄枹栎等的研究取样存在局限性,群体间也存在较远的地理隔离,并不能完全反映该物种水平下的遗传多样性和遗传结构。本研究基于前人对栎属植物研究的SSR引物,通过筛选出适用于麻栎的SSR引物,并对麻栎天然群体的遗传多样性进行研究,以期为麻栎种质资源的保护和利用提供理论基础。
1 材料与方法
1.1 试验材料
2018—2020年在河南、辽宁、山西、云南、河北、山东、江苏等7省的麻栎自然分布区,共采集了8个麻栎群体150份种质资源。收集地点的经度和纬度用GPS定位软件记录,各居群的采样样本数见表1。为保证样品具有代表性且避免采集到同一家系的个体,群体内随机选取成年大树,且每个样本个体至少间隔50 m。采集新鲜无病斑的麻栎叶片,使用硅胶干燥保存。
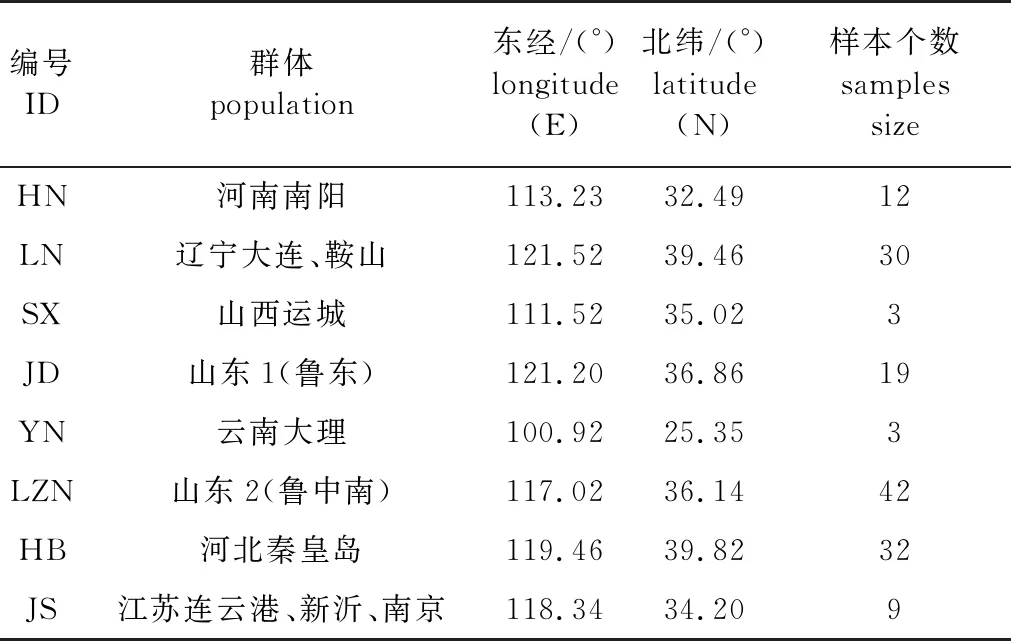
表1 采样点基本信息
1.2 DNA提取及PCR扩增
1.2.1 基因组DNA提取
利用DNA提取试剂盒(9768)(宝生物,大连)提取所有麻栎样本,用ND-2000紫外分光光度计(上海天能科技有限公司,上海)对其DNA浓度进行检测,并采用1%(质量分数)的琼脂糖凝胶电泳检测DNA质量,放置-20 ℃保存,用于后续试验。
1.2.2 PCR反应体系及电泳检测
PCR反应在Veriti 96孔快速梯度PCR仪(ABI,美国)上完成,25 μL PCR反应体系如下:10 ×TaqBuffer(with MgCl2)2.5 μL,10 μmol/L dNTP(mix)1 μL,20 ng模板DNA,10 μmol/L正、反向引物各1 μL,5 U/μL的Taq0.5 μL,ddH2O 18 μL。PCR反应程序为:95 ℃预变性3 min,94 ℃变性30 s,退火30 s,72 ℃延伸30 s,循环次数为35次;最后72 ℃延伸6 min,将扩增产物于4 ℃条件下保存。
PCR扩增产物用8%(质量分数)聚丙烯酰胺凝胶电泳检测,经银染法染色并显色后,在凝胶成像仪(上海复日科技有限公司)上拍照保存。
1.2.3 SSR引物筛选
由于微卫星两端的序列是比较保守的,所以同属物种之间的差异很小,SSR引物具有通用性[15],SSR引物来自前人已开发的栎属SSR引物[16],由上海生工生物有限公司合成,从56对引物中筛选出了18对稳定性高、多态性好的引物(表2)用于后续试验。将筛选出的引物和DNA样品送至上海生工生物有限公司进行微卫星分型试验。
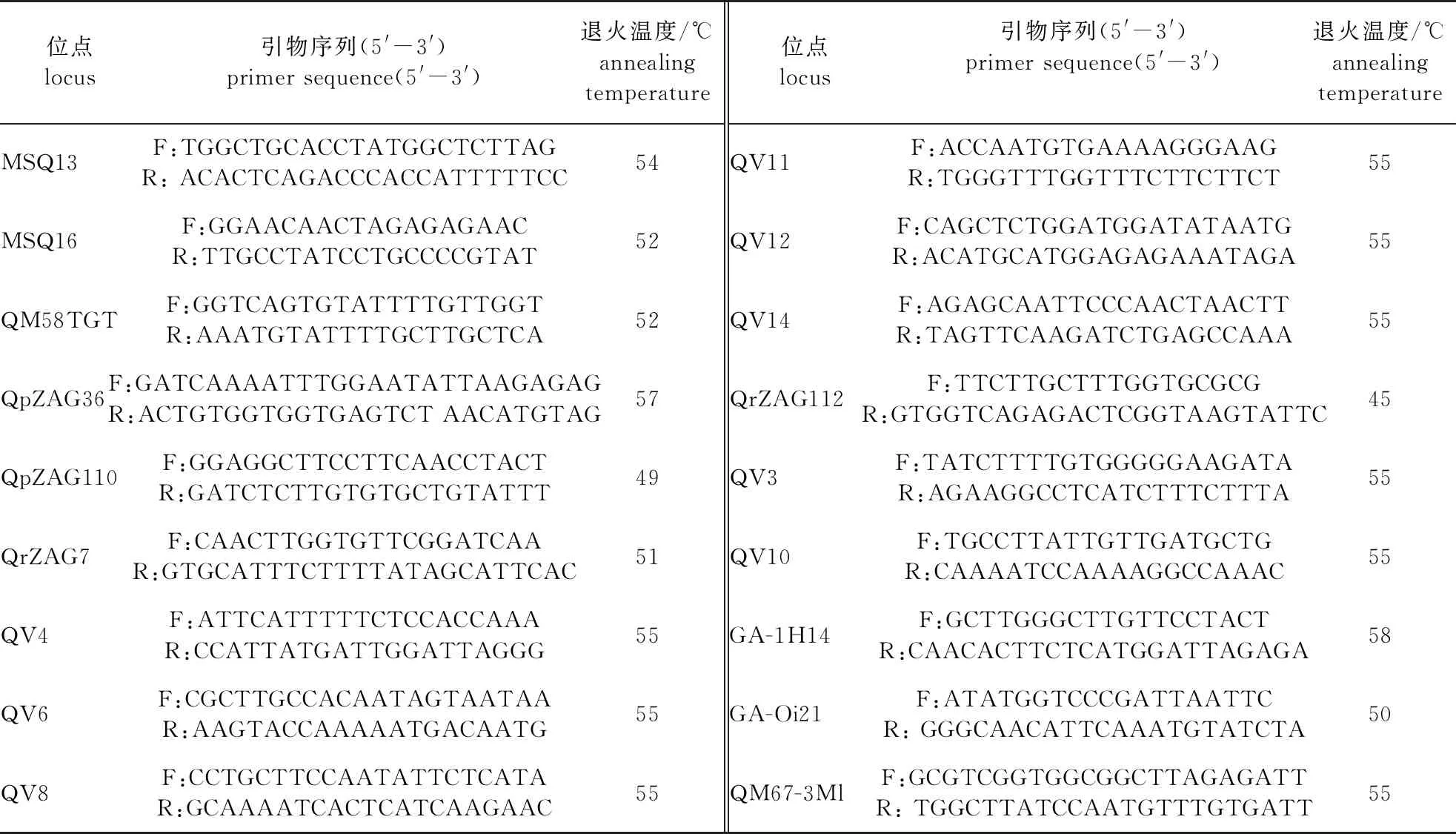
表2 18对SSR引物序列信息
1.3 数据处理
使用Genemarker 2.2.0软件对毛细管电泳数据分析,采用GenAIEx 6.51软件[17]将数据整理,并计算各群体的遗传多样性参数期望杂合度(He)、观测杂合度(Ho)、等位基因数(Na)、有效等位基因数(Ne)、Shannon多样性指数(I)等,并进行主成分分析(PCoA)和AMOVA分析。同时,计算遗传分化系数(Fst),并根据公式Nm=(1-Fst)/4Fst[18]计算基因流(Nm)。基于Nei’s遗传距离,使用MEGA 7.0.26[19]软件基于非加权组平均法(UPGMA)进行群体聚类。
利用Structure 2.3.4软件[20]对麻栎群体的遗传结构进行分析,设置K值为2~7,每个K值的运行次数为10次,每次不作数迭代(length of burn-in period)和MCMC(Markov chain monte carlo)均设置为100 000次,采用ΔK法进行聚类数值设定,使用Structure Harvester计算ΔK的最大值。
2 结果与分析
2.1 SSR位点的多样性
18个SSR位点的遗传多样性、F检验及基因流估算结果见表3。由表3可知,平均等位基因为5.625个,等位基因数(Na)的变化范围为2.000~13.125个,其中以QM58TGT位点的等位基因数最多,而QM67-3M1的等位基因数最少。各个位点的有效等位基因数(Ne)的变化范围为1.778~9.547个,平均值为4.104个,最多的位点为QM58TGT,其次为MSQ16(6.944),最少的位点为QM67-3M1。Shannon多样性指数(I)的变化范围为0.632~2.267,平均为1.338。各个位点的观测杂合度(Ho)为0.625~1.000,平均为0.895。期望杂合度(He)的变化范围为0.444~0.861,平均为0.645。表明筛选出的18对麻栎SSR引物具有丰富的多态性。
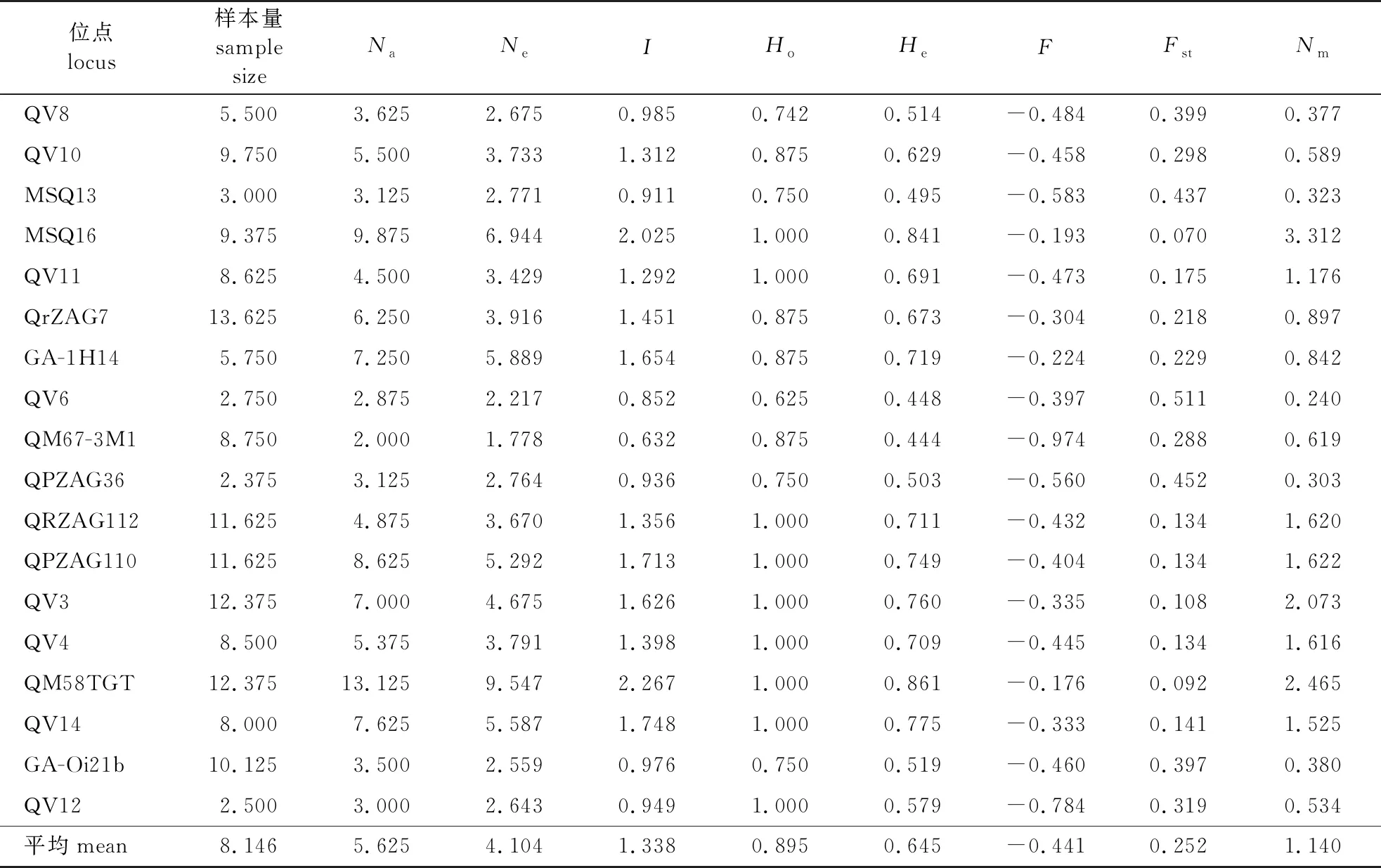
表3 18个SSR位点的遗传多样性、F检验及基因流估算
2.2 不同麻栎群体的遗传多样性
麻栎群体遗传多样性分析见表4。麻栎群体等位基因数(Na)为2.278~10.389个,平均为5.625个,山东鲁中南群体(LZN)的最高,云南群体(YN)的最低。有效等位基因数(Ne)为2.102~6.405个,平均为4.104个,山东鲁中南群体(LZN)的最高,云南群体(YN)的最低。Shannon多样性指数(I)为0.748~1.929,平均为1.338,山东鲁中南群体(LZN)的最高,云南群体(YN)的最低。观测杂合度(Ho)为0.722~1.000,平均为0.895。期望杂合度(He)为0.441~0.805,平均为0.645。多态性百分率(PPPL)平均为89.58%。以上说明麻栎群体的遗传多样性丰富,遗传多样性水平较高。各群体的固定系数(F)均小于0,均值是-0.441,说明麻栎群体内的基因型分布不平衡,各群体中存在杂合子剩余,种群内存在较多的杂交和远交现象。
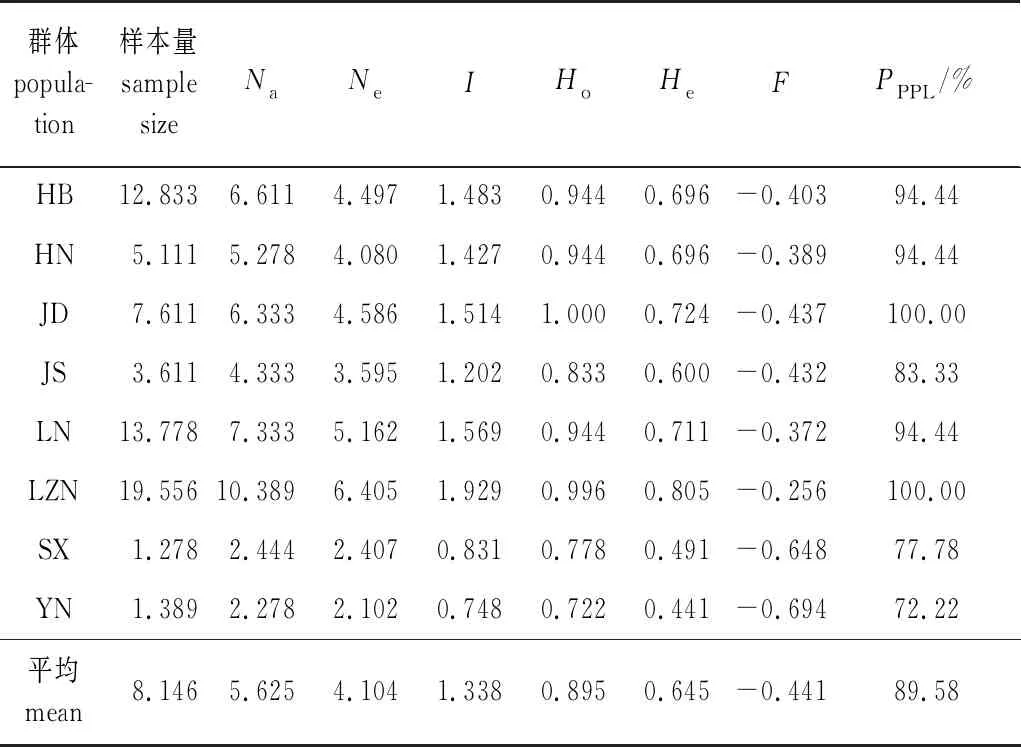
表4 麻栎群体遗传多样性
2.3 不同麻栎群体的遗传分化
麻栎群体的遗传分化系数(Fst)平均为0.252,表明麻栎群体间的遗传分化中等。基因流(Nm)为0.240~3.312,平均为1.140,亦表明麻栎群体间可能发生遗传分化的程度较小(表3)。AMOVA(表5)分析结果显示,麻栎群体的变异主要发生在群体内(98%),而只有2%的变异发生在群体间。
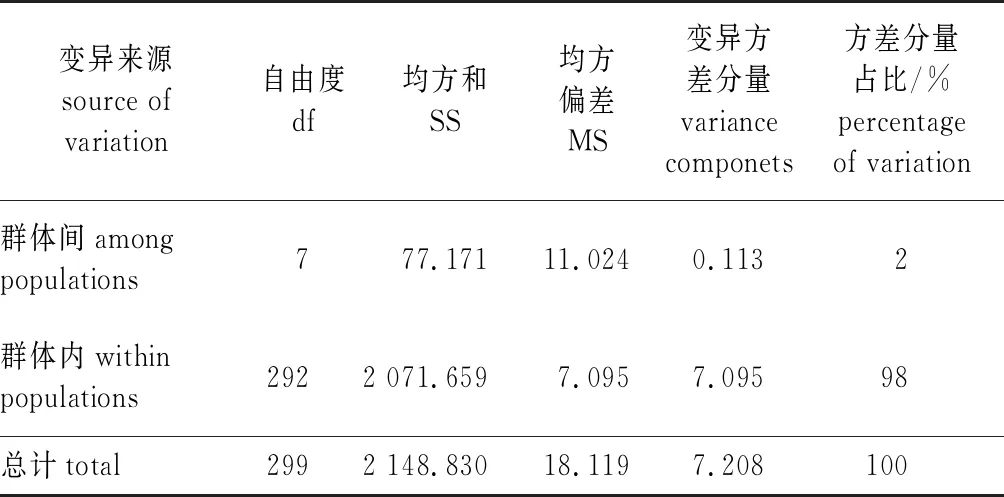
表5 麻栎群体分子方差分析表
2.4 麻栎天然群体间遗传关系分析
8个麻栎天然群体的遗传相似度与遗传距离分析显示(表6),麻栎群体间的遗传距离为0.222~1.587,遗传相似度为0.205~0.801。其中,辽宁群体(LN)和鲁中南群体(LZN)群体间的遗传距离最小,遗传一致度最大;云南群体(YN)和江苏群体(JS)群体的遗传距离最大,其遗传一致度最小。
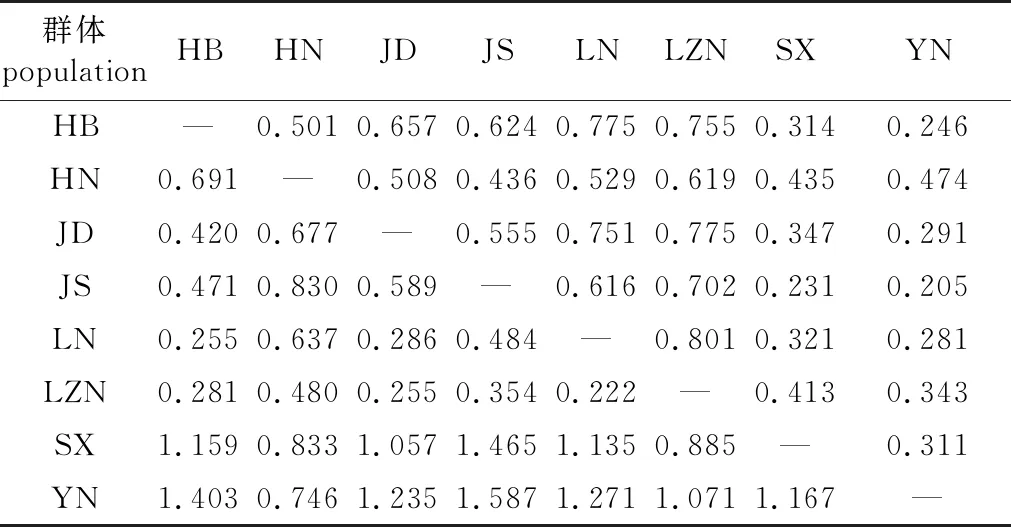
表6 麻栎8个天然群体遗传相似度与遗传距离
根据欧氏距离类平均法(UPGMA)对8个麻栎天然群体进行聚类分析,结果如图1所示。当欧式距离为12时,可以将8个群体划分为2组,其中,Cluster1包括辽宁群体(LN)、河北群体(HB)、山东鲁东(JD)群体、山东鲁中南(LZN)群体、江苏群体(JS)、山西群体(SX),各群体间的距离依次增大,亲缘关系逐渐变远。Cluster1包括2个亚组,其中Cluster1A,包含1个群体,为山西群体(SX);Cluster1B包括其他5个群体。Cluster 2只包括2个群体,分别为河南群体(HN)、云南群体(YN),其与上述Cluster1的群体间的距离最大,亲缘关系较远。上述麻栎群体总体呈现出沿“东北—西南”方向的地理变异规律。
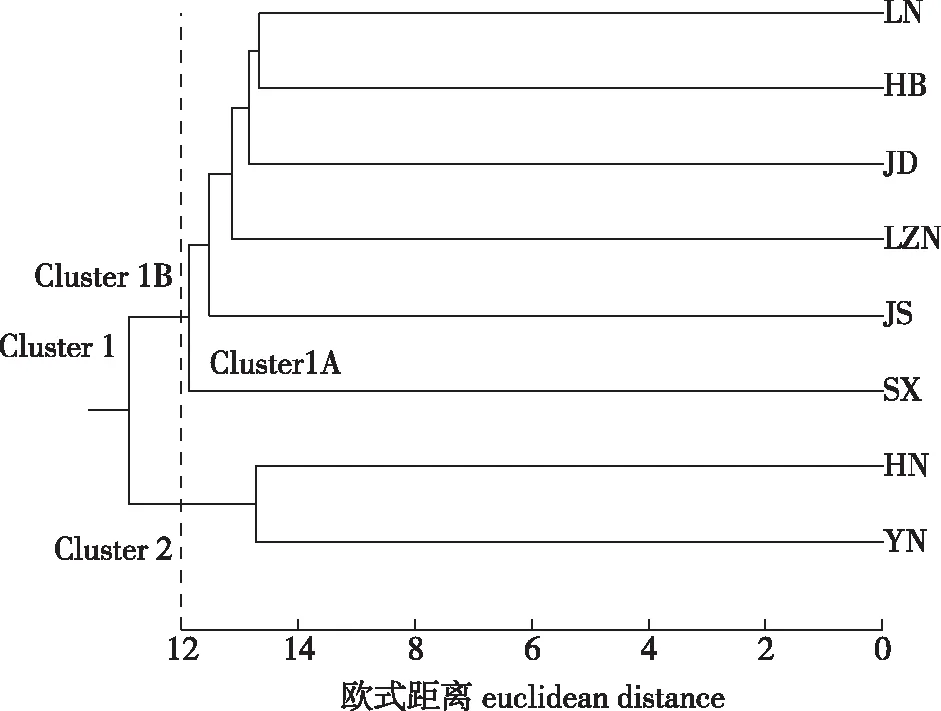
图1 8个麻栎居群UPGMA聚类树
对8个麻栎群体的150个个体的主成分分析显示(图2),西南方向群体[河南群体(HN)与云南群体(YN)]的不同个体基本聚在一起,山东鲁中南群体(LZN)的少量个体迁入;河南群体(HN)的少数个体迁出,并与其他6个群体的个体聚在一起。东北方向麻栎群体的个体之间聚类后仍较为分散,规律性不强,其可能存在交叉引种或者基因渐渗等现象。
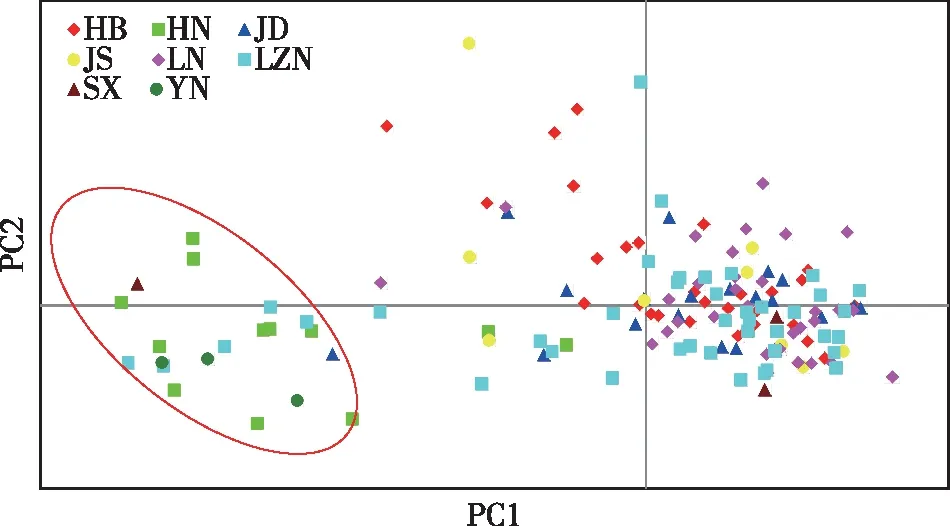
图2 麻栎群体150个个体主成分分析结果
2.5 不同麻栎群体间遗传结构分析
为了进一步研究麻栎群体间的遗传关系,基于贝叶斯方法对8个麻栎群体进行Structure分析(图3)结果显示,当K=2时,ΔK值最大(图3A),表明150份麻栎材料可能来自2个不同的理想类群。按照相似度对不同个体进行归类,可分为红色标注群体和绿色标注群体(图3B)。其中,云南群体(YN)的全部个体,河南群体(HN)的大部分个体,江苏群体(JS)、山西群体(SX)、山东鲁东(JD)和鲁中南群体(LZN)的少量个体,它们之间的相似度高,归为一类,为红色标注群体;辽宁群体(LN)全部个体,江苏群体(JS)、山西群体(SX)、山东鲁东(JD)和鲁中南群体(LZN)的多数个体,河南群体(HN)的少量群体,它们之间的相似度高,归为另一类,为绿色标注群体。部分个体的遗传信息显示来自上述两个群体。进一步表明,麻栎群体沿“西南—东北”方向加剧分化,并且在部分群体之间存在基因渐渗现象。在聚类分析中也观察到类似的结果,笔者还发现了Structure分析和聚类分析之间的一些差异,这可能是因为聚类分析为每条线分配了一个固定的分支位置,而结构分析导致了将个体分配给组的亚群成员百分比[21]。
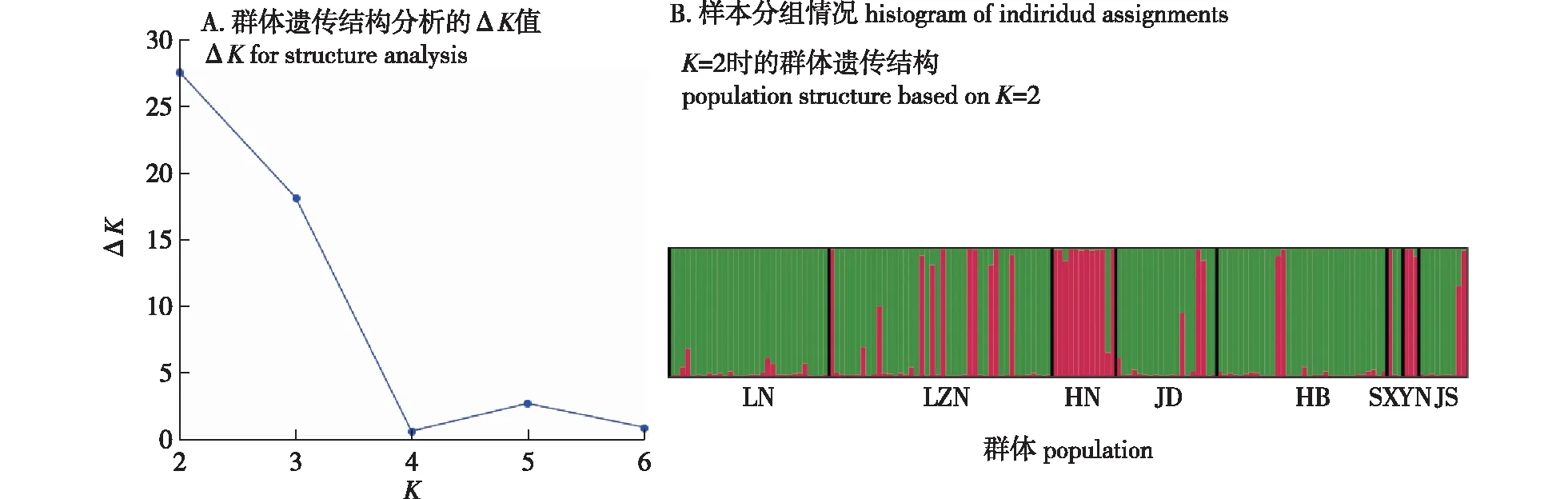
图3 基于贝叶斯分析的麻栎居群Structure聚类结果
3 讨 论
3.1 麻栎群体的遗传多样性
遗传多样性是物种长期生存和进化最基础的条件[22],遗传多样性越高,物种适应环境的能力就越强[23]。本研究中麻栎群体的等位基因数(Na)平均为5.625个,有效等位基因数(Ne)平均为4.104个,Shannon多样性指数(I)平均为1.338,观测杂合度(Ho)平均为0.895,期望杂合度(He)平均为0.645,表明麻栎群体的遗传多样性丰富,遗传多样性水平较高。这与叶青雷等[8]、彭礼琼等[9]分别基于SRAP和 ISSR标记得到的结论基本一致。但低于部分栎属其他植物的遗传多样性。其原因可能为,麻栎属于风媒传粉的植物,距离增加会降低花粉的浓度和传播效率,使产生的种子数量下降,随着距离增加,孤立的个体更容易发生近交产生种子[24];人为活动如砍伐、移植等的影响会使麻栎群体范围缩小,降低了其遗传多样性[25-26];另外,SSR标记的无效等位基因广泛存在,也是影响其遗传多样性的一个重要原因[27]。
3.2 麻栎群体遗传分化
群体的遗传结构改变是由于随机的遗传漂变及基因流的隔离[28]。Kremer 等[29]研究表明栎属种内群体间的遗传分化很小,分化系数在0. 01~0. 17,平均值为 0. 06。本研究中麻栎的群体间遗传的遗传分化程度亦不高(Fst=0.252,0.007~0.051),仅有2%的遗传变异发生在群体间,而在群体内的遗传变异达到了98%。Hamrick等[30]研究显示基因流Nm>4时,群体间的基因交换相对充分,从而抵抗遗传漂变,防止群体间因遗传漂变而发生遗传分化。本研究的基因流平均为1.140(0.240~3.312),进一步说明麻栎群体间存在遗传漂变,且其遗传分化程度较低。这与栎属植物的高遗传性和低遗传分化的普遍现象是基本吻合的[2]。
3.3 麻栎群体的遗传结构
遗传变异在空间分布上产生遗传分化,造就了一个物种种群的遗传结构[31]。群体遗传结构很大程度上受人为活动的影响,如对次生林分的改造行为[32],本研究采样的林分多属于次生林,林分生长过程中的不同环节,如交配、繁殖、地理分布范围改变及遗传交流等都会改变其群体的遗传结构[33]。Structure分析和UPGMA聚类分析中,均将麻栎群体分为2组,PCoA对麻栎个体之间的分析结果与其基本一致。并且群体之间呈现出沿“东北—西南”方向的变异规律。但是群体间存在交叉引种或基因渐渗现象。
3.4 麻栎的保护建议
野外调查发现,麻栎自然群体多为零星分布,人类的活动使麻栎所处生境严重片段化。因此,应切实地保护麻栎的生存环境,并且通过有效措施来阻止人为活动造成的破坏,逐渐恢复群体的生机。另外,可通过运用常规方法与分子生物学技术来研究和评价麻栎遗传多样性,制定相应的原地保存与迁地保存策略,开展栎类树种的杂交育种工作,加快培育速生、抗逆性强的优良品种,以此来丰富麻栎资源的遗传多样性。本研究基于SSR分子标记的麻栎遗传多样性表明,麻栎在物种水平上的遗传多样性水平较高,但是,西南方向群体相对于东北群体的遗传多样性低。在开展保护麻栎种质资源时,首先对其野生群体及其生境进行原地保护,遗传多样性高的群体应重点保护,当然对于边缘地区的保护也不能放弃;还可以在全分布区建立长期观测种源的试验基地,基于当前及未来研究目标建立麻栎种质资源迁地保存库,合理利用麻栎资源[34]。
猜你喜欢
中国糖料(2022年3期)2022-07-04
中国农学通报(2022年12期)2022-06-01
中国糖料(2022年2期)2022-04-06
智慧健康(2021年17期)2021-07-30
中学生物学(2019年7期)2019-10-17
新课程·下旬(2018年9期)2018-11-14
考试周刊(2017年26期)2017-12-12
校园英语·下旬(2017年7期)2017-07-14
科技视界(2016年27期)2017-03-14
中学生理科应试(2016年7期)2016-05-14
